NaCl处理下全缘冬青和红果冬青根系的转录组活性比较
2022-11-30刘昌来余永帆杨甲定
张 强,周 鹏,刘昌来,余永帆,张 敏,杨甲定*
(1.南京林业大学,南方现代林业协同创新中心,南京林业大学生物与环境学院,江苏 南京 210037;2. 江苏省林业科学研究院,江苏 南京 211153)
土壤盐碱化是世界范围内最严重的环境问题之一,既造成巨大的农业、林业和园艺产业损失,也限制了野生植物种的分布[1]。全世界盐碱土面积约为9.54×104万hm2,而且还在继续增长[2];我国盐碱土总面积约3 600万 hm2,主要分布在东北、华北、西北内陆地区、黄淮海平原以及长江以北的滨海地带[3],其中以北方内陆的草甸盐土和海岸沿线的滨海盐土(滩涂)所占比例最大[4]。近年来随着地下水过度开采导致海水倒灌,沿海盐土面积呈现上升趋势。江苏省境内分布有大量的沿海滩涂,约占全国滩涂总面积的25%,被视为江苏省重要的后备土地资源[5]。但由于其土壤条件并不能直接用于农林作物的栽培,滩涂需要采用工程、物理、化学、农艺、生物等不同的措施进行改良[6],其中生物措施具有独特的优势,即提高植物耐盐性或培育耐盐新品种,使之能在滩涂正常栽培生长,通过吸收土壤的盐分改良滩涂土壤的物理化学性质,进而促进微生物和动物的定居而改善滩涂生态环境。因此耐盐植物的选育、栽培与推广正成为滩涂利用实践的重要内容。
由于不同区域滩涂的气候、环境等差异,通常需要检测具有较高应用潜力的乡土植物或外来引种植物的耐盐性,筛选出真正适合本地域滩涂生长的耐盐植物。1年生植物可通过比较植物分别在高盐和对照条件下的生物量积累来评估其耐盐性;但对木本植物,由于其相对较慢的生长速度,则可以利用一些简便的生理指标评估其耐盐性[7]。红果冬青(Ilexpurpurea)是在长江流域广泛分布的乡土树种,因其秋冬季节鲜艳的果实而被用于园林绿化观赏栽培,具有较强的抗风和耐湿潮土壤的能力,但不耐长期淹水[8];全缘冬青(I.integra)分布在我国东南浙江和福建的沿海地带,在日本和朝鲜半岛南部也有分布[9],鉴于长期生长在高盐贫瘠的滨海地带,推测全缘冬青应具有较高的耐盐性,因此被作为江苏滩涂栽培的备选引种植物之一。笔者前期选择全缘冬青和红果冬青作为测试对象,对其根系在外界NaCl处理下的生理变化进行了检测[10],发现全缘冬青根系在盐处理中积累的Na+相对较少、细胞膜脂过氧化程度相对较低,相比红果冬青,全缘冬青根系具有较高的耐盐性。但目前对两种冬青的根系在分子水平的耐盐机理却鲜见报道。本研究在前期生理检测的基础上,对NaCl处理下两种冬青根系的转录组进行了检测和生物信息学分析比较,以期从分子水平对全缘冬青根系高耐盐性的形成机理进行初步探究。
1 材料与方法
1.1 供试材料及处理
将生长于苗圃的红果冬青和全缘冬青的两年生扦插苗(株高约50~60 cm)从地里带根系挖出,用自来水洗去泥土后,移栽到1 L的塑料花盆(盆内基质为大田土壤和珍珠岩的等体积混合物,每盆1株植物)中。每种冬青各移栽75株,于南京林业大学白马教学科研基地(119°18′E, 31°06′N)玻璃温室内进行栽培,用1/2浓度的霍格兰德溶液从花盆基质表层对植株进行浇灌(每盆500 mL),每4天浇灌1次,共5次。 随后对植株进行NaCl盐胁迫处理,NaCl盐溶液为1/2浓度的霍格兰德溶液内含250 mmol/L NaCl,在实验开始时(0 h)取样,随即每个花盆浇灌500 mL的盐溶液3次(即0、24和48 h, 均在9:00—10:00完成),使花盆内基质NaCl盐溶液达饱和。
每种冬青在实验的0、1、6、24、72 h等时间点随机选择9株,随机分成3组作为3个生物学重复。将每个重复的植株根系挖出,快速洗去粘附的基质,剪取根系混合后再分成2部分,一部分冻存于液氮中带回实验室于-80 ℃保存,后用液氮冷冻研磨机磨成粉末作为鲜样;另一部分在105 ℃烘箱内烘烤3 h,然后在65 ℃下烘烤48 h后用普通电动磨样机磨成粉末作为干样。
1.2 总RNA的提取与检测
基于前期根系生理指标的检测与分析,选择两种冬青经0、6 (短期)和72 h(长期)盐处理等3个时间点,共18份根系样品提取总RNA。总RNA的提取使用北京天根生物技术有限公司的RNAprep Pure Kit RNA提取试剂盒抽提,操作过程按照试剂盒说明书进行。
1.3 文库构建与测序
RNA样品经质量检测合格后由美吉生物公司(http://www.majorbio.com/)基于Illumina Novaseq 6000 测序平台采用标准Illumina操作流程进行转录组测序。将获得的原始测序数据(raw data)用软件SeqPrep 和Sickle进行质控纯化,得到高质量的纯化数据(clean data);随后用Trinity软件(V.2013-02-25)[11]对每种冬青的所有纯化数据进行从头组装,将来自同一基因的不同同源转录本中最长的成员作为一个“unigene”。
通过与六大数据库(NR、Swiss-Prot、Pfam、COG、GO 和 KEGG)比对进行unigene的功能注释,即预测其功能或参与的分子途径。对转录因子预测通过与植物转录因子数据库PlantTFDB 4.0(http://planttfdb.cbi.pku.edu.cn/)比对完成。采用RSEM软件进行转录组的定量,获得基因的测序所得片断数(read counts),然后使用DESeq2差异分析软件进行盐处理样本与盐处理前样本间的基因差异表达分析,筛选出样本间的差异表达基因,即以0 h盐处理前的表达量为参照,筛选阈值为|log2(表达差异倍数)|≥1且校正后P值(Padjust) < 0.05,获得在盐处理6和72 h后根系内发生差异表达的基因。
1.4 基因差异表达分析及表达水平的荧光定量PCR检测
将两种冬青根系中的差异表达基因相对于各自0 h表达的log2(表达水平变化)值导入MeV(Multiple Experiment Viewer 4.9.0),以热图展示差异基因上调/下调表达的概况;利用美吉生物公司在线生信云平台(https://cloud.majorbio.com/),对每种冬青中的差异表达基因进行KEGG通路富集分析,默认当Padjust< 0.05时,认定此 KEGG 通路存在显著富集,并选择某些特定通路进一步分析其基因表达水平趋势,即以盐处理前0 h表达为参照,计算log2(表达水平变化),并根据基因注释信息分析其在特定代谢途径中的功能。
对每个用于RNA-Seq的RNA样品,取500 ng进行反转录合成cDNA(Takara反转录试剂盒,宝日医生物技术有限公司,北京),稀释10倍后用于荧光定量PCR检测[12]。选取本研究重点关注的与甜菜红素生物合成、亚油酸代谢、花生四烯酸代谢相关基因和过氧化氢酶基因共20个,根据各自的蛋白编码区序列,用Primer3在线软件(http://primer3.ut.ee)设计引物,引物长度为120~240 bp,引物序列见表1;qRT-PCR 检测使用艾科瑞公司 的 SYBR Green Premix Pro Taq HS试剂盒,在QuantStudio3荧光定量PCR仪(Appliedbiosystems,Thermo Fisher, 美国)中进行,反应条件为: 95 ℃ 30 s;95 ℃ 5 s,60 ℃ 30 s,共40 个循环;用 2-ΔΔCt法计算目标基因相对于内参基因的表达水平,最后计算其log2值与相对应的RNA-Seq表达水平(log2值)进行对比。

表1 荧光定量PCR 引物

表1(续)
2 结果与分析
2.1 RNA测序与差异表达基因的筛选
对每种冬青各9个RNA样品进行转录组测序获得的原始序列数据 (raw reads)经质检过滤,获得各自的纯化测序数据(clean reads);随后使用Trinity对全缘冬青的纯化测序数据进行从头组装,从全缘冬青中得到333 194个转录本(transcript)和239 429个unigene;从红果冬青中得到196 173个transcript和129 073个unigene。
鉴于unigene作为最长的转录本,通常能更好地代表该基因的编码信息,故笔者的分析以unigene为对象。如根系盐处理6和72 h后差异表达基因的数量(图1A)所示,全缘冬青盐处理6 h的差异表达基因数量为2 616,盐处理72 h为1 802;而红果冬青该指标在盐处理6 h为1 831,盐处理72 h为3 490。由于差异表达基因的筛选是以盐处理前为对照,从差异表达基因数量可以初步判断,全缘冬青根系对短期盐处理(6 h)响应迅速,差异表达基因数目较多,随着盐处理时间的延长,差异表达基因数目降低,转录组活性有向盐处理前回归的趋势;而红果冬青盐处理后期(72 h)差异表达基因数量多于盐处理初期,表明红果冬青根系随着盐处理时间的延长,其转录组活性趋向更偏离盐处理前的状态。同时从差异表达基因上调/下调热图概况(图1B)可知,盐处理6 h的全缘冬青根系中表达上调的差异基因远多于下调差异基因,而盐处理6 h的红果冬青根系中多数是表达下调的基因;和各自的盐处理6 h相比,盐处理72 h的全缘冬青和红果冬青根系中均表现为表达上调基因的数目下降而表达下调基因的数目上升。
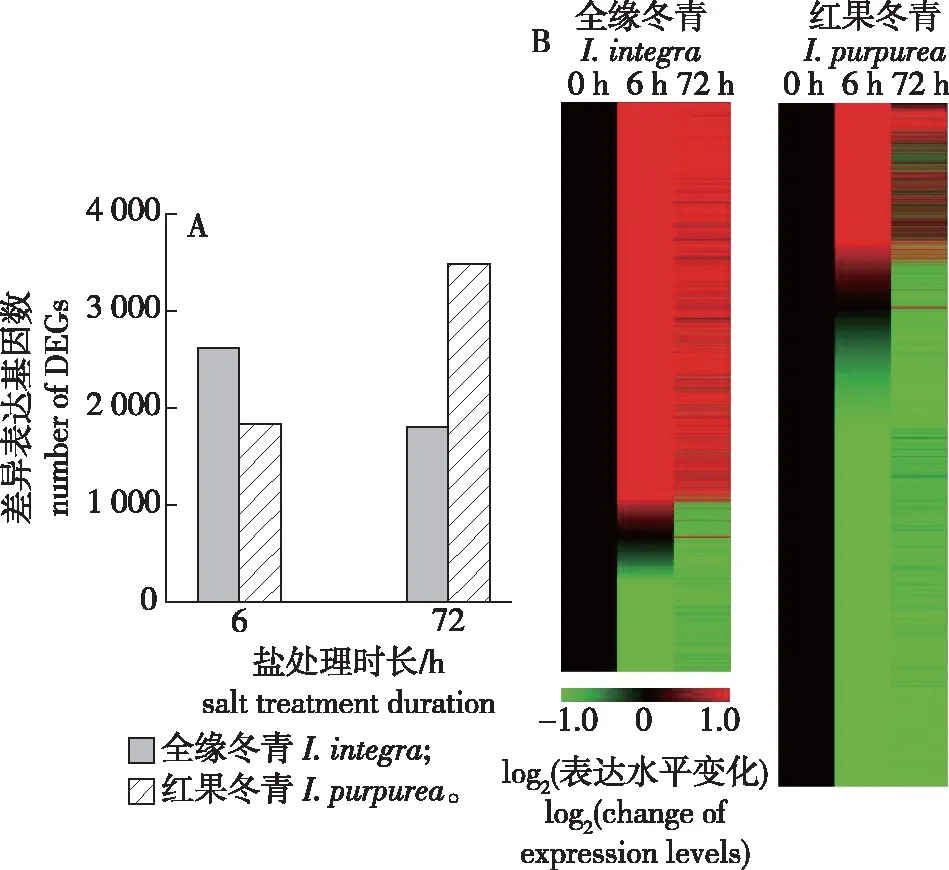
图1 全缘冬青和红果冬青根系盐处理6和72 h后差异表达基因的数量(A)和上调/下调热图(B)
2.2 两种冬青根系中差异表达基因的KEGG功能富集分析
如全缘冬青根系盐处理后的KEGG功能途径(图2A)所示,盐处理6 h全缘冬青根系的KEGG通路绝大部分基因属于代谢基因(203/223),包括碳(糖)代谢、氨基酸(酪氨酸)代谢、脂肪酸(亚油酸)代谢和次生代谢(苯丙烷、甜菜红素、类胡萝卜素生物合成)等,另有13个基因属于环境信息处理(植物激素信号转导)基因。全缘冬青根系盐处理72 h的KEGG富集通路包括细胞进程(内噬体,49/142)、环境信息处理(植物MAPK信号途径和植物激素信号转导,33/142)和代谢(29/142),代谢包括脂肪酸(亚油酸和花生四烯酸)代谢和甜菜红素、类胡萝卜素生物合成。6和72 h盐处理根系中共同的富集途径为植物激素信号转导、亚油酸代谢、类胡萝卜素生物合成和甜菜红素生物合成。
红果冬青根系盐处理6 h的KEGG通路(图2B)包括环境信息处理(植物激素信号转导和植物MAPK信号途径,49/99)、代谢(41/99)和植物昼夜节律(9/99),代谢则包括苯丙烷生物合成、光合作用、多种次生代谢物合成和类胡萝卜素生物合成)(图2B)。红果冬青根系盐处理72 h的KEGG富集通路包括细胞进程(内噬体,131/310)、环境信息处理(植物激素信号转导,31/310)和代谢(88/310),代谢包括苯丙烷生物合成、氮代谢、光合作用-天线蛋白、类胡萝卜素、双萜以及萜类和甾醇类生物合成。6和72 h盐处理共同的富集途径为植物激素信号转导、苯丙烷生物合成和类胡萝卜素生物合成。

图2 盐处理6和72 h后两种冬青根系中富集的KEGG功能途径
2.3 全缘冬青根系响应盐处理的特异代谢途径
比较两种冬青根系经6 h盐处理的富集途径(图2)发现,植物激素信号转导、苯丙烷生物合成、类胡萝卜素生物合成和植物昼夜节律为二者的共同途径;但全缘冬青有9种特异途径,包括酪氨酸代谢、甜菜红素生物合成、亚油酸代谢和六类碳代谢途径(即淀粉和蔗糖代谢、半乳糖代谢、糖酵解/糖异生、戊糖磷酸途径、戊糖与葡萄糖醛酸相互转化、乙醛酸和二碳酸代谢)等。
两种冬青根系盐处理72 h的共同途径有植物激素信号转导、内噬体、类胡萝卜素生物合成和肝癌等4种(图2),而全缘冬青特有的途径为植物MAPK信号途径、甜菜红素生物合成、亚油酸代谢和花生四烯酸代谢等4种。这些结果表明全缘冬青根系在6 h短期和72 h长期盐胁迫处理中,甜菜红素生物合成和脂肪酸(亚油酸、花生四烯酸)代谢均为特异的富集代谢通路,可能对全缘冬青根系的耐盐性形成具有特定的生理作用,将做进一步的分析。
2.4 全缘冬青中甜菜红素合成相关基因的表达
KEGG富集分析显示有9个基因可能参与甜菜红素生物合成,这些基因可以根据其在盐处理6和72 h后相对表达变化分为3类(图3):第1类4个基因在盐处理6 h时表达显著升高,至72 h保持平稳表达或表达降低[基因编号(下同)为TRINITY_DN294234_c0_g1、TRINITY_DN234089_c0_g1、TRINITY_DN509330_c0_g4和TRINITY_DN413471_c0_g1];第2类为盐处理6 h的表达水平相对于盐处理0 h无明显变化,盐处理72 h后表达显著降低(包括TRINITY_DN33926_c0_g1、TRINITY_DN62285_c0_g3和TRINITY_DN247553_c0_g1);第3类为盐处理6 h时表达显著降低,至72 h无显著变化(即TRINITY_DN350783_c0_g1、TRINITY_DN109022_c0_g1)。说明尽管这些甜菜红素合成相关的基因均注释为酪氨酸酶(tyrosinase)或包含酪氨酸酶中心功能域,但其表达水平对盐处理的响应存在上调/下调、早期/晚期的模式差异。
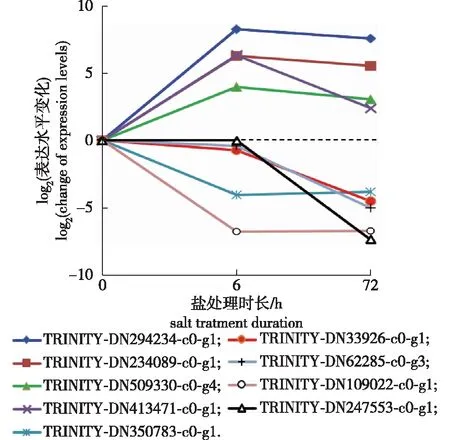
图3 全缘冬青根系甜菜红素生物合成9个相关基因表达水平在盐处理中的变化
2.5 全缘冬青中亚油酸代谢和花生四烯酸代谢相关基因的表达
KEGG功能富集分析显示全缘冬青根系在盐处理中有8个基因可能参与亚油酸代谢(图4),其中编号为TRINITY_DN173841_c0_g1、TRINITY_DN22314_c0_g1、TRINITY_DN75152_c0_g1和TRINITY_DN81998_c0_g1的4个基因(均注释为亚油酸酯10R-脂氧合酶类似物,linoleate 10R-lipoxygenase-like),TRINITY_DN173841_c0_g2(ψ-产氧酶A类似物,psi-producing oxygenase A-like)和TRINITY_DN383657_c0_g1(胞质磷脂酶A2γ,Cytosolic phospholipase A2 gamma)的表达水平在盐处理6 h后显著升高。盐处理72 h后,表达水平平稳保持或有所降低。另外编号为TRINITY_DN524668_c0_g2(细胞色素P450单加氧酶,Cytochrome P450 monooxygenase)和TRINITY_DN16496_c0_g1(三酰甘油酯酶,Triacylglycerol lipase)的2个基因则在盐处理过程中表达水平降低,其中TRINITY_DN524668_c0_g2仅在72 h盐处理后表达水平降低,而TRINITY_DN16496_c0_g1在盐处理6和72 h其表达水平持续降低。
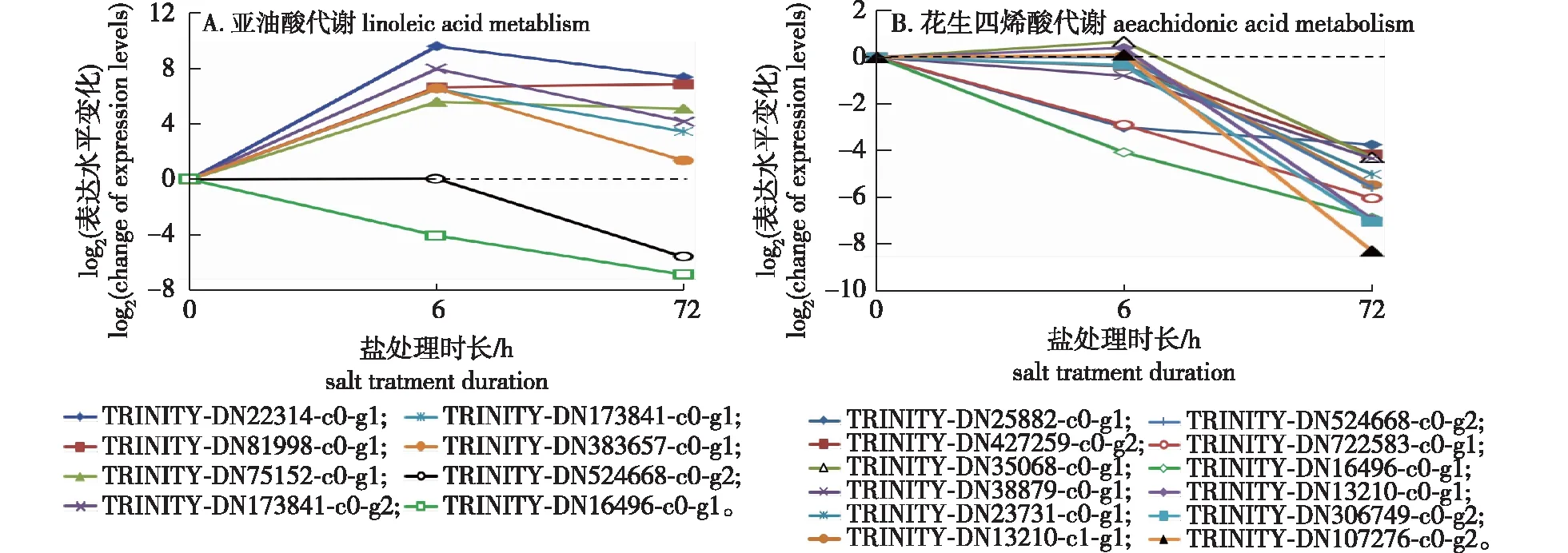
图4 全缘冬青根系中与亚油酸和花生四烯酸代谢相关基因在盐处理中的表达水平变化
如图4B所示,与亚油酸代谢相关基因不同,所有12个花生四烯酸代谢相关基因的表达水平没有显著升高的表现,其中编号为TRINITY_DN107276_c0_g2、TRINITY_DN13210_c0_g1、TRINITY_DN13210_c1_g1、TRINITY_DN23731_c0_g1、TRINITY_DN306749_c0_g2、TRINITY_DN38879_c0_g1、TRINITY_DN427259_c0_g2的7个基因(均注释为谷胱甘肽S-转移酶,glutathione S-transferase)、编号为TRINITY_DN35068_c0_g1(花生四烯酸15-脂氧合酶B,arachidonate 15-lipoxygenase B)和TRINITY_DN524668_c0_g2(细胞色素P450单加氧酶,Cytochrome P450 monooxygenase)的基因在盐处理6 h时表达水平无显著变化,但在盐处理72 h后显著降低;另有编号为TRINITY_DN25882_c0_g1(谷胱甘肽S-转移酶,glutathione S-transferase)、TRINITY_DN722583_c0_g1(谷胱甘肽S-转移酶,glutathione S-transferase)和TRINITY_DN16496_c0_g1(三酰甘油酯酶,Triacylglycerol lipase)的3个基因在盐处理6 h时表达水平即显著降低,盐处理72 h后其表达水平继续降低或保持平稳。
2.6 两种冬青根系中过氧化氢酶基因的比较
基于前期发现全缘冬青根系在盐处理中过氧化氢酶活性升高而红果冬青根系中过氧化氢酶活性降低的差异表现,笔者对两种冬青中可能编码过氧化氢酶的基因表达情况进行了分析。由于抗氧化酶相关的代谢并不在KEGG功能富集的通路中,用关键词搜索全缘冬青和红果冬青差异表达基因的功能注释,在全缘冬青发现7个注释为过氧化氢酶的差异表达基因,在红果冬青中找到3个注释为过氧化氢酶的差异表达基因(图5)。
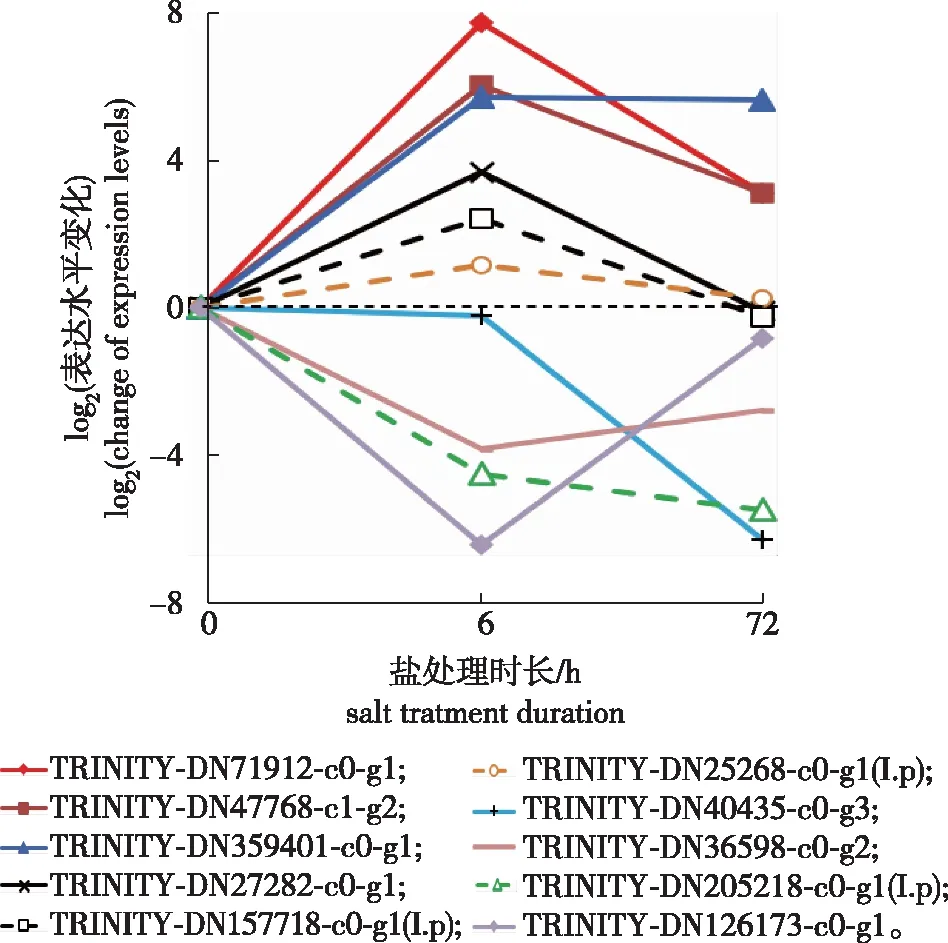
I.p表示红果冬青的 I. purpurea’s。下同。The same below.
全缘冬青7个过氧化氢酶基因中,有4个基因(编号为TRINITY_DN27282_c0_g1、TRINITY_DN359401_c0_g1、TRINITY_DN47768_c1_g2、和TRINITY_DN71912_c0_g1)在盐处理6 h时表达急剧升高,其表达水平分别是各自0 h时的12.9、53.4、66.2和216.4倍;盐处理72 h后,编号为TRINITY_DN27282_c0_g1的基因表达水平降低至0 h水平,TRINITY_DN359401_c0_g1表达水平依然是0 h水平的50倍,而TRINITY_DN71912_c0_g1和TRINITY_DN47768_c1_g2的表达水平相对其6 h表达水平有降低,但依然是盐处理前0 h水平的约9倍。其他2个基因(编号为TRINITY_DN36598_c0_g2和TRINITY_DN126173_c0_g1)在盐处理6 h时表达明显降低,72 h盐处理后表达水平恢复至0 h表达水平;另有TRINITY_DN40435_c0_g3盐处理6 h表达无变化,盐处理72 h,表达水平降低为其0 h水平的1.2%。
红果冬青中搜索到3个可能的过氧化氢酶基因,其中编号为TRINITY_DN157718_c0_g1和TRINITY_DN25268_c0_g1的基因表达水平在盐处理6 h时分别升高3倍,盐处理72 h后其表达降低至0 h水平;另有TRINITY_DN205218_c0_g1,其表达水平在盐处理中持续降低。这些结果说明全缘冬青和红果冬青根在盐胁迫处理时均有过氧化氢酶基因或上调或下调,但全缘冬青中受盐胁迫正调控的过氧化氢酶基因其表达水平在盐处理中远高于红果冬青,这与笔者前期生理测定中发现全缘冬青中过氧化氢酶活性在盐处理中迅速上升的变化趋势是吻合的[10]。
2.7 基因表达水平的荧光定量PCR验证
对上述可能与全缘冬青根系较高耐盐性相关的基因,其在盐处理中的表达水平进行了荧光定量PCR检测。这些随机选择的 20 个基因包括全缘冬青5个甜菜红素生物合成相关基因、5个亚油酸代谢相关基因、5个花生四烯酸代谢相关基因,还有3个全缘冬青过氧化氢酶基因和2个红果冬青过氧化氢酶基因。尽管荧光定量PCR检测的结果(图6)和转录组测序得到的结果(图3—图5)在具体升降倍数上并不完全相同,但在盐胁迫0、6和72 h的过程中,这些基因的表达变化趋势是一致的,由此证实了转录组测序结果的可靠性。

图6 基因表达水平的荧光定量PCR检测
3 讨 论
耐盐性是植物对外界高盐环境的综合响应,要成功提高某一目标植物的耐盐性,很大程度上依赖于首先对其关键耐盐机理的解析[13],进而通过操控关键代谢途径的活性达到提高植物耐盐性的目的。RNA测序技术(RNA-Seq)作为高通量手段,对不同生理/环境条件下植物器官、组织或细胞内所有转录本的表达情况进行检测分析;特别是对那些尚无参考基因组的物种,通过转录本的从头组装(Denovoassembly)可以全面理解其表型特征与转录组活性之间的关系,是目前深入研究植物转录组的有力工具之一[14]。由于没有近缘的植物基因组作为参考,本研究采用RNA-Seq测序技术对全缘冬青和红果冬青的根系在盐处理中的转录组活性进行了分析。
1)全缘冬青和红果冬青根系在盐处理过程中均有数千个基因表现出差异表达,但值得注意的是,全缘冬青根系盐处理6 h的差异基因数(2 616)多于盐处理72 h时的基因数(1 802);而红果冬青在盐处理6 h的差异基因数(1 831)少于盐处理72 h时的基因数(3 490),可能说明全缘冬青根系对短期盐处理(6 h)响应相对迅速,差异基因数较多,随着盐处理时间延长,其转录组活性有向对照回归的趋势;而红果冬青盐处理后期(72 h)差异基因数多于盐处理初期,表明红果冬青根系随着盐处理时间的延长,盐胁迫对其转录组活性的干扰越大(相对于盐处理前的转录组状态),这一结果暗示相对于红果冬青,全缘冬青根系可能具备一定程度上消除盐胁迫影响的能力。
2)植物对盐胁迫的响应和适应需要体内多种激素的交流和互作[15]。本研究中,植物激素信号转导是两种冬青根系在6 和72 h盐处理中均表现富集的通路,表明激素信号转导在两种冬青的盐胁迫响应中均发挥一定的作用;同时,类胡萝卜素生物合成也是两种冬青根系在6和72 h盐处理中共同的富集途径,暗示类胡萝卜素可能参与了两种冬青的盐胁迫响应。类胡萝卜素虽然在植物光合组织中主要作为光合色素的组分参与对光能的吸收[16],同时可作为抗氧化剂,清除活性氧,缓解植物组织的过氧化伤害[17]。番茄红素环化酶(lycopene cyclase,LCY)是类胡萝卜素生物合成途径中直接催化α-和β-胡萝卜素生成的关键酶,由α-和β-胡萝卜素生成类胡萝卜素(如芦丁、玉米黄质、环氧玉米黄质、紫黄质、新黄质等)[18],有研究表明,过表达盐角草(Salicorniaeuropaea)β-LCY基因可提高拟南芥叶片中类胡萝卜素的含量而减少了过氧化氢的形成,由此提高了拟南芥对200 mmol/L NaCl 的耐受性[19]。另外由于类胡萝卜素(9-cis-violaxanthin 和 9′-cis-neoxanthin)是植物胁迫激素脱落酸(ABA)的直接合成前体[20-21],而且某些类胡萝卜素合成突变体中生长素和ABA的生物合成均受到阻碍[22],暗示在两种冬青的盐胁迫响应中类胡萝卜素生物合成和植物激素信号转导可能存在某种关联,将来可对两种冬青根系在盐胁迫中的类胡萝卜素组分、含量、各类激素的合成/降解动态进行检测,解析是哪一类类胡萝卜素或激素在冬青盐胁迫响应中发挥重要作用。
甜菜红素生物合成和脂肪酸(亚油酸、花生四烯酸)代谢是全缘冬青根系在6 h(短期)和72 h(长期)盐胁迫处理中特异的富集代谢通路,在红果冬青中并不富集,因此推测可能与全缘冬青根系的耐盐性形成相关联。甜菜红素是一种水溶性含氮生物碱类色素,目前从17种植物中鉴定出了约75种甜菜红素结构[23],在盐地碱蓬(Suaedasalsa)中已证明甜菜红素既受高盐碱条件的诱导,也受黑暗、过氧化氢的诱导,是植物在逆境胁迫下产生的一种次生代谢物质,具有清除氧自由基的功能[24],甜菜红素是否参与全缘冬青根系的耐盐响应尚有待研究。
3)细胞膜作为植物细胞在胁迫条件下维持稳定胞内代谢环境的屏障,在盐碱胁迫中, 通常是细胞膜的结构和功能先受到伤害[25],在许多植物中已观察到盐胁迫会使细胞膜脂(磷脂、糖脂或鞘脂)含量、组分发生改变,特别是一些在正常条件下仅微量合成的特殊脂类会在胁迫条件下大量合成,由此改变膜的流动性和通透性,进而影响膜蛋白的特性和膜上信号分子的活性[26]。脂肪酸是膜脂分子的主要构成元件,膜脂脂肪酸的组分在植物各物种之间是保守的,通常软脂酸和亚油酸所占比例最大[27];饱和脂肪酸和不饱和脂肪酸的比率变化,因其能调控膜的流动性和通透性,被认为参与了植物对盐胁迫的响应[28]。本研究中,全缘冬青根系在盐处理中有8个基因可能参与亚油酸代谢,其中6个基因的表达水平在盐处理6 h后显著升高,随着盐处理72 h,表达水平平稳保持或有所降低,暗示亚油酸代谢与全缘冬青根系的盐胁迫响应相关联,这与花生(Arachishypogaea)研究相吻合,即经150 mmol/L NaCl胁迫7 d后的花生叶片中,单半乳糖甘油二酯(MGDG)、二半乳糖甘油二酯(DGDG)、硫代鼠李糖甘油二酯(SQDG)和磷脂酰甘油(PG)的双键指标(double bond index,DBI)升高,特别是软脂酸与亚油酸在这些膜脂中的含量有升高[29]。花生四烯酸是人体的一种必需脂肪酸,为5,8,11,14-二十碳四烯酸[30],拟南芥中的研究表明花生四烯酸作为一种信号分子,不仅可以激发脂肪酸介导的病原物防御反应,而且会引发普遍的胁迫信号网络[31],但目前尚无花生四烯酸与植物非生物胁迫关联的文献报道。本研究中全缘冬青根系所有12个花生四烯酸代谢相关基因在盐胁迫处理过程中总体表现为表达水平降低,这一趋势与全缘冬青耐盐性之间的相关性值得深入研究。
4)过氧化氢酶显著区别于其他过氧化代谢酶的特征是不需要还原剂的参与就能清除过氧化氢,而且对过氧化氢具有很高的专一性,对有机过氧化物的活性很低[32]。前期研究发现,尽管盐胁迫处理前(0 h),红果冬青根系中的过氧化氢酶活性显著高于全缘冬青;盐胁迫处理使过氧化氢酶活性在红果冬青中迅速降低,而在全缘冬青中迅速升高,全缘冬青过氧化氢酶活性在盐处理1~24 h均略高于红果冬青(无统计学差异),盐处理72 h则全缘冬青过氧化氢酶(CAT)活性显著高于红果冬青[10],说明全缘冬青根系在盐处理中具有更高的清除过氧化氢的能力。本研究中,全缘冬青中受盐胁迫正调控的过氧化氢酶基因数目多于红果冬青,并且其表达水平在盐处理中远高于红果冬青,说明盐处理中过氧化氢酶基因的转录上调是全缘冬青根系过氧化氢清除能力提高的重要原因,暗示将来可以尝试通过盐胁迫诱导表达过氧化氢酶基因来提高红果冬青的耐盐性。
综上所述,对全缘冬青和红果冬青盐处理根系的转录组测序为将来进一步分析二者耐盐性差异的生理学机理提供了有益的提示,植物激素信号转导、类胡萝卜素生物合成、甜菜红素生物合成和脂肪酸(脂类)代谢都是值得关注的生理过程。同时,功能鉴定全缘冬青根系中受盐处理高效诱导的过氧化氢酶基因,有可能用于将来通过遗传转化培育具有较高耐盐性的红果冬青或其他木本新品种。
