细菌微室及其在合成生物学中的应用进展
2022-12-02张俊哲刘自勇马小清刘立成李福利
张俊哲,张 全,刘自勇,3,马小清,刘立成,3,李福利,3
(1.中国科学院 青岛生物能源与过程研究所,山东 青岛 266101;2.中国石化大连(抚顺)石油化工研究院,辽宁 大连 116045;3.山东能源研究院,山东 青岛 266101)
合成生物学是如今比较热门的生物科学的分支学科,主要是通过制造与组装生物功能元件、装置和系统,从而设计、改造细胞与生命体,使其获得符合人类需求的生物学功能,甚至实现创造全新的生命系统这一目标的学科[1]。合成生物学的分支合成代谢工程着眼于构建“自然界中不存在的生化系统”,它引入工程学理念,利用标准化、模块化的基因、酶等生物元件,重构细胞的代谢通路,从而高效地生产符合人类需求的代谢产物[2-3]。目前,合成生物学已取得丰硕的成果,如,稀有人参皂苷[4]、青蒿素[5]、番茄红素[6]的生物合成等。但是也应看到,合成代谢工程有时会面临着一些难题,例如,代谢通量不高、中间产物对细胞有毒性、存在内源性代谢的串扰和副反应等,无法预测性能,这些可能会阻碍其生产过程[7]。
在真核细胞中,由于细胞内复杂的内膜系统的存在,胞内空间被区域化与功能化,这使得相互区别的代谢反应能够同时进行,而不会发生串扰,使细胞能够实现复杂的生理功能。近些年来,在原核细胞中也发现了一些功能上类似于真核生物细胞器的结构——细菌微室 (bacterial microcompartments,BMCs) 便是其中之一。这是一种多面体形细胞器,由半透性的蛋白质外壳包裹着酶核心构成。在功能上,细菌微室外壳包封着核心酶与周围细胞质分离,可以保护细胞免受毒性代谢中间体的侵害,还可增大局部反应物浓度,防止副反应的发生,提高多步骤反应的反应通量。细菌微室在原核生物中的存在相当普遍,通过生物信息学比对,目前在19个细菌门类里发现了BMCs编码基因簇,且BMCs基因簇经常发生水平基因转移。细菌微室的一大特点就是外壳结构保守而功能多样,既可参与合成代谢,也可参与分解代谢[8]。细菌微室具有多样化功能,是原核生物代谢工程中重要的工具元件。
1 细菌微室的结构
细菌微室的结构可以分为两部分,蛋白质外壳与被外壳包裹的酶核心。其中外壳蛋白的结构非常保守,所有细菌微室的外壳都是由3类蛋白BMC-hexmer(BMC-H)、BMC-trimer(BMC-T)和BMC-pentamer(BMC-P)构成的。其中BMC-H含有1个Pfam00936结构域,在外壳中以同源六聚体的形式存在,而BMC-T含有2个串联的Pfam00936结构域,以三聚体的形式存在。BMC-H六聚体和BMC-T三聚体都是六边形,共同构成了多面体外壳的面的部分[9]。而BMC-P是五聚体,用于形成外壳顶点的部分[10]。外壳蛋白之间结合紧密,而且这种相互作用保守且通用,若将其中一个外壳蛋白的编码基因敲除,则该外壳蛋白的位置会被其他外壳蛋白组分所取代,并不会在外壳上形成缺口[11-12]。而在外壳蛋白聚体的中心存在孔道,可供底物与产物进出,外壳蛋白中心孔道对于不同小分子的通透性不同,且具有一定的选择性,这与细菌微室的功能相适配[13]。同时,在研究BMC-T的晶体结构时观察到中心孔道存在开启和关闭两种构象,这说明外壳蛋白可能存在变构机制,用来控制物质进出[14]。另外发现部分外壳蛋白的孔道中结合有Fe-S簇,推测这些外壳蛋白可能参与电子传递[11]。细菌微室及微室外壳蛋白结构示意见图1[15-17]。
构成细菌微室核心的酶通常带有封装肽 (encapsulation peptides,EPs),多数为15~20个氨基酸残基组成的两亲性α-螺旋,出现在封装蛋白的N端或C端,并通过一段连接肽与封装蛋白相连。封装肽参与细菌微室的组装,有些封装肽通过静电力与疏水作用力与外壳蛋白相结合[18],而另一些存在更加复杂的相互作用。研究发现,即使不存在外壳蛋白,封装肽也可以使封装蛋白发生凝聚,并形成具有催化活性的包涵体[19],如果去除封装蛋白的封装肽,封装蛋白就不再定位于细菌微室中[20]。目前对于封装肽的认识还不够全面,其在细菌微室的组装过程中发挥的作用仍未得到完全解析。
2 细菌微室的功能分类
细菌微室的功能十分多样,可以分为参与合成代谢的羧酶体和参与分解代谢的代谢体两种类型(图2)。羧酶体出现于蓝细菌和部分化能自养微生物中,羧酶体的核心蛋白主要是碳酸酐酶和1,5-二磷酸核酮糖羧化酶/加氧酶 (RuBisCO) 以及其他辅助蛋白,卡尔文循环中的CO2固定部分就在其中进行,外壳蛋白的存在可以提供一个高CO2浓度的微环境,减少O2的干扰[21]。
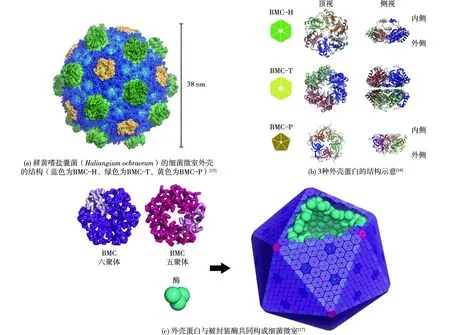
图1 细菌微室的结构示意Fig.1 Schematic diagram of the structure of BMCs
参与分解代谢的代谢体分布更为广泛,存在于多个细菌门类中,功能也极为多样化。尽管不同代谢体的底物和功能各不相同,但均含有一个构成基本相同的酶核心。该酶核心由特征酶(signature enzymes)、醛脱氢酶(aldehyde dehydrogenase,AldDH)、醇脱氢酶(alcohol dehydrogenase,AlcDH)和磷酸转酰基酶(phosphate acyltransferase,PTAC)组成。代谢体的作用模式可以概括为:底物在特征酶的作用下,裂解产生醛类;生成的醛经由醛脱氢酶和醇脱氢酶作用发生歧化,产生对应的醇和酰基-辅酶A;而酰基-辅酶A又在PTAC的作用下,生成酰基磷酸,产生的醇与酰基磷酸会离开代谢体,为细胞提供能量与碳源[8]。代谢体的外壳对代谢中间产物有着很好的包装能力,只有很少量的醛类从代谢体中逸出,远低于毒性浓度[11]。
特征酶是一系列具有底物特异性和反应特异性的醛生成酶,因为决定着代谢体的功能,故被称为特征酶。发现最早的一类特征酶是辅酶B12依赖的裂解酶,它含有多个亚基,需要腺苷钴胺素作辅因子,这类酶以1,2-丙二醇或乙醇胺为底物,可以将1,2-丙二醇脱水形成丙醛或将乙醇胺裂解形成乙醛和氨。由于这类裂解酶很容易由于副反应而失活,以及辅酶B12的稀缺性,拥有这类特征酶的代谢体还含有特征酶复活酶和辅酶B12合成酶[22]。另一大类特征酶属于甘氨酰自由基酶 (glycyl radical enzymes,GREs),它不需要辅酶B12作为辅因子,而是通过甘氨酰自由基 (Gly·) 来催化反应的发生,含有这类特征酶的代谢体也被称为GRMs(GRE-associated microcompartments,GRMs),这是代谢体中数量最多的一类[23]。根据其基因座的结构,GRMs又可以分为GRM1~GRM5等5种类型。其中,GRM1和GRM2基因座分别编码Ⅰ型和Ⅱ型胆碱裂解酶,可以将胆碱裂解为乙醛和三甲胺;GRM3和GRM4则编码1,2-丙二醇脱水酶;而GRM5除了编码1,2-丙二醇脱水酶外,还编码1个醛缩酶和1个乳醛还原酶[23]。GRM5参与L-岩藻糖或L-鼠李糖的降解,L-鼠李糖先在胞质中转化为L-鼠李酮糖-磷酸 (L-rhamnulose-P),L-鼠李酮糖-磷酸进入GRM5后,在醛缩酶作用下生成(S)-乳醛与磷酸二羟丙酮,后者会离开代谢体,而(S)-乳醛会在乳醛还原酶的作用下,还原为1,2-丙二醇,再在1,2-丙二醇脱水酶作用下,生成丙醛,最终被代谢为丙醇与丙酸,L-岩藻糖的代谢方式与L-鼠李糖的代谢方式一致[24-25]。GREs的甘氨酰自由基是在蛋白翻译后由GREs特异的激活酶 (AEs) 催化产生的,但具体过程尚未完全阐明。
除了上述得到表征的代谢体类型之外,近年来还通过宏基因组、宏转录组等手段发现了不少全新的代谢体类型,不过这些代谢体尚未得到全面的表征,很多只是推测存在功能[26-27]。其中一类新发现的代谢体称为糖磷酸利用(sugar-phosphate utilization,SPU)代谢体,在超过20个细菌门类中都发现其编码基因座,是分布最广的BMCs类型之一。这类代谢体基因座中含有2个糖磷酸加工酶基因,可以代谢磷酸脱氧核糖和5′-磷酸核糖,表明这类代谢体的潜在功能是代谢DNA的降解产物,用来回收环境中核酸碎片的生物质[26]。还有一类新发现的代谢体可能参与芳香类化合物的代谢,被称为ARO代谢体,其基因座中编码1个开环加氧酶,推测可能的底物为2-氨基苯酚。该类代谢体的醛脱氢酶非常特别,在进化树中单独聚类,而且其外壳蛋白也很特殊,只包含2种BMC-H和1种BMC-P蛋白,是所有细菌微室中最简单的[26]。另一类功能未知的BMCs称为BUF1 (BMCs with unknown function),推测其参与分解代谢,但基因座中不存在特征酶编码基因[28],近年来通过调控基因和转运蛋白基因等辅助基因功能分析,推测其可能参与嘌呤(黄嘌呤、次黄嘌呤)的降解[27]。但以上代谢体的功能都是根据基因座结构分析推测得出,尚待全面地表征。
3 细菌微室的组装
3.1 羧酶体的组装
相比于代谢体,羧酶体的组装研究得较为清晰。羧酶体可分为两类,α-羧酶体和β-羧酶体,其组装方式并不相同。β-羧酶体的组装是先完成核心的组装,之后再由外壳蛋白进行封装。而α-羧酶体的核心与外壳的组装则是同时进行。
β-羧酶体的组装过程研究得最为清晰(图3),其组装由CcmM蛋白引发,CcmM含有C端串联排列的3到5个RuBisCO小亚基同源结构域 (small subunit-like domains,SSLDs) 和N端的γ-碳酸酐酶结构域两部分[29-30](部分羧酶体的CcmM缺失碳酸酐酶结构域,称为截短CcmM)。SSLDs可代替RuBisCO小亚基掺入RuBisCO中,从而引发RuBisCO的聚集与沉淀,形成前羧酶体[31](procarboxysomes,PCs)。CcmM的N端碳酸酐酶结构域可与另一个保守的结构蛋白CcmN的N端结构域结合,而CcmN的C端具有封装肽,可与外壳蛋白相互作用,从而引发外壳蛋白的组装。最后与属于BMC-P的外壳蛋白CcmL结合,形成羧酶体多面体形外壳的顶点,从而完成封装,并实现成熟羧酶体从前羧酶体的出芽[31]。

γ-CA—γ-碳酸酐酶,即CcmM;SSLDs—RuBisCO小亚基同源结构域;CcmN—结合外壳蛋白的结构蛋白;CcmK2—外壳蛋白BMC-H;CcmO—外壳蛋白BMC-T;CcmL—外壳蛋白BMC-P图3 β-羧酶体的组装过程[31]Fig.3 The process of β-carboxysome assembly[31]
α-羧酶体的结构蛋白与β-羧酶体的结构蛋白存在差异,组装过程也不同。根据冷冻电镜的观察结果,α-羧酶体的核心与外壳蛋白的组装同时进行[32]。α-羧酶体的组装机制目前仍不甚明了,但已经发现是由一个保守的固有无序蛋白CsoS2引发。CsoS2的序列可分为3个区段:N端、中段、C端,如果C端区段缺失,则不能形成完整的羧酶体结构[33]。目前推断3个区段的功能分别为:N端区段可招募外壳蛋白CsoS1,提高其局部浓度,引发其自组装;中段区段结合RuBisCO;C端区段可结合外壳蛋白,从而将聚集的酶核心锚定在外壳蛋白上[33-34]。
3.2 代谢体的组装
由于构成代谢体的酶种类较多且复杂,还有很大的差异性,代谢体的组装过程目前仍有很多未知之处,其大体框架是封装肽使得酶聚集,并沉淀形成酶核心,同时封装肽与外壳蛋白结合,实现酶核心的封装。为实现代谢体相较于羧酶体更为复杂的功能,代谢体的外壳也承担着底物选择、多种酶的结合、电子传递等多样的职责,因此,代谢体中含有更多种类的外壳蛋白,例如,沙门氏菌的PDU (丙二醇利用)代谢体就含有PduA、PduBB’、PduJ、PduK、PduN、PduT、PduU共7种外壳蛋白[35],这就带来了不同外壳蛋白如何正确装配的问题。对于这一问题也有相关研究,证明了PDU代谢体的外壳的装配是由PduA和PduJ引发的,并提出了外壳装配模型[36](图4)。PduA和PduJ这两个外壳蛋白与其他外壳蛋白的一个不同之处是其具有强的自组装倾向,异源表达PduA或PduJ时,会在宿主细胞中形成空心管状结构,这是由于外壳蛋白之间以一定的夹角相互结合。正是这种带有夹角的结合方式,形成了代谢体多面体结构的棱。而其余外壳蛋白不具有自组装能力,而是掺杂进由PduA和PduJ自组装形成的框架中,赋予其丰富的功能,并改变了自组装结构的形状,使其不再形成管状。其他的六边形外壳蛋白可以与PduA和PduJ平直地结合,从而形成平面,而属于BMC-P的PduN会与周围5个PduA和PduJ结合,并形成多面体的顶点。棱、平面、顶点这3种局部结构,最终会使得多面体形的代谢体结构得以形成。
4 细菌微室的表达调控
羧酶体参与CO2的固定,对自养微生物的生存至关重要,因此羧酶体基因在这类微生物中为组成型表达。而代谢体只在相应底物存在时才会形成,其基因座受底物诱导表达。细菌微室基因座的转录调节系统可分为两类,一类是单组分的转录调节系统,另一类是双组分的转录调节系统[27],结果见图5。
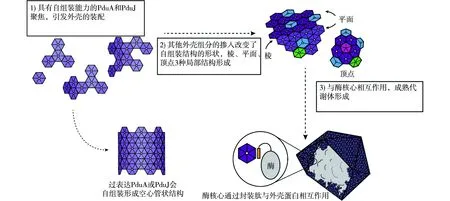
图4 沙门氏菌PDU代谢体外壳的装配机制模型[36]Fig.4 Model of the assembly mechanism of the PDU metabolosome shell in Salmonella enterica[36]
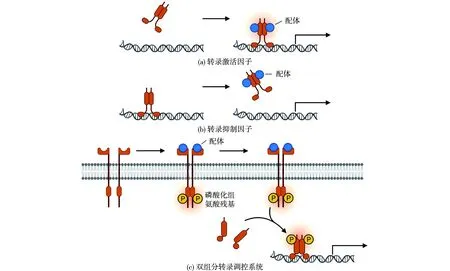
图5 BMCs基因座的调控方式[24]Fig.5 Transcriptional regulation types associated with BMCs loci[24]
单组分的转录调节系统又可细分为转录激活因子和转录抑制因子。这类转录因子含有1个N端DNA结合结构域和1个C端配体结合结构域,配体的结合会改变转录因子对DNA的亲和力。对转录激活因子而言,配体结合会触发转录因子与基因座启动子的结合,并招募转录相关蛋白。目前在BMCs基因座中发现的转录激活因子家族包括PucR、PocR、LysR、AraC/XylS、Crp/Fnr等。而转录抑制因子与配体结合后会与启动子分离,解除转录抑制,从而起始转录过程。在BMCs基因座中发现的转录抑制因子家族包括DeoR、TetR和GntR。转录因子的配体通常为代谢体的底物,例如,丙二醇利用微室的调控基因pocR可以被1,2-丙二醇特异性地激活[37],而乙醇胺利用(ethanolamine utilization,EUT)微室的调控基因eutR可以被乙醇胺激活[38]。
而双组分的转录调节因子包括两部分:受体组氨酸激酶和应答调控因子。受体组氨酸激酶可以具有跨膜结构域,定位于细胞膜上,在胞外侧有1个配体结合结构域,而胞质侧有1个激酶结构域。当结合配体(代谢底物)后,会在1个保守的组氨酸残基上发生自磷酸化,随后该磷酸基会转移至应答调控因子的保守天冬氨酸残基上,而被磷酸化的应答调控因子会激活BMC基因座的转录。而大部分受体组氨酸激酶并不具有跨膜结构域,这类受体组氨酸激酶定位于胞质中,结合由转运蛋白运送到胞质中的配体。
5 细菌微室在合成生物学中的应用
5.1 细菌微室直接作为代谢元件
由于细菌微室的保守性以及在自然界中经常发生基因簇的水平转移,因此细菌微室很适合作为通用的即插即用的代谢元件。此技术已有不少成功先例,例如,将弗氏柠檬酸杆菌(Citrobacterfreundii)的pdu基因座在大肠杆菌中表达,可以使得大肠杆菌具有利用1,2-丙二醇的能力[39]。同样的,羧酶体也可以异源表达,例如,将可化能自养的那不勒斯嗜盐杆菌(Halothiobacillusneapolitanus)的羧酶体基因座在大肠杆菌中表达,可以在大肠杆菌中产生与羧酶体非常相似的结构,该结构在体外实验中表现出了固定CO2的活性[40]。此外,不只是原核生物,羧酶体还可以在真核细胞中表达。将蓝细菌的β-羧酶体基因座导入烟草的叶绿体中,可以在叶绿体中观察到类似羧酶体的结构产生。将宿主原有RuBisCO敲除后发现,导入了β-羧酶体基因座的烟草依旧可以生长,并且比野生型具有更高的CO2固定效率,表明在叶绿体中异源表达的羧酶体能够正常发挥功能,相比原本的RuBisCO效率更高[41]。
5.2 细菌微室的外壳蛋白改造
由于细菌微室的外壳蛋白高度保守,且外壳蛋白间的相互作用是通用的,因此异源的细菌微室外壳蛋白可以用于构建功能正常细菌微室。例如,将沙门氏菌的PDU微室外壳蛋白PduA替换为EUT微室外壳蛋白EutM,嵌合体微室不仅结构正常,以1,2-丙二醇为碳源时,菌株的生长速率还有了明显提升[42]。不同外壳蛋白对可通过的小分子选择性存在差异的原因,是由于外壳蛋白孔道的大小和电荷情况不同,因此可以通过对孔道周围的氨基酸残基进行突变,调节外壳蛋白的通透性,从而调节细菌微室的性能[42]。还可以通过类似的手段,赋予外壳蛋白全新的功能,例如,将一种BMC-T外壳蛋白设计成在其孔中结合具有氧化还原活性4Fe-4S簇,该簇通过氧化还原循环保持稳定,并使得该外壳蛋白表现出类似于低电位细菌铁氧还蛋白的特性[43]。
5.3 人工构建新型细菌微室
人工构建新型细菌微室的工作也在深入进行中。在前期的概念验证实验中,将绿色荧光蛋白(GFP)与细菌微室核心酶的封装肽融合,可以在宿主菌中观察到点状的荧光,证明GFP被成功导入细菌微室内[44]。利用相同的原理,在大肠杆菌中表达来源于弗氏柠檬酸杆菌的PDU微室外壳蛋白以及连接有封装肽的来源于运动发酵单胞菌(Zymomonasmobilis)的丙酮酸脱羧酶(pdc)和醇脱氢酶(adh)基因,可以将两种酶封装入细菌微室中,从而构建出能够合成乙醇的人造细菌微室,并且乙醇合成活性相比于游离酶的合成活性更高[45]。与之类似,将人工构建的融合有封装肽序列的多磷酸盐激酶(ppk1)基因与编码细菌微室外壳蛋白的操纵子在大肠杆菌中共表达,可以在细菌微室中大量积累多磷酸盐,同时不对细菌生长造成太大影响,这表明细菌微室具有作为胞内毒分子储蓄池的潜力[46]。由于封装肽与外壳蛋白的互作机制尚不明确,也可以利用原理更加清晰、可控性更好的蛋白质组装SpyTag-SpyCatcher系统将外源蛋白导入细菌微室中。可以在外壳蛋白上融合表达SpyCatcher蛋白,而在外源蛋白上连接SpyTag序列,SpyCatcher蛋白会识别SpyTag序列,并且两者之间会在特定的氨基酸残基上通过侧链形成酰胺键,从而将外源蛋白共价连接在外壳蛋白上[47-48]。细菌微室外壳蛋白的半渗透性可以对气体起效,理论上可以利用其解决产酸梭菌利用合成气发酵时胞内CO供应不足的问题[49-50]。由于细菌微室具有提高局部反应物浓度以及减少有毒代谢中间产物扩散的特性,同时还可以通过是否添加诱导物来启停相关基因的表达,因此,以细菌微室为基础构建的生物反应器可以具有高效率以及高灵活性的特点,有着巨大的应用潜力[51]。
6 结语
细菌微室在自养生物的CO2固定和异养生物的有机底物分解代谢中有着至关重要的作用,这类细胞器提升了细菌的代谢多样性,并为细菌提供在特定的生境中的竞争优势。在过去十多年中,对细菌微室结构与功能方面的研究取得了较大进展,这不仅加深了对细菌微室生理功能的认识,还使得将其应用于合成生物学成为可能。而细菌微室展现出强大的可操作性,使得其在合成生物学中的应用具有广阔的前景与巨大的潜在价值。
