黄河源区季节性湖泊湿地退化对土壤微生物碳源利用的影响
2022-12-01姚步青赵之重韦鎔宜张永超秦文萍
段 鹏,姚步青,赵之重,3*,韦鎔宜,张永超,秦文萍, 王 婷
(1.青海大学,青海 西宁 810016;2.中国科学院西北高原生物研究所,青海 西宁 810008;3.青海民族大学,青海 西宁 810007;4.青海省畜牧兽医科学院,青海 西宁 810016)
季节性湖泊湿地属于湖泊湿地的二级分类湿地,由面积较小的(1 km2)季节性积水的湖沼组成,是青藏高原高寒湿地的重要组成部分[1],对我国的生态安全具有非常重要的保护作用[2]。但由于全球变暖与人类不合理放牧,青藏高原高寒湿地不断发生退化[3]。高寒湿地退化是以水分流失为驱动的演替过程,其退化过程为从高寒湿地到沼泽化草甸,再到退化草甸,最后退化为“黑土滩”[4-5]。湿地退化过程中湿地面积减少,湿地呈斑块化分离,湿地植物群落组成发生退化,土壤物理结构遭受破坏,土壤养分大量流失[6]。生境的改变会引起土壤微生物对碳源利用能力的变化[7-8],而微生物对碳源的利用能力可以作为土壤质量和土壤肥力变化的重要指标[9]。因此研究湿地退化对土壤微生物碳源利用的影响以及土壤微生物碳源利用能力与不同环境因子的关系,对于湿地退化研究具有重要意义。
土壤微生物作为湿地土壤的重要组成部分,在维持湿地土壤生态系统的生态稳定等方面起着重要作用[10]。湿地土壤微生物是由多种类微生物组成的结构极为复杂的微生物群落[11-12],土壤微生物碳源利用受到土壤微生物群系结构、种类、固有性质的影响[13]。土壤微生物对不同类型碳源的利用不仅会影响土壤微生物活性还会影响其功能多样性,微生物对碳源的利用能力以及微生物功能多样性作为关键性土壤微生物生态指标,可以敏感地反映出土壤质量和健康状况[14-16],对探索湿地退化对土壤微生物群落的影响,了解湿地生态系统结构和功能具有重要意义[17-18]。李慧等人[19]对草原的土壤微生物群落特征研究发现,不同根系微生物对碳源的利用能力有差异,该差异主要受微生物对氨基酸类和碳水化合物类碳源利用变化的影响。翁晓红等人[20]对三江平原小叶章湿地研究发现,小叶章湿地土壤微生物对胺类碳源的相对利用率最高。段鹏等人[21]通过对高寒河流湿地退化研究发现,随着高寒湿地的退化,其土壤微生物对碳源的利用强度下降,其中对脂类碳源的相对利用率最高,脂类碳源为优势碳源。因此,研究季节性湖泊湿地退化过程中,土壤微生物群落功能多样性和微生物对各类碳源利用情况的变化,可以了解不同退化阶段土壤微生物群落碳源利用的差异及其与各环境因子间的关系,为调控季节性湖泊湿地微生物群落结构和功能提供理论依据[22]。黄河源区地处中国青海省三江源自然保护区内,属于内陆高寒湿地,其湿地资源对当地的生态环境、气候变化、水质净化以及下游水系水质具有重要影响[21]。但有关黄河源区季节性湖泊湿地退化对土壤微生物碳源利用的影响以及不同环境因子和土壤微生物碳源利用的关系等方面仍不明确。
Biolog微平板分析法作为一种操作简单快捷,并能准确地获取微生物群落碳源利用和结构与功能多样性等相关信息的研究方法,可以分析土壤微生物对不同碳源的利用能力和土壤微生物多样性变化规律[23-24],已被广泛应用于土壤生态系统研究。因此,本研究以黄河源区退化季节性湖泊湿地为研究对象,采用空间代替时间的方法,利用Biolog-Eco法研究季节性湖泊湿地不同退化阶段土壤微生物结构与功能多样性以及土壤微生物碳源利用情况,解析季节性湖泊湿地退化对土壤微生物功能多样性、土壤微生物群系结构和土壤微生物碳源利用的影响,以期揭示季节性湖泊湿地过程中土壤微生物群落碳源利用差异及其与各环境因子的关系,为黄河源的季节性湖泊湿地退化监测和近自然修复提供科学理论依据。
1 材料与方法
1.1 试验区自然概况
试验区(33°50′~35°40′N,96°55~99°20′E)位于中国青海省三江源自然保护区内[21],地处果洛州玛多县境内,属高寒草原气候,平均海拔4 400 m以上,昼夜温差与年温差较大[25],除植被生长季以外,各月平均温度处于0℃以下。有一条河流自北向南流过试验区,并在试验区内形成由湖泊湿地组成的湿地核心区[21],核心区的湖泊湿地主要是由面积较小(<1 km2),且在雨季能得到部分水源补充的季节性湖沼组成的。
1.2 样点布设及采样概况
2018年5月份,在实验区内,采用空间代替时间的方法,根据水分梯度和植被群落变化特征,以核心区的季节性湖泊为中心向外围出发,垂直距离湖泊湿地由近及远,设置3条重复样线。参考李飞[26]、李成一[27]、李希来[4]和林春英[3]等人对季节性湖泊湿地退化的划分,从湖泊湿地中心向外围延展将退化季节性湖泊湿地划分为4个阶段,且第4阶段为非湿地景观,并采用Gao等方法和高寒沼泽湿地退化等级标准(DB 63/T 1794-2020)中对湿地退化的划分[21,28-29],结合本实验样地的实际地形及环境状况,每条重复线上设立4个退化样点,将季节性湖泊湿地退化人为分为未退化湿地(湿地)、轻度退化湿地(沼泽化草甸)、中度退化湿地(退化草甸)、重度退化湿(退化草原)。
2018年8月份对所划分的季节性湖泊湿地不同退化阶段,采用0.5 m×0.5 m样方框进行植被群落取样,测定植被高度、盖度和生物量,并进行三次重复测量。测定土壤湿度并使用土钻(直径5 cm)采样不同退化阶段0~10 cm土层的土壤,每个退化阶段区域内随机选取5个采样点进行采样,每个样点随机采集3钻土壤[21],并将3钻土合并为一个土壤样品[21],去除植被根系和碎石,土壤样品及时送回实验室4℃保存。
1.3 试验方法
1.3.1植被处理 采用目测法对每个样方框内的植被进行覆盖度测定,将样方框内植被地上部分齐根剪下,使用烘干箱进行烘干处理(65℃,48 h),计算植被地上生物量[21]。使用根钻采集0~10 cm土层中的植被根系,去除杂草、泥土和碎石等,清洗烘干(65℃,48 h),计算植被地下生物量。
1.3.2土壤处理 运用重铬酸钾-内稀释热法对土壤有机碳测定,使用流动注射分析仪(SEAL Auto Analys-er 3,德国)对土壤全氮、全磷含量进行测定,采用Top Cloud-agri WET速测仪对土壤含水量和电导率进行测定[21]。
1.3.3土壤微生物功能多样性测定 Biolog微平板分析法作为一种研究微生物群落功能多样性的常用方法,其主要通过分析土壤微生物对各类碳源的代谢功能,进而获得土壤微生物群落对碳源的代谢能力等相关数据[30],Biolog-Eco微平板为标准的96孔板,板内包含3组重复,每组中包含31个含有不同单一碳源的培养基孔和1个空白对照孔[21]。土壤微生物功能多样性测定的具体实验过程:称取10 g鲜土和90 mL 0.85%无菌NaCl溶液倒入锥形瓶中,150 r·min-1条件下震荡30 min,后静置20 min。用移液枪吸取150 μL的释至10-3的土壤悬浮液接种到Biolog-Eco板中[29],置于25℃恒温培养箱连续培养240 h,并使用酶标仪在590 nm条件下每24 h测定各孔吸光值[21]。
1.4 数据处理
公式计算如下[21,31]:
AWCD=[∑(C-R)]÷N
(1)
公式中AWCD为土壤微生物平均颜色变化率(Average well color development),C为单一碳源孔的吸光值,R为对照孔的吸光值,N为所分析的碳源数量[21,32]。
(2)
公式中U为土壤微生物Mclntosh指数,ni为第i孔的相对吸光值[21]。
H′=-∑PilnPi
(3)
公式中H′为土壤微生物Shannon-wiener物种丰富度指数,Pi为第i个孔的相对吸光值/所有孔的相对吸光值之和[21]。
碳源相对利用率=恒温培养168 h下的某一类型碳源AWCD之和/31种碳源的AWCD值之和×100%。Biolog生态板中的31种碳源根据其化学基团的性质的不同大体分为六大类:单糖/糖苷/聚合物类、醇类、氨基酸类、酯类、胺类和酸类[33]。
样地基本数据采用Excel进行数据分析,采用SigmaPlot分析成图,采用Origin进行主成分和冗余分析,运用SPSS进行单因素方差分析(One-way ANOVA)[21],利用Pearson进行相关性分析。
2 结果与分析
2.1 不同退化阶段植被及土壤特征
季节性湖泊湿地退化过程中植被基础数据见表1,随季节性湖泊湿地退化加重,其植被盖度、地上生物量和地下生物量显著下降(P<0.05)。重度退化湿地与未退化湿地相比,植被盖度下降了33.33%,地上生物量下降了82.82%,地下生物量下降了85.16%。

表1 不同退化阶段植被基本群落特征的变化
季节性湖泊湿地退化过程中土壤基本理化性质见表2,季节性湖泊湿地退化过程中,其土壤全氮、全磷含量和含水量均呈先升高后下降趋势,且变化显著(P<0.05),而土壤电导率显著降低(P<0.05)。重度退化湿地与未退化湿地相比,土壤全氮含量下降了45.61%、全磷下降了15.00%、有机碳含量下降了23.29%,含水量下降了42.23%。

表2 不同退化阶段土壤理化性质变化
2.2 不同退化阶段土壤微生物群落碳源利用总体特征
土壤微生物群落AWCD值可以反映土壤微生物群落的活性和土壤微生物群落的碳源利用强度[34-36]。季节性湖泊湿地不同退化阶段的AWCD值如图1所示。由图1A可知,在培养过程中,0~24 h土壤微生物AWCD值变化较小,24~96 h土壤微生物AWCD值迅速增长,168 h后土壤微生物的AWCD值逐渐稳定。因此本研究通过对168 h培养时间下季节性湖泊湿地各退化阶段的AWCD值进行分析。由图1B可知,随季节性湖泊湿地退化,土壤微生物活性呈先升高后下降趋势,其中轻度退化湿地AWCD值显著高于其他退化阶段(P<0.05),而重度退化湿地AWCD值显著低于其他退化阶段(P<0.05)。
2.3 不同退化阶段土壤土壤微生物多样性特征
本研究采用Mclntosh指数、Shannon指数反映季节性湖泊湿地退化过程中土壤微生物均匀度和丰富度的变化[8,37],168 h培养时间下,季节性湖泊湿地不同退化阶段的土壤微生物均匀度和丰富度的分析结果如图2所示。由图2A可知,季节性湖泊湿地各退化阶段土壤微生物均匀度从24 h培养时间开始,随培养时间的增长而不断提高,到168 h增长减缓趋于平稳。由图2B可知,轻度退化湿地土壤微生物均匀度显著高于重度退化湿地(P<0.05),随季节性湖泊湿地退化加重,其土壤微生物均匀度整体上呈现先升高后下降趋势。由图2C可知,季节性湖泊湿地土壤微生物丰富度随培养时间增长无明显变化。由图2D可知,轻度退化湿地土壤微生物丰富度显著高于重度退化湿地(P<0.05),随季节性湖泊湿地退化加重,其土壤微生物丰富度整体上呈现先升高后下降趋势。
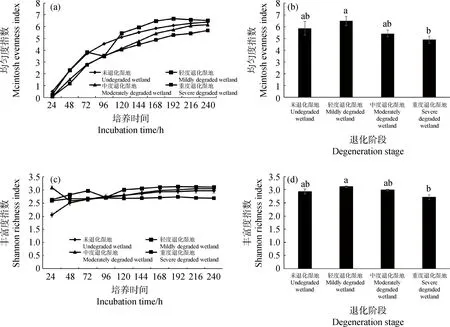
图2 季节性湖泊湿地不同退化阶段土壤微生物群落功能多样性
2.4 不同退化阶土壤微生物对不同类型碳源利用特征
选取AWCD值趋于平稳的168 h培养时间下的季节性湖泊湿地不同退化阶段土壤微生物对不同碳源的利用能力进行分析,如表3所示,季节性湖泊湿地退化使土壤微生物对单糖/糖苷/聚合物类、酯类、醇类、胺类碳源的相对利用率先增加后减少,重度退化湿地与未退化湿地相比,土壤微生物对单糖/糖苷/聚合物类碳源的相对利用率下降了93.33%,酯类下降20.00%,醇类下降42.71%,胺类下降36.59%。其中土壤微生物对酯类碳源的相对利用率始终高于其它碳源,土壤微生物对酸类碳源的相对利用率随退化的加重而增长,重度退化湿地微生物对酸类类碳源的相对利用率与未退化湿地相比增加了32.69%。随季节性湖泊湿地退化的加重,土壤微生物对单糖/糖苷/聚合物类碳源的相对利用率变化趋势与土壤生物群落碳源利用总体特征的变化趋势相似,均呈现先升高后下降趋势。
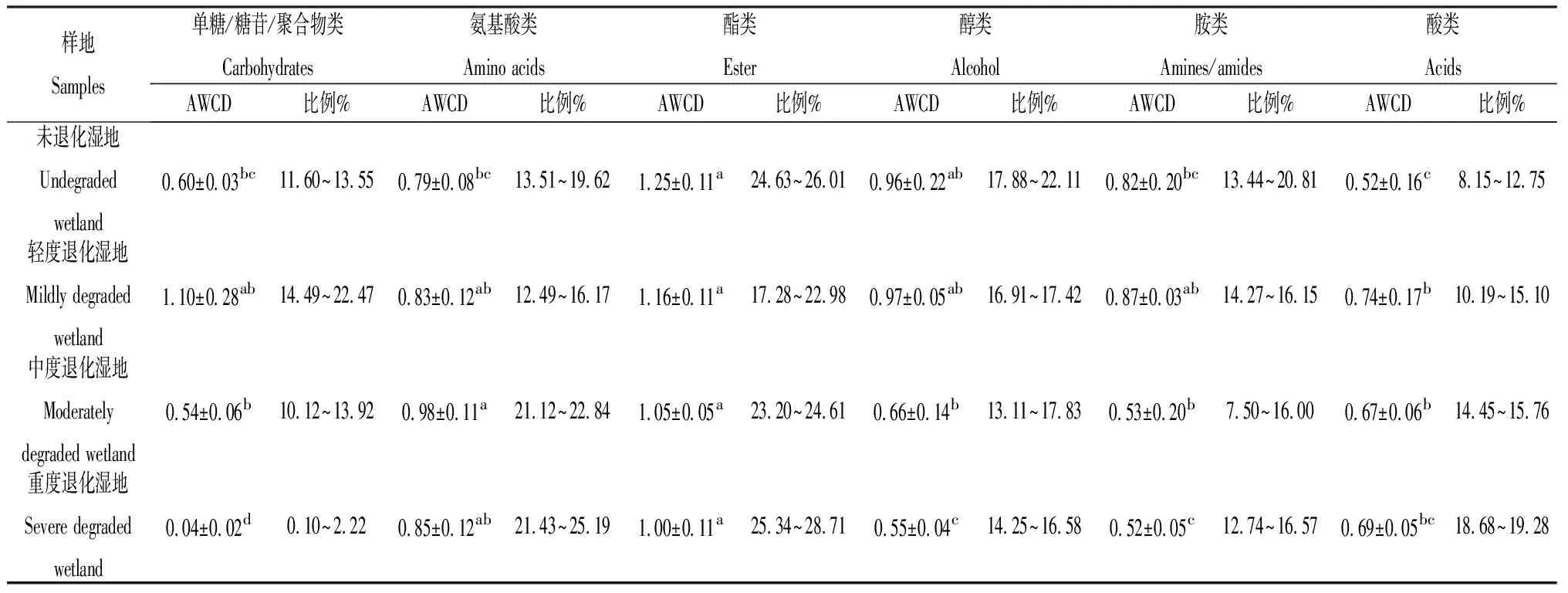
表3 季节性湖泊湿地不同退化阶段土壤微生物对各类碳源的利用情况
2.5 退化湿地土壤微生物群落主成分分析
选取AWCD值趋于平稳的168 h培养时间下季节性湖泊湿地不同退化阶段土壤微生物对不同碳源的利用率进行主成分分析,提取了2个主成分,第一主成分(PC1)为51.5%,第二主成分(PC2)为23.9%,选取第一、二主成分作图来表征不同退化阶段土壤微生物对六大类碳源的利用状况,各退化阶段之间距离大小可以表示其土壤微生物对六大碳源的利用状况的相似度,距离越小其利用状况的相似度越高。如图3所示,未退化阶段和轻度退化阶段分布在一个区域,离散性较小,中度退化阶段和重度退化阶段分布在一个区域,离散性较小,但两组间分布在不同区域,离散性较大,表明未退化湿地和轻度退化湿地土壤微生物对碳源的利用状况相近,中度退化湿地和重度退化湿地土壤微生物对碳源的利用状况相近,但未退化湿地和轻度退化湿地与中度退化湿地和重度退化湿地土壤微生物对碳源的利用状况差距较大。

图3 不同退化阶段土壤微生物对不同碳源利用主成分分析
2.6 退化湿地土壤微生物对碳源利用与环境因子之间的冗余分析
为进一步了解不同退化阶段土壤微生物群落对各类碳源的利用情况与环境因子的关系,本研究对土壤微生物AWCD和其相应的基本植被与土壤理化数据进行冗余分析。首先对各环境因子进行了方差膨胀因子分析(表4),分析结果表明各环境因子之间具有较高的相关性,根据分析结果剔除方差膨胀因子较高的土壤含水量、植被地上生物量和地下生物量等环境因子。重新进行共线性检测,共线性检测结果见表5,各环境因子的方差膨胀因子小于10,平均方差膨胀因子小于5。已有研究表明,膨胀因子大于10的环境因子不进入RDA分析[38-39],因此土壤含水量、植被地上生物量和地下生物量环境因子不进入冗余分析。分析结果如图4所示,1轴和2轴的解释变量特征值分别为83.09%和11.74%,说明季节性湖泊湿地退化过程中的基础土壤和植被指标对土壤微生物碳源利用类型影响较大。土壤全氮含量和植被盖度与土壤微生物对糖类和醇类碳源的利用率呈显著正相关关系(P<0.05)。但根据土壤微生物对各类碳源的利用情况与环境因子之间的相关系分析可知(表6),土壤含水量与糖类、酯类、醇类和胺类碳源的利用有显著相关性。通过各环境因子之间的相关性分析可知(表7),含水量与全氮含量、全磷含量、电导率和植被盖度显著正相关。
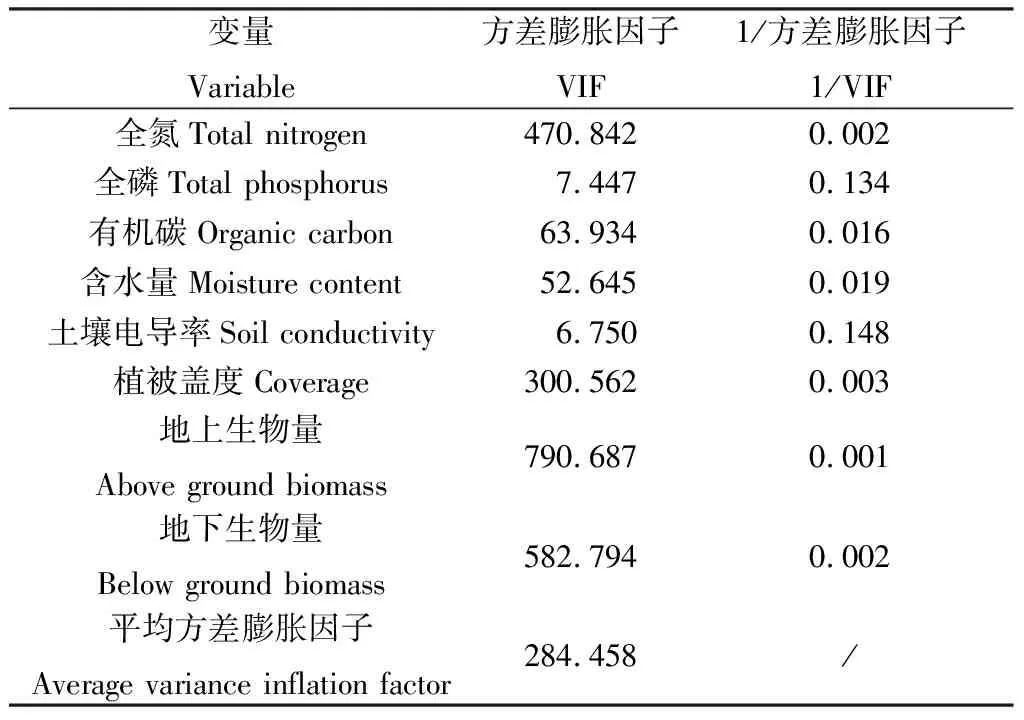
表4 多重共线性检验结果

表5 多重共线性检验结果

表6 环境因子之间的相关性
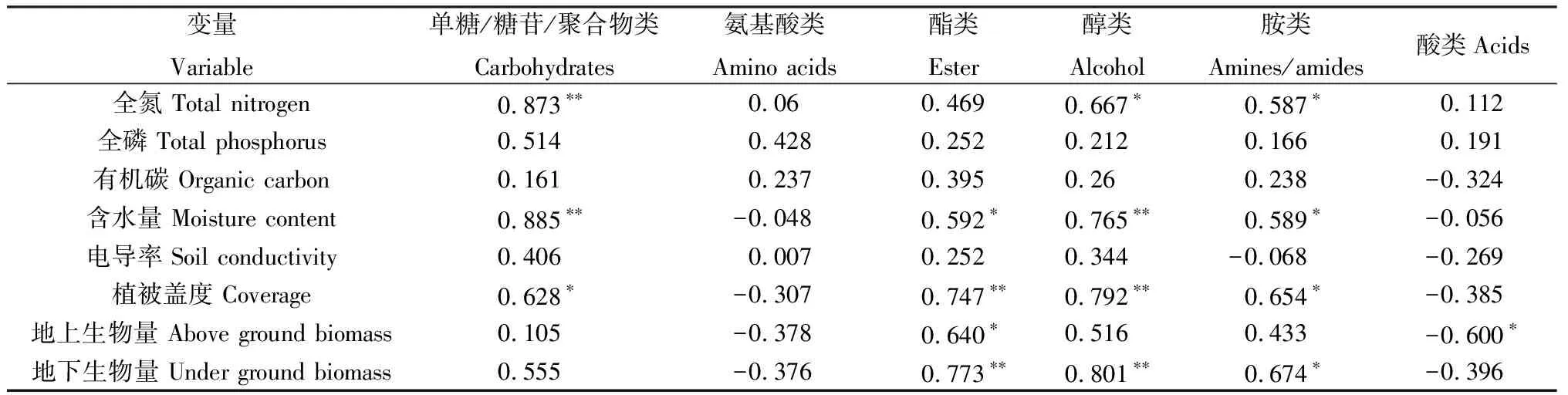
表7 土壤微生物对各类碳源的利用情况与环境因子之间的相关系分析

图4 不同退化阶段土壤微生物对碳源利用和环境因子的冗余分析
2.7 退化湿地土壤微生物群落特征与环境因子之间的相关性分析
黄河源区季节性湖泊湿地退化过程中各环境因子对土壤微生物功能多样性的影响见表8,土壤全氮含量、土壤含水量和植物盖度与土壤微生物AWCD值和土壤微生物均匀度呈显著正相关关系(P<0.05),土壤全氮、全磷含量和土壤含水量与土壤微生物丰富度呈显著正相关关系(P<0.05)。

表8 平均颜色变化率、均匀度、丰富度与环境因子之间的相关性分析
2.8 退化湿地土壤微生物群落特征与其对碳源利用情况相关性分析
黄河源区季节性湖泊湿地退化过程中,土壤微生物利用各类碳源的能力对其功能多样性的影响见表9,在季节性湖泊湿地退化过程中,土壤微生物对单糖/糖苷/聚合物类和醇类碳源的利用与土壤微生物AWCD值、土壤微生物均匀度和土壤微生物丰富度显著正相关(P<0.05)。且土壤微生物对单糖/糖苷/聚合物类碳源的利用与土壤微生物功能多样性具有极显著的相关性(P<0.01)。即不同退化阶段微生物功能多样性差异主要体现在对单糖/糖苷/聚合物类碳源和醇类碳源的利用上,其中土壤微生物对单糖/糖苷/聚合物类碳源的利用能力尤为突出。

表9 平均颜色变化率、均匀度、丰富度与土壤微生物对碳源利用之间的相关性分析
2.9 不同退化阶段土壤微生物代谢指纹数据聚类分析
1991年,代谢指纹技术首次被应用于微生物群落分析,微生物代谢指纹可以通过分析土壤微生物群系的特性,有效地区分不同退化程度的土壤微生物群系在时间和空间的变化以及相似度的关系。聚类分析图谱可以直观显示出各退化阶段土壤微生物对不同种类碳源的利用情况,并将利用状况相似的样点聚为一类[8]。
由于168 h培养时间下微生物AWCD值趋于平稳,因此将接种土壤悬液的Biolog-Eco板168 h培养时间下的数值做聚类分析,并形成树状图。由图5可知,当聚合水平大于10时将季节性湖泊湿地不同退化程度的土壤分为两类。第一类包括未退化湿地、轻度退化湿地和中度退化湿地,第二类包括重度退化湿地。重度退化湿地与未退化湿地、轻度退化湿地和中度退化湿地较长的距离尺度相聚,说明重度退化湿地微生物群系结构与其他退化阶段相比产生明显改变。

图5 季节性湖泊湿地不同退化阶段土壤微生物群系代谢指纹聚类分析图谱
3 讨论
本研究发现,在黄河源区季节性湖泊湿地退化过程中,随着植被盖度、生物量、土壤全氮含量、全磷含量、有机碳含量和含水量的显著下降(P<0.05),土壤微生物对碳源利用的强度以及微生物群落功能多样性发生改变,其土壤微生物对碳源的利用能力以及活性整体呈下降趋势。这与刘燕等[32]对黄河源区黑土滩退化的研究结果相近,说明湿地退化过程中土壤微生物对碳源的利用能力等特征受多环境因子的影响。对湿地不同退化阶段的土壤微生物对碳源的利用能力与环境因子相关性分析可知,植被盖度和土壤含水量与土壤微生物对碳源的利用能力呈显著正相关关系,其主要原因是植被盖度下降导致土壤呼吸增强、土壤含水量下降[40],从而影响土壤微生物的碳源代谢[41]。已有文献表明,土壤水分下降,土壤物理结构受到破坏,在风蚀的影响下土壤大团聚体被破坏,大量有机碳流失与分解[42-44],土壤有机碳作为土壤微生物可利用的碳源[45],其流失影响土壤微生物对碳源的利用。李飞等[26]对土壤微生物碳代谢功能多样性影响的研究中也发现湿地全氮含量、全氮/全磷、含水量和植被盖度可以显著影响微生物活性和其碳源代谢。已有研究表明,土壤微生物的丰富度受到土壤养分、含水量、容重和植物盖度等因素的影响[46],但本研究发现黄河源区季节性湖泊湿地退化过程中,植被盖度对其土壤微生物丰富度没有显著影响,这可能是由于植被类型和数量的不同导致的,微生物群落丰富度受植被群落物种和数量的共同影响[47],因此退化高寒湿地植被群落结构对微生物功能多样性的影响有待进一步研究。
由主成分分析结果可看出,不同退化阶段的土壤微生物在对各类碳源的利用上出现明显差异,可见湿地退化改变了土壤微生物群落对碳源的代谢能力(图3)。季节性湖泊湿地退化过程中,土壤微生物对酯类碳源的相对利用率最高,酯类碳源为优势碳源,土壤微生物对单糖/糖苷/聚合物类碳源利用率的变化与土壤微生物活性的变化相似,均呈现先升高后下降趋势。糖类碳源极易被微生物同化,可以形成碳源比较丰富的环境[48],说明季节性湖泊湿地退化抑制了土壤中的糖类形成,不利于微生物生长繁殖,从而降低了土壤微生物活性及多样性。土壤微生物对酸类碳源的相对利用率随季节性湖泊湿地退化的加剧而增长,引起此变化的原因可能是在未退化阶段土壤中存在某些土壤微生物可以代谢产生酸类物质,其代谢产生的酸类物质可以供微生物利用[49],但随着季节性湖泊湿地退化该类微生物数量减少,因此土壤微生物对酸类碳源的相对利用率增加。本研究根据环境因子的共线性检测结果,在环境因子和土壤微生物利用各类碳源的能力的冗余分析中去除了土壤含水量、植被地上生物量和地下生物量等环境因子。但根据环境因子间相关性分析和土壤微生物对各类碳源的利用情况与环境因子之间的相关系分析结果,本研究推测土壤含水量可能是通过影响全氮含量、全磷含量、电导率和植被盖度等环境因子进而对土壤微生物碳源的利用产生影响。
环境因子与微生物碳源利用的冗余分析结果与土壤微生物AWCD,U,H′值与环境因子之间的相关性分析结果对比可知,土壤全氮含量和植被盖度对土壤微生物利用各类碳源的能力具有显著性影响(P<0.05),其中土壤微生物对单糖/糖苷/聚合物类碳源的相对利用率受各环境因子影响最大,并且影响土壤微生物对单糖/糖苷/聚合物类碳源的相对利用率的环境因子与影响土壤微生物活性的环境因子相同。而姚玉娇等人[50]通过对高寒草甸退化研究发现,其土壤微生物对碳源的利用主要受土壤有机质含量和电导率等影响。通过对AWCD,U,H′与土壤微生物利用不同碳源能力之间的相关性分析发现,在季节性湖泊湿地退化过程中,土壤微生物群落代谢功能多样性变化主要受土壤微生物对单糖/糖苷/聚合物类碳源和醇类利用率变化的影响。而刘燕等人[33]对退化草地的土壤微生物群落特征研究发现,随着退化加剧,微生物对不同碳源的利用产生差异,差异主要表现在对酸类、氨基酸类和酯类的利用。金裕华等人[51]通过对不同海拔的武夷山研究发现,武夷山不同海拔高度土壤微生物群落对各种碳源的利用度有所不同。因此,推断不同生态系统由于其生境的不同,导致其土壤微生物群系结构不同,从而使土壤微生物对各类碳源的利用能力产生差异,影响了微生物群落功能多样性。本研究推测各环境因子主要通过影响土壤微生物对单糖/糖苷/聚合物类碳源的利用,进而影响土壤微生物活性。本研究中选取的环境因子为基础的植被群落特征指标和土壤理化指标,因此,未来还需增加环境因子的类型,并结合高通量测序技术、宏基因组技术等多技术手段深入探讨季节性湖泊湿地退化过程中,土壤微生物对碳源的利用与土壤微生物群落结构的关系,从而进一步揭示季节性湖泊湿地退化过程中各环境因子对土壤微生物碳源利用的影响。
4 结论
在黄河源区季节性湖泊湿地退化过程中,土壤微生物对碳源的利用存在明显的空间分异,土壤微生物对酯类碳源的相对利用率始终最高,对酸类碳源的相对利用率呈增长趋势,对单糖/糖苷/聚合物类碳源的相对利用率变化与土壤微生物碳源代谢强度和功能多样性变化相似,均呈现先升高后下降趋势。不同退化阶段土壤微生物功能多样性的差异主要受微生物对单糖/糖苷/聚合物类和醇类碳源的相对利用率变化的影响。土壤全氮含量、含水量和植被盖度对微生物单糖/糖苷/聚合物类碳源的相对利用和土壤微生物活性均有显著影响。
