池塘高密度培育刀鲚当年鱼种的脂肪酸组成变化
2022-12-05施永海谢永得刘永士徐嘉波袁新程邓平平
施永海,谢永得,刘永士,徐嘉波,袁新程,邓平平,税 春,蒋 飞
(上海市水产研究所,上海市水产技术推广站,上海 200433)
【研究意义】刀鲚(Coilianasus),又名长颌鲚,俗称刀鱼、毛刀鱼,属于鲱形目(Clupeiformes)鳀科(Engraulidae)鲚属,是溯河洄游性鱼类,与长江鲥鱼(Tenualosareevesii)、河豚(Takifugu)统称“长江三鲜”[1]。近年来,由于自然生境的变化,长江的“大刀”刀鲚(150 g/尾以上的刀鲚)的比率明显减少,其年渔获量也呈现波动下降趋势[1]。中国政府2019年开始禁捕长江刀鲚,2020年开始“长江十年禁捕”,禁止在长江捕捞野生长江刀鲚,市民要吃刀鲚,只能选择海捕的刀鲚或者养殖的刀鲚。早在2013年,上海市水产研究所就获得了人工繁育的批量刀鲚苗种[1],但是国内刀鲚人工养殖技术还没有完全破解,存在养殖产量低、规格小的现象,主要关键技术参数比较模糊,还需要长期地、分阶段地集中突破各技术难关。在刀鲚规模化人工繁育技术完全成熟的情况下,接下来需要攻关的难题是刀鲚当年鱼种规模化生产,改变长江刀鲚当年鱼种粗放型池塘养殖模式,解决刀鲚当年鱼种培育成活率低、产数量少且不稳定的问题,提高长江刀鲚0+龄鱼种池塘单位产量,以实现刀鲚当年鱼种池塘高效培育,生产出大规格刀鲚鱼种:①可提供给广大水产养殖户进行成鱼养殖,给养殖户带来巨大的经济利益,同时让美味重回百姓餐桌;②可提供给增殖放流,对长江刀鲚渔业资源保护做出较大贡献。【前人研究进展】鱼类体内的脂类不仅是其生长发育所需能量的主要储存物质,而且还参与调节生理活动和组织器官的构建[2-4],同时,分析脂肪水平和脂肪酸组成及含量也可以评价鱼类的营养状况[5]。近年来,国内外有关刀鲚的营养状况和营养需求的应用基础研究主要集中于发育早期脂肪酸组成变化[1]、亚成鱼[6]及成鱼的肌肉营养成分分析及评价[7-11]。【本研究切入点】刀鲚0+龄幼鱼阶段是刀鲚自然洄游过程中入海前营养积累的重要阶段,也是刀鲚实际养殖过程中冬季越冬前的营养积累阶段,而高密度培育的刀鲚当年鱼种的营养积累和能量储存过程及状况还未涉及。笔者之前研究了常规密度(4167 ind/667 m2)和高密度(7216 ind/667 m2)培育的刀鲚当年鱼种的生长、消化及非特异性免疫能力[12]。【拟解决的关键问题】本文研究高密度培育刀鲚当年鱼种各生长阶段(当年7—11月)鱼体脂肪酸组成及含量的变化规律,评价刀鲚当年鱼种(0+龄)池塘高密度培育过程中的脂类营养状况,以期为刀鲚当年鱼种池塘高效培育提供基础数据。
1 材料与方法
1.1 试验鱼
实验用刀鲚当年鱼种(0+龄)是上海市水产研究所奉贤五四基地自行全人工繁育获得。养殖试验方法:池塘(56#)面积2335 m2,水深1.5 m。鱼种放养前,池塘中培养饵料生物(枝角类和桡足类等,脂肪酸组成及含量见表1),当生物量达到20~40 ind/L,即可放养;于2018年6月26日放养刀鲚鱼种25 255 ind,放养密度为7216 ind/667 m2,放养规格:体长(2.93±0.48)cm,平均体重(0.11±0.05)g,n=30。试验期间,不投喂配合饲料,刀鲚的饵料主要依靠池塘中活饵料,每周检测水体中的饵料生物,当生物量低于1 ind/L,及时从其他池塘中捞取补充;池塘中设置1台1.5 kW增氧机,每天晚上开机9~10 h,遇特殊天气增加开机时间;试验用水为当地河水(盐度1.8~2.3),每2周换水1次,换水量为1/3。实验期间,自然水温17.8~36.8 ℃(图1),pH 7.85~8.52,DO≥7.57 mg/L,TAN≤0.22 mg/L,NO2-N≤0.19 mg/L。当年鱼种养殖试验于2018年11月1日结束,养殖成活率为79.7%。

表1 淡水枝角类桡足类的脂肪酸组成及含量
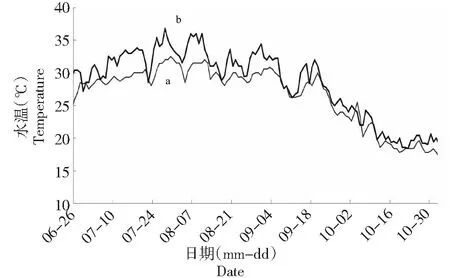
图1 池塘水温变化情况(a为上午、b为下午)Fig.1 The change of water temperature in pond (a is morning,b is afternoon)
1.2 样品采集与预处理
将鱼种的放养日期(2018年6月26日)设定为养殖0日龄,于7月26日开始,每月拉网随机捞取30尾,平均分为3个重复,取样日期和规格详如表2所示。取样后,立即在冰盘上解剖去除内脏团,将剩下的整条鱼体放入-20 ℃的冰箱先预冷冻,再置于实验室-76 ℃冰箱中保存[1]。

表2 取样幼鱼的基本情况
1.3 水分、总脂和脂肪酸测定方法
鱼体(去内脏的胴体)样品中的水分用冷冻干燥法测定,冷冻干燥后的鱼体重量即为鱼体干重,鱼体干重的总脂含量采用氯仿甲醇法测定;各脂肪酸含量先按照GB/T22223—2008测定,再用峰面积归一法计算其相对含量[1]。
1.4 数据处理和统计
用单因素方差分析对刀鲚生长各阶段的数据差异进行统计分析,用Duncan氏法作多重比较,以P<0.05为差异显著[1]。
2 结果与分析
2.1 水分和总脂含量变化
池塘高密度培育刀鲚鱼种的鱼体水分含量随着生长直线下降,从7月的82.27%显著下降到11月的76.17%(表3);除了7月外,鱼体干重的总脂含量随生长呈现显著的上升趋势(P<0.05),9月鱼体干重总脂含量显著高于8月(P<0.05),显著低于10月和11月(P<0.05),而总脂含量在10月和11月之间无显著性差异(P>0.05,表3)。

表3 池塘高密度培育的刀鲚当年鱼种水分和总脂含量(n=3)
2.2 脂肪酸的组成变化
2.2.1 各脂肪酸相对含量的变化 刀鲚当年各时间段鱼种的C16:0百分含量在8月和9月均排第一,在7月、10月和11月均排第二,除7月(23.31%)外,C16:0的百分含量随生长发育显著下降 (P<0.05),从8月的29.13%跌至11月的21.21%(表4)。C18:1n9c百分含量在7月、10月和11月均排第一,在8—9月排第二,随着刀鲚生长,C18:1n9c的百分含量呈现显著的连续隔月波动趋势(P<0.05),波动范围为23.56%~28.90%。各时间段的C16:1百分含量均排第三,除7月(13.75%)外,C16:1的百分含量随生长发育显著下降 (P<0.05),从8月的14.29%跌至11月的8.71%。C18:0百分含量较丰富,呈现先升后降 (P<0.05)的趋势,最高值出现在8—9月份(7.62%~7.76%)。
刀鲚当年鱼种的C22:6n3(DHA)、C20:5n3(EPA)和C22:5n3(DPA)的百分含量随生长发育呈现相似的先降后升趋势(P<0.05),最低值均出现在9月,其值分别为0.25%、0.65%和0.18%,最高值均出现在11月,其值分别为5.78%、4.47%和1.66%,导致EPA+DHA出现相似的趋势(P<0.05),最低值(0.90%)在9月(P<0.05),最高值(10.26%)在11月(P<0.05)。
2.2.2 ∑SFA、∑MUFA和∑PUPA百分含量的变化 刀鲚当年鱼种的脂肪酸组成中∑SFA比例随生长呈现显著的先升后降趋势(P<0.05),最高值(49.08%)出现在9月(P<0.05),最低值(34.49%)出现在11月(P<0.05) 。∑MUFA比例随生长呈现显著的连续波动趋势(P<0.05),波动范围为35.29%~44.27%,11月的∑MUFA比例显著低于(P<0.05)7—10月的(图2)。但刀鲚当年鱼种的∑PUPA比例随生长呈先降后升趋势 (P<0.05)。最低值(8.29%)在9月,最高值(30.22%)在11月(P<0.05),其中∑n3PUPA和∑n6PUPA的含量也呈现相似的先降后升趋势(P<0.05),最低值均在9—10月(3.56%~4.04%和0.99%~1.00%),最高值均在11月(18.28%和3.42%,P<0.05) 。
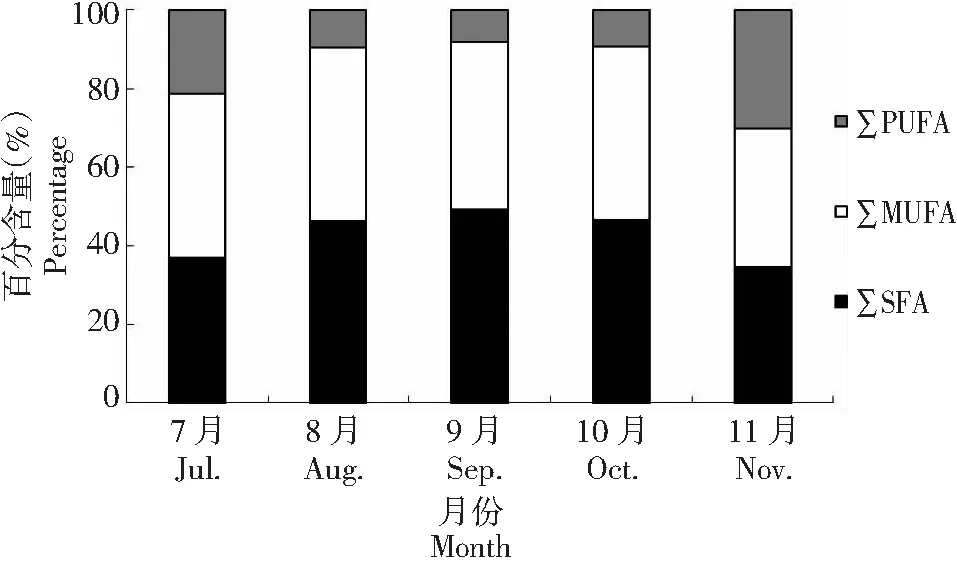
图2 池塘高密度培育刀鲚当年鱼种SFA、MUFA和PUPA的变化Fig.2 Changes of SFA, MUFA, and PUPA of 0+ age young fish C. nasus high-density cultured in pond (n=3)
2.2.3 ∑SFA/∑UFA和∑n3PUPA/∑n6PUPA比率的变化 随刀鲚当年鱼种生长,其∑SFA/∑UFA呈现显著的先升后降趋势(P<0.05),最高值(0.96)出现在9月(P<0.05),最低值(0.53)出现在11月(P<0.05)。但当年鱼种的∑n3PUPA/∑n6PUPA比率随生长呈现先微弱下降、再显著上升的趋势(P<0.05),最低值(3.58)出现在9月,最高值(5.35)出现在11月(P<0.05)。
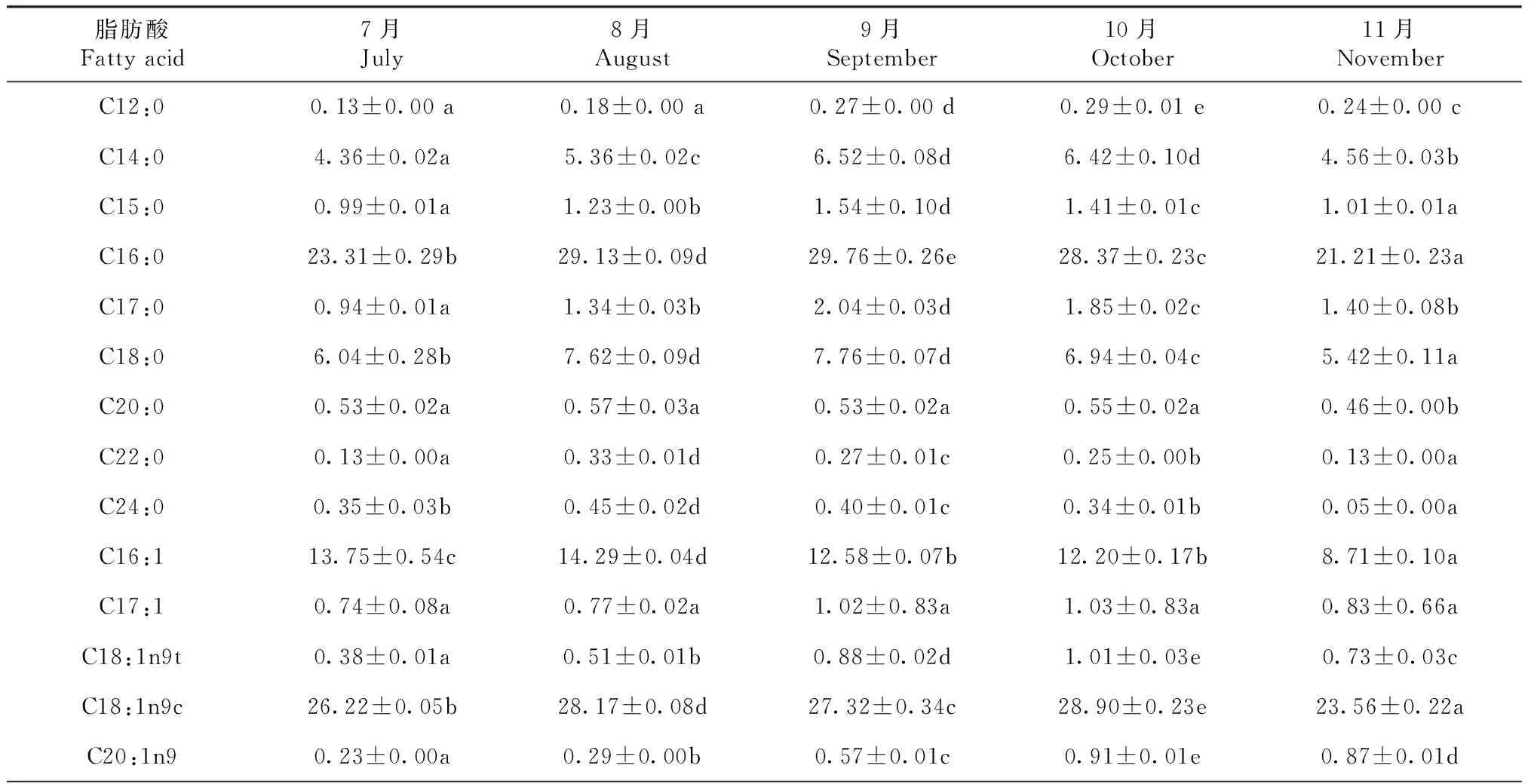
表4 池塘高密度培育的刀鲚当年鱼种脂肪酸组成及含量

续表4 Continued table 4 (n=3, %)
3 讨 论
3.1 池塘高密度培育刀鲚当年鱼种总脂含量的变化
在鱼类早期发育过程中,脂肪是主要的能量储存物质,还参与其组织构建和生理调节。本研究显示,池塘高密度培育刀鲚当年鱼种在培育早期(7月26日至8月7日)鱼体干重的总脂含量无显著变化,甚至从7.89%略微下降到7.56%,造成这现象的可能原因有二:一是因为培育早期刀鲚鱼苗还没有完全被鳞,鱼体内的器官特别是神经系统可能没有发育完全,鱼体除了支持生长外,还需要一定量的额外能量来支持其发育完全,造成刀鲚当年鱼种培育早期,鱼体脂肪积累较少;二是因为此时正值高温季节,高水温可能造成刀鲚幼鱼的胁迫,刀鲚鱼种摄食可能受到一定影响。
随着培育时间的推移,进入刀鲚当年鱼种培育到中期(8月7日至10月5日),幼鱼发育完全,鱼体脂肪迅速富集,鱼体脂肪含量从8月7日的7.56%显著增加到10月5日的10.27%,造成这种现象的可能原因有二:第一,鱼种培育至中期,池塘水温进入刀鲚鱼种的适宜温度范围(依据苗种培育适宜水温为23~24 ℃[13]和笔者10多年的养殖刀鲚经验,刀鲚养殖的适宜范围在20~28 ℃),其新陈代谢较盛夏季节平稳,同化作用上升,异化作用下降;第二,随着生长发育,刀鲚鱼种个体捕食能力增强,摄食量大增,鱼体内营养物质积累加速。培育到中期,刀鲚鱼体内脂类富集的现象在养殖银鲳(Pampusargenteus)[5]和美洲鲥(Alosasapidissima)[14]均有相似体现。
但是,刀鲚当年鱼种培育到后期(10月5日至11月1日),鱼体内脂类的积累速度减缓,其含量在10月和11月之间无显著差异,可能原因是池塘平均水温降至20 ℃以下,池内饵料生物的繁殖受到一定影响,特别是淡水枝角类。随着刀鲚幼鱼的生长,个体长大,同时刀鲚培育密度高,池塘内刀鲚总量多,总的食物需求量明显增加,导致池塘内的饵料不能完全满足高密度培育池塘刀鲚幼鱼的摄食,造成鱼体内营养物质的储存减缓,虽然有外源饵料生物补充,但是外源补充是基于“每周检测池塘中的饵料生物低于1 ind/L”限定的,这就可能导致池塘内饵料生物量长期处于刀鲚幼鱼需求量的低限,也最终导致刀鲚脂类积累受到不利的影响,这也印证了袁新程等[12]的高密度培育刀鲚生长受到影响的研究结果:常规密度(4167 ind/667 m2)刀鲚当年鱼种的体重相对生长率(710.50%)远远高于高密度(7216 ind/667 m2)培育的(108.48%)[12]。
3.2 池塘高密度培育刀鲚当年鱼种的脂肪酸变化规律
鱼体内的脂肪酸组成与其摄食饵料密切相关[14-16]。本研究中,池塘高密度培育刀鲚当年鱼种摄食相对寡含C16:0 (14.92%)和C16:1(9.54%)的淡水饵料生物后,随着个体生长发育,鱼体的C16:0和C16:1的百分含量分别从8月的29.13%和14.29%下降到11月的21.21%和8.71%,体内脂肪酸的含量与其食物中的脂肪酸含量大多呈正相关,这与施永海等[14]和刘志峰等[15]的研究结果一致。但是这种密切相关也不是绝对的,如:本研究刀鲚当年鱼种C18:1n9c的百分含量呈现显著的连续隔月波动趋势,波动范围为23.56%~28.90%,但其食物中的C18:1n9c的百分含量只有9.64%;再如:刀鲚当年鱼种的C20:5n3(EPA)的百分含量随生长先降后升,最低值出现在9月(0.65%),最高值出现在11月(4.47%),其食物中的C20:5n3(EPA)含量丰富(14.71%),该现象在美洲鲥上也有体现[14],因此,鱼体内脂肪酸含量除与其食物相关外,还可能与其生长发育阶段及环境条件有关[17]。
研究发现鱼体内的DHA主要是通过食物链富集起来的[18],刀鲚当年鱼种的C22:6n3(DHA)随生长呈现先降后升趋势,从7月的2.07%降到9月的0.25%,然后再上升到11月的5.78%。培育早期的刀鲚鱼种还属于稚鱼阶段,发育还没有完全,特别是神经系统,如脑组织,需要消耗大量的DHA[1],而刀鲚鱼种食物(淡水饵料生物)中的DHA也不丰富(1.19%),同时刀鲚仔幼鱼又不具备将18C PUFA转化为20C和22C HUFA的功能[1],导致鱼体DHA迅速下降,到培育中期,刀鲚幼鱼发育完全,原来消耗DHA用以发育的现象已不存在,虽然刀鲚幼鱼继续摄食DHA含量不丰富的淡水饵料生物,但是幼鱼体内可能对DHA进行了蓄留,减少了DHA的消耗,导致池塘高密度培育后期刀鲚鱼体的DHA显著上升(5.78%)。
在池塘高密度培育刀鲚当年鱼种后期,DHA和EPA等含量迅速上升,导致∑n3PUPA从9月的3.56%迅速升至11月的18.28%,这符合刀鲚洄游习性:在自然环境下,刀鲚在江河中繁育[19],当年幼鱼于秋冬季下海越冬,随着气温下降和刀鲚洄游入海,体外环境盐度逐渐提升和水温逐渐下降,刀鲚幼鱼机体为了维系细胞膜的渗透性和流动性,就需要更多的∑n3PUPA[1-2],同时早期研究也表明,随着刀鲚仔稚幼鱼生长,对盐度的适应性也增强[20];然而,在本研究中,刀鲚当年鱼种池塘培育环境的盐度一直保持在1.8~2.3,刀鲚幼鱼生活环境的盐度相对稳定,刀鲚幼鱼依然能启动∑n3PUPA的积累来应对生活环境可能产生的盐度上升和水温下降,可能与其内在调节机制有关,刀鲚幼鱼机体为了适应洄游入海越冬的需要。
另外,鱼类的脂肪酸组成与饵料关系密切,本研究只是提供了初始的饵料生物组成和脂肪酸组成,但在刀鲚当年鱼种培育池塘中饵料生物组成有一定的变化,这在一定程度上会影响本研究高密度培育刀鲚当年鱼种的脂肪酸变化。
3.3 池塘高密度培育刀鲚当年鱼种脂肪酸组成的特点
高密度培育造成鱼类的应激会诱导其脂肪酸代谢发生变化[21],尤其是长时间的高密度胁迫后,鱼类的脂肪酸组成会产生变化,这可能与机体采用消耗脂肪酸来应对应激胁迫和血清皮质醇的释放有关[22-23]。也有研究发现,高密度培育鱼类脂肪酸的改变与该鱼肝脏和肌肉中的去饱和酶上升有关,如,虹鳟鱼(Oncorhynchusmykiss)[24]。比较刀鲚各阶段主要脂肪酸组成及百分含量:脂肪酸含量排前三的均是C18:1n9c、C16:0和C16:1[1, 6-8];本研究中,高密度培育的刀鲚幼鱼的C16:0含量(21.21%~29.76%)与亚成鱼及成鱼肌肉(18.54%~27.41%)[6-8]相似,明显高于仔鱼阶段(7.45%~9.15%)[1];而当年鱼种鱼体的C16:1含量(8.71%~14.29%)与仔鱼阶段的(13.66%~13.98%)[1]相似,略高于亚成鱼及成鱼肌肉(6.96%~9.53%)[6-8];当年鱼种鱼体的C18:2n6c含量(2.52%~3.56%)处于仔鱼阶段(4.12%~4.34%)[1]和亚成鱼及成鱼之间(0.64%~3.16%)[6-8]。
在鱼体内,脂类主要储存在白肌组织中[25],脂类的合成和分解在肝脏中[25],高密度胁迫会造成鱼类肝脏MUFA中的占比较高的脂肪酸C18:1n9的降低,如,金头鲷 (Sparusaurata)[21]。鱼类中,MUFA (特别是 C18:1n9) 用于能量供给[26],高密度胁迫下鱼类需要消耗更多的能量去应对环境应激的胁迫,因此,MUFA的下降与过于拥挤的鱼类的能量需求直接相关。本研究中,池塘高密度培育后期(10月5日至11月1日),由于个体生长,刀鲚幼鱼拥挤胁迫更加明显,刀鲚当年鱼种鱼体的∑MUFA和 C18:1n9c含量均分别从10月的44.42%和28.90%迅速下降到35.29%和23.56%,说明刀鲚幼鱼拥挤胁迫导致了MUFA (特别是 C18:1n9)的消耗;与此同时,池塘高密度培育的刀鲚当年鱼种鱼体的C18:1n9c含量(23.56%~28.90%)均低于仔鱼阶段(50.39%~57.00%)[1]、亚成鱼阶段(45.40%~49.97%)[6]和成鱼阶段(41.73%~44.84%)[7-8],同时,当年鱼种的∑MUFA含量(35.29%~44.42%)均低于仔鱼阶段(65.14%~72.26%)[1]、亚成鱼及成鱼阶段(54.06%~61.43%),这再次证明过于拥挤导致刀鲚幼鱼需要额外消耗鱼体内的MUFA (特别是C18:1n9)来应对拥挤的应激。另外,这可能也与本研究刀鲚摄食的饵料淡水枝角类桡足类的C18:1n9c含量(9.64%)和∑MUFA含量(21.13%)较低有一定相关。
另外,本研究中池塘高密度培育的刀鲚当年鱼种鱼体的C14:0、C18:0和C18:3n3含量(4.36%~6.52%、5.42%~7.76%和2.24%~5.91%)均分别高于仔鱼阶段(2.38%~2.92%、1.93%~2.50%和0.85%~0.97%)[1]、亚成鱼阶段(1.71%~1.28%、2.12%~2.38%和0.34%~0.77%)[6]和成鱼阶段(1.82%~3.58%、2.04%~4.03%和0.46%~0.88%)[7-8],同时,当年鱼种的∑SFA含量(34.49%~49.08%)均高于仔鱼阶段(12.20%~15.27%)[1]、亚成鱼及成鱼阶段(28.41%~31.24%)[6-8],这也可能与本研究中刀鲚摄食的饵料淡水枝角类桡足类的C14:0(5.66%)、C18:0(2.76%)、C18:3n3(12.13%)和∑SFA(26.13%)含量较丰富有关,也可能与仔鱼和成鱼阶段的饵料和饲料的脂肪酸组成有关。
通常,海水鱼的∑n3PUPA/∑n6PUPA较淡水鱼高[1-2],刀鲚是溯河繁育洄游性鱼类[20],鱼苗在淡水中生长发育,到当年秋冬季,幼鱼随江河水流下海过冬。刀鲚胚胎和仔鱼的∑n3PUPA/∑n6PUPA数值较低(1.78~2.38),其特征更接近淡水鱼[1, 27-28],而刀鲚亚成鱼或成鱼肌肉的∑n3PUPA/∑n6PUPA数值较大(7.71~8.85)[6-8],其脂肪酸特征偏向于海水鱼类[2-5, 18],本研究的池塘高密度培育刀鲚当年鱼种鱼体的∑n3PUPA/∑n6PUPA数值(3.58~5.35)在他们之间,这符合刀鲚幼鱼从淡水到海水的洄游规律,脂肪酸特性也符合从淡水特征逐步转变成符合海水鱼类特征,本研究刀鲚幼鱼正处于转变适应期。
4 结 论
高密度培育的刀鲚当年鱼种鱼体的水分含量随生长发育直线下降;培育早期(7月26日至8月7日),鱼体干重的总脂含量略有下降,培育中期(8月7日至10月5日)鱼体脂肪迅速富集,到培育后期(10月5日至11月1日),随个体生长,饵料竞争和拥挤胁迫更加明显,刀鲚幼鱼鱼体内脂肪富集速度减缓,同时,幼鱼为应对拥挤胁迫需要消耗更多MUFA (特别是C18:1n9),导致刀鲚鱼种鱼体的∑MUFA(尤其是C18:1n9c)含量迅速下降。池塘高密度培育的刀鲚当年鱼种可能已经受到环境拥挤胁迫。
