A chromosome minimalist view of genome regulation and evolution based on mouse chromosome engineering
2022-11-29qizhou
What is the minimum number of chromosomes that a eukaryotic species can tolerate? This question has long fascinated biologists as chromosomal rearrangements are associated with both speciation and various genetic diseases.Nature already has an answer, at least for mammals. In the 1970s, researchers at the Kunming Institute of Zoology (KIZ),Chinese Academy of Sciences (CAS), performed a comparative cytogenetic study of Indian muntjac (Muntiacus muntjak, deer species with 2n=6♀/2n=7♂ chromosomes),Chinese muntjac (M. reevesi, 2n=46 chromosomes), and their F1 hybrids to infer karyotypic evolution (Shi et al., 1980). The dramatic reduction in chromosome number from the ancestral state of 2n=70 in water deer (Hydropotes inermis) occurred in less than 10 million years following a series of Robertsonian(centromere-to-centromere, Rb) and tandem (centromere-totelomere, Td) fusions (Yin et al., 2021). How were these chromosomal fusions generated? What advantages were favored by natural selection? What were their functional impacts on the genome? As intriguing as they are, muntjacs are not a feasible experimental model for addressing these questions, whose answers will provide important insights into the principles of genome evolution and regulation at the chromosomal level.
The challenge of engineering chromosome numbers to a minimum was first conquered in yeast four years ago (Luo et al., 2018; Shao et al., 2018). Most recently, two groups of researchers at CAS independently achieved artificial chromosome fusions in mice (Wang et al., 2022; Zhang et al.,2022; Figure 1). The technical hurdles are obviously immense:in yeast, simultaneous deletions of centromeres and telomeres of any two of the total 16 chromosomes followed by their linkage through recombination are required, resulting in as few asn=1 or 2 (Luo et al., 2018; Shao et al., 2018), or even one circular chromosome (Shao et al., 2019). In mice,the sophisticated manipulation of yeast-like haploid embryonic stem cells (haESCs) may be accompanied by unexpected accidents. For example, in Wang et al. (2022), after the telomere of chr1 (the largest chromosome) was engineered to tandemly fuse with the centromere of chr2 (chr1+2), the fused chromosome broke into a partial chr1, which, in turn, fused with chr2. This accident lead to valuable discoveries.
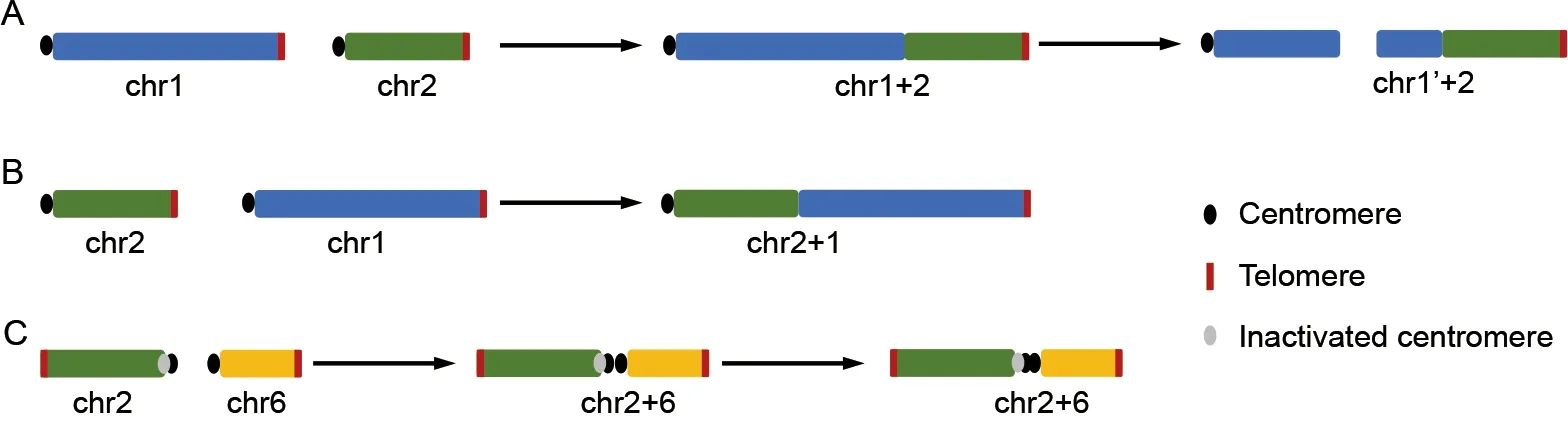
Figure 1 Engineered chromosome fusions in mice
A striking common outcome of minimalist trials to reduce chromosome numbers in yeast and mice, despite their great differences, is that chromosome fusions have little impact on global gene expression and result in relatively mild phenotypic changes. As few as 0.5% of yeast genes and less than 10% of mouse genes show significant differences in gene expression in response to chromosome fusions. This must be interpreted under the context that immediately after fusion, dramatic alterations in the three-dimensional (3D) genome architecture,e.g., topologically associated domains (TAD) and novel interchromosomal contacts, have already occurred compared to unfused chromosomes. These counterintuitive results have several important implications: first, chromosome fusions may occur frequently and reach fixation as slightly/mildly deleterious mutations during evolution, particularly when deleterious consequences can be counteracted or overwhelmed. For example, in humans and chickens (Daniel,2002; Dinkel, 1975), Rb-fused chromosomes can be preferentially transmitted during asymmetric female meiosis(the “centromeric drive hypothesis” (Henikoff & Malik, 2002;Malik & Bayes, 2006)). This is also related to the considerable karyotypic diversity of muntjac deer, in which chromosome numbers are mainly shaped by fusion (Yin et al., 2021).Second, 3D genome architecture appears to play a more structural than a regulatory role in the nucleus. TADs are hypothesized to regulate gene expression by restricting specific interactions betweencis-regulatory elements and their target genes. However, complex chromosomal rearrangements (e.g., inversions and segmental duplications)that disrupt TADs inDrosophilado not result in corresponding changes in gene expression (Ghavi-Helm et al., 2019). These weak transcriptomic responses to chromosome fusions challenge the importance of TADs in regulating gene expression. Lastly, despite minor expression changes,crosses between yeast strains with fused or unfused chromosomes result in fewer offspring (Luo et al., 2018).Furthermore, down-regulation of theCapn11gene is associated with higher levels of anxiety in mice with the chr1+2 Td-fusion karyotype compared to wild-type mice(Wang et al., 2022). These phenotypic results recapitulate in the laboratory how reproductive isolation or speciation may occur in nature due to cellular and behavioral factors.
As mentioned above, chromosome engineering studies have been achieved in the laboratory for the first time (Wang et al., 2022; Zhang et al., 2022), yielding several unexpected and noteworthy findings that raise new questions for future studies. First, selective constraints from nuclear space seem to act on certain combinations of chromosome fusions, which may explain the secondary breakage of chr1+2 in mice (Wang et al., 2022), and the reduction in yeast chromosome number ton=2 (Luo et al., 2018), but not further. Although different combinations and orientations of fusions do not appear to have a large effect on gene expression, they may impact how fused chromosomes are folded and fitted into the nucleus or how they behavior during cell division. For example, chr2+1 mice show a higher tendency toward anaphase lag and polyploidization than chr1+2 in mice (Wang et al., 2022). This may result in a secondary breakage, similar to the case of chr1+2, manifesting as a translocation between two different chromosomes (Wang et al., 2022). This process may have played an important role during the evolution of mammalian chromosomes, which show many more translocations than other amniotes. However, future in-depth studies of these maladapted fused chromosomes are required to test this hypothesis. Second, mouse models with engineered chromosome fusions provide an excellent experimental system for understanding centromere biology. Despite their essential role in cell division, little is known about the formation and evolution of eukaryotic centromeres due to their rapidly evolving and complex sequence features. Surprisingly,centromeres of the classic model organismD. melanogasterwere only recently characterized at the sequence level (Chang et al., 2019). Interestingly, Zhang et al. (2022) found that Chr2 has one inactivated and one activated centromere. The inactivated centromere can only bind to one, not both, of the functional centromeric proteins. This may reflect the first critical step for the fixation of an Rb fusion during evolution,before the subsequent loss of the centromeric repeat sequence observed in much older Rb fusions, e.g., in muntjacs (Yin et al., 2021). In their study, Zhang et al. (2022)targeted the centromeric satellite sequences for excision,providing experimental evidence that such deletions can produce Rb chromosome fusions.
In short, the mouse chromosome engineering work by Wang et al. (2022) and Zhang et al. (2022) provides a platform for future in-depth study of the evolutionary mechanisms and diseases associated with chromosome fusions.
COMPETING INTERESTS
The author declares that he has no competing interests.
AUTHORS’ CONTRIBUTIONS
Q.Z. wrote the draft and revised, read, and approved the final version of the manuscript.
杂志排行

Zoological Research的其它文章
- Optimization of sgRNA expression strategy to generate multiplex gene-edited pigs
- Virome in healthy pangolins reveals compatibility with multiple potentially zoonotic viruses
- Chronic lithium treatment ameliorates ketamineinduced mania-like behavior via the PI3K-AKT signaling pathway
- A glimpse into the biodiversity of insects in Yunnan: An updated and annotated checklist of butterflies(Lepidoptera, Papilionoidea)
- Unveiling the functional and evolutionary landscape of RNA editing in chicken using genomics and transcriptomics
- Animal models of Alzheimer’s disease: Applications,evaluation, and perspectives