What does it mean to be a macronutritional generalist?A five-year case study in wild rhesus macaques(Macaca mulatta)
2022-11-29zhen-weicui,yingzhang,jia-baoyan等
DEAR EDITOR,
Dietary generalism is widely associated with ecological flexibility, but the mechanisms of this relationship remain poorly understood. A recently developed body of theory on the relationship between generalism/specialism at the levels of foods selected vs. nutrients ingested, termed multi-level dietary niche theory, offers a powerful opportunity to deepen our understanding of the dietary generalist-specialist spectrum and its ecological significance. We applied the multi-level dietary niche framework in an intensive and extensive study of food selection and nutrient regulation responses to natural resource variation in a wild population of rhesus macaque(Macaca mulatta). We used full-day individual animal follows over 17 successive seasons to examine the patterns of resource (food and nutrient) availability and responses of macaques at the level of food selection and macronutrient intake. In addition, we analyzed the seasonal availability of their keystone resource, i.e., acorns rich in fat and carbohydrates, over a longer term, spanning 10 successive years. Results showed that even though macronutrient intakes resembled a pattern previously associated with species that eat a wide range of foods (“food generalists”), in the population we studies it is associated with the consumption of few food types (i.e., they are “food specialists”) that varied widely in macronutrient composition and fluctuated predictably in relative availability across seasons and alternate years. We integrated these results with energy budgets and literature data to build a verbal model of how a generalist pattern of macronutrient regulation interfaces with nutritional physiology and resource variability to buffer a primate population that is reliant on a few food types from broad climatic fluctuations.
Understanding what it means to be a dietary generalist vs.specialist is important because the variety of diets an animal can subsist on has a strong bearing on its capacity to tolerate ecological variation, including global change (Monaco et al.,2020). Multi-level dietary niche theory offers new opportunities for understanding dietary generalism vs. specialism through recognizing that the capacity of an animal to exploit a diversity of foods (a “food generalist”) is different from the capacity to exploit a diversity of nutritional mixtures (a “nutrient generalist”) and enabling the modelling of these relationships using nutritional geometry (Machovsky-Capuska et al., 2016).Rhesus macaques (M. mulatta) provide an excellent model system for applying multi-level dietary niche theory to a species in the wild. As a primate that is geographically widespread and important in biomedical research, rhesus macaques have been the subject of more research than any other primate, including research examining the relationships between food selection and macronutrition in the wild. Rhesus macaques are also the most ecologically generalist nonhuman primate, occupying a range spanning tropical to temperate habitats. Further, rhesus macaques are the only vertebrate known to date to exhibit the pattern of macronutrient regulation that has been associated in theoretical models and laboratory comparisons of generalist vs. specialist insects with ecological generalism (Cui et al.,2018). Based on its parameters in nutritional geometry models, this pattern is termed “Equal Distance” regulation (Cui et al., 2018).
In this study, we applied the multi-level dietary niche framework to examine the ecological basis of dietary generalism in the temperate sub-species of rhesus macaque that lives at the edge of the species range, i.e., Taihangshan population (M. mulatta tcheliensis). We analyzed 10 years of data on the availability of the keystone resource, fat- and carbohydrate-rich acorns, together with full-day individual animal follows over 17 successive seasons to examine the relationships between macronutrient selection, food selection,and seasonal and interannual patterns of resource variation.We integrated the ecological and behavioural results with energy budgets calculated using seasonal body weight changes and energy expenditure equations for primates to build a model of how generalist patterns of macronutrient regulation interface with nutritional physiology and resource variability to buffer rhesus macaques from resource fluctuations. Further details on study site, group, and specific methods are available in the Supplementary Materials.
Our results showed that the production of acorns, which are the principal source of lipids and carbohydrates for Taihangshan macaques and are thus a critically important resource for this temperate population (Figure 1A), varied widely across years and seasons. Between 2007 and 2016,acorn production, which occurs in early autumn and determines lipid and carbohydrate availability for that autumn and winter and following spring, showed a two-year mast cycle(Figure 1B), as has previously been documented in other species ofQuercus(Veldhorst et al., 2009). This cycle was statistically significant, with the current acorn crop negatively correlated with crops 1, 3, 5, 7, and 9 years prior (r=-0.757,-0.680, -0.617, -0.398, -0.105, and -0.067, respectively,P<0.05) and positively correlated with crops 2, 4, 6, and 8 years prior (r=0.680, 0.472, 0.211, and 0.079, respectively,P<0.05). The biennial cycle did not affect acorn consumption in autumn (GLMM:F1,86=1.149,P=0.287), when acorn consumption was consistently high, or in summer, when acorns were not eaten because by that time the crop was invariably depleted. It did, however, affect consumption in winter (F1,1.991=19.666,P=0.048) and especially spring(F1,3.028=39.694,P=0.008) (Figure 1C). During these seasons,in non-mast years, acorns were largely or entirely replaced in the diet by other foods (Figure 1D), principally bark and twigs in winter and leaves in spring (Figure 1C).
The biennial cycle in acorn availability had a significant impact on dietary macronutrient balance (Figure 1E). The ratio of dietary energy derived from protein to non-protein energy(lipid and carbohydrate), henceforth termed the protein to nonprotein ratio, was consistently low in autumn (when acorn consumption was high) and consistently high in summer when acorns were absent from the diet. In winter and spring,however, the protein to non-protein ratio varied with annual variation in acorn availability, being low in mast years and high in non-mast years. Despite this interannual variation in the protein to non-protein ratio , total energy intake (protein + nonprotein energy) did not differ between mast and non-mast years (Figure 1E), demonstrating that the macaques compensated for the ecologically-enforced reduction in energy eaten as lipids and carbohydrates by a counter-balancing increase in energy eaten as protein. In spring, the season in which reproductive females lactate, the effect of non-mast years on dietary macronutrient ratios was especially high(12.69% protein in 2013 to 30.12% protein in 2014, GLMM:F4,126=707.101,P<0.0001), but did not differ between lactating and non-lactating monkeys (GLMM:F1,126=0.061,P=0.805).As with non-lactating monkeys, total energy intake in lactating monkeys did not vary with annual variation in the protein to non-protein ratio (Kruskal-Wallis test:chi-squared=9.345,df=4, 40,P=0.053). Total energy intake, however, was significantly higher during lactation (414 kJ/kg/day vs. 321 kJ/kg/day, GLMM:F4,131=215.527,P<0.0001, Figure 1E),equivalent to autumn and winter intake in non-lactating adults.This pattern of macronutrient intake, in which total energy intake (protein + fat + carbohydrate energy) remains constant despite large variations in the dietary macronutrient ratio, is the Equal Distance pattern referred to above.
Our energy budget calculations (Figure 1F) showed that the sum of non-protein energy ingested and energy liberated from lost body fat was less than energy expenditure in summer(GLMM:F1,160=399.800,P<0.001) and spring of non-mast years (GLMM:F1,66=81.304,P<0.001). This suggests that dietary protein was required to balance the energy budget in these periods. In contrast, in the spring of mast years, energy ingested in the form of lipids and carbohydrates was sufficient to meet energy expenditure (GLMM:F1,99.866=0.555,P=0.458).However, the intake of lipids and carbohydrates was not sufficient to account for the observed gain in body fat,suggesting that even in mast years dietary protein is used as an energy source by the Taihangshan macaques. Cui et al.(2018) was the first study to previously document the Equal Distance pattern of macronutrient regulation in a species of primate, in that case using data from only three successive springs. By extending the analysis to 17 successive seasons,results of the present study confirm the existence of the Equal Distance pattern in Taihangshan macaques and, most importantly, place it in the broader context of seasonal and inter-annual variation in resource availability and macronutrient and energy budgets of the monkeys.
As discussed by Cui et al. (2018), Equal Distance pattern(Figure 1E), has previously been observed only in generalist insect herbivores, even though many studies have used nutritional geometry to analyze patterns of macronutrient regulation in other primate species. In all other primates studied, either protein or non-protein energy is maintained more constant than the other and consequently total energy intake varies with ecologically imposed variation in dietary protein to non-protein ratio. This aligns with the theoretically derived expectation that generalists should be selected for metabolic flexibility, enabling them to switch metabolic fuels depending on mix of macronutrients in the diet. A key physiological component of metabolic flexibility is gluconeogenesis, in which amino acids are deaminated in the liver to generate glucose for energy metabolism and nitrogenous excretory products (Watford, 2015). In primates,including humans, gluconeogenesis typically occurs at significant levels only during prolonged fasting and starvation(Watford, 2015). We are unaware of any physiological studies directly testing for abnormally high rates of gluconeogenesis in rhesus macaques, but a recent population genomics study found that several genes associated with gluconeogenesis are under positive selection in the Taihangshan population (Liu et al., 2018).
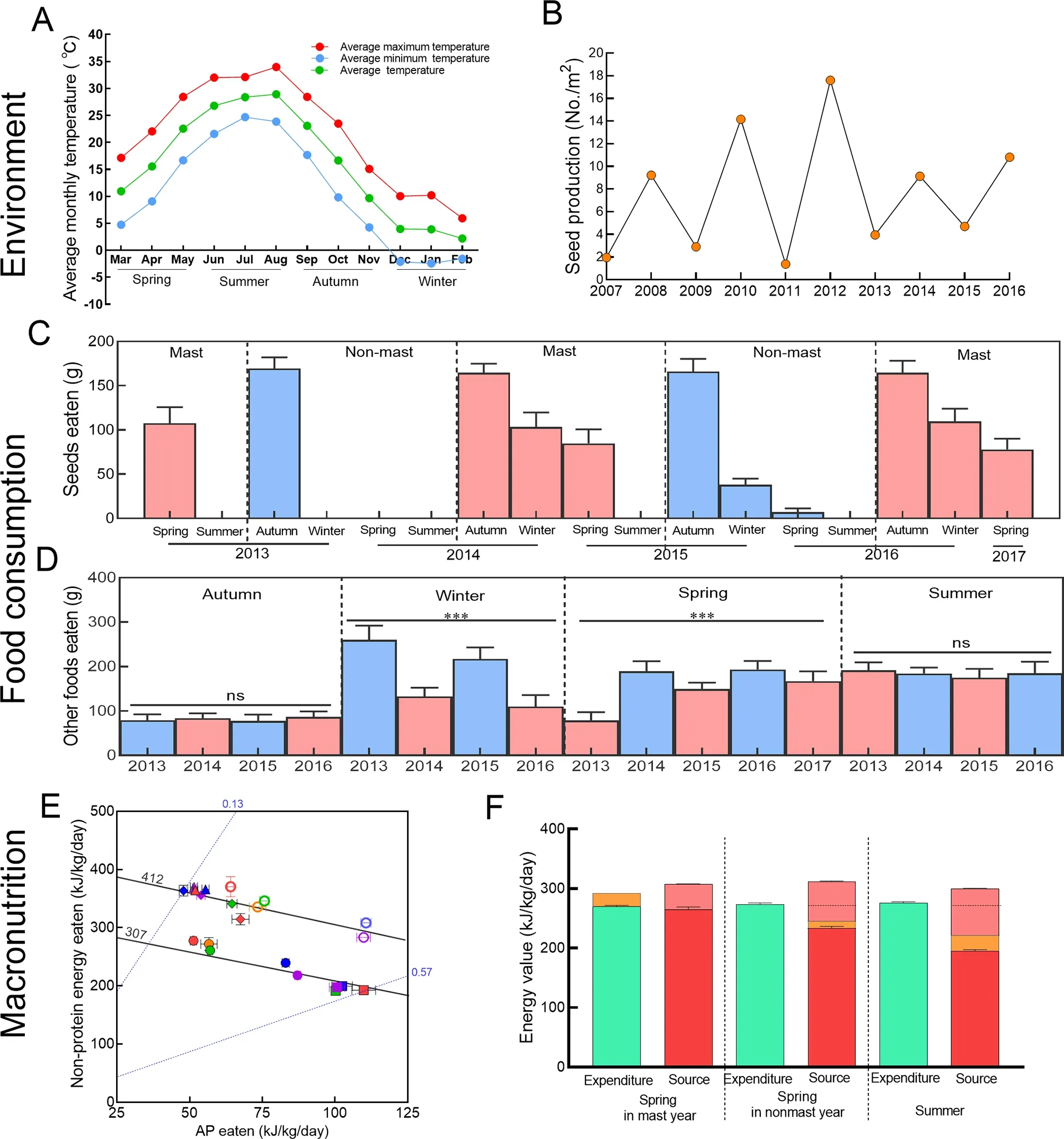
Figure 1 Relationships among environmental (A, B), dietary (C, D), and nutritional (E, F) dimensions of variability in the nutritional ecology of Taihangshan rhesus macaques
These results raise the question of which ecological circumstances selected for the Equal Distance pattern of macronutrient regulation in Taihangshan macaques. In the context of insects, it has been hypothesized that animals that feed on a large number of foods (termed “food generalists” by Machovsky-Capuska et al. (2016)) would be selected to ingest nutritionally imbalanced macronutrient mixtures because their broad diet increases the probability that they can access complementary foods that enable them to rebalance their diet(Raubenheimer & Simpson, 2003). However, this alone cannot explain Equal Distance regulation in the Taihangshan macaques. Although rhesus macaques as a species eat many foods, our study population was observed to eat only 57 different foods in eight food categories from 33 species over 5 years of detailed study (Cui et al., 2019), which is lower than that found in several other primates without Equal Distance regulation (Supplementary Table S1). On the other hand, our study population utilized foods with protein to non-protein ratio ranging from 0.12 to 0.58, a broad range compared to several species that do not show Equal Distance (Supplementary Table S2).
A factor very likely associated with the evolution of different macronutrient regulatory strategies, but which hitherto has remained unexplored in this context, is the temporal pattern of resource variability. The relative simplicity of our temperate study system enabled us to examine this in detail by measuring patterns of availability and exploitation of resources across seasons and over several consecutive years. Our results showed a complex yet vivid pattern comprising three important components of variability. First, there was substantial seasonal variation in availability of the keystone resource for this population i.e., fat- and carbohydrate-rich acorns from consistently high in autumn to absent in summer.Second, seasonal variation was superimposed on a two-year masting cycle alternating between large and small crops(Figure 1B). Third, there were differences in acorn production during non-mast years (Figure 1B), and hence in the rate of acorn depletion (Figure 1C). Across the seasons in our study in which acorns were eaten (i.e., autumn, winter and spring),acorn consumption ranged from low in winter and spring(2015-2016), to depletion in autumn resulting in no observed acorn consumption in winter and spring (2013-2014).
Our results suggest a model according to which temporal patterns of resource availability interact with the low number of different foods available and large disparities in dietary macronutrient ratios to favor Equal Distance regulation in Taihangshan macaques. In this model, annual confinement for at least the three months of summer to foods with high protein has favored physiological adaptations to substitute excess ingested protein for limited non-protein energy (fat and carbohydrate) to fuel energy metabolism. This increased metabolic flexibility not only enables the monkeys to meet their energy needs during summer, but also serves as a fallback strategy to attain energy equilibrium when confined to a high protein diet during winter and spring of non-mast years when fat- and carbohydrate-rich acorns are depleted (Figure 1F).Our study population was observed to subsist on a diet of approximately 30% of protein for nine consecutive months(winter 2013 to autumn 2014) - a level of protein close to the upper limit for humans and equivalent to the diets of some carnivores, such as domestic dogs. The data presented in Figure 1B suggest this may not be an uncommon occurrence.Our model is consistent with the observation that other taxa associated with high protein diets likewise have macronutrient regulatory strategies in which protein is over-ingested in diets with protein to non-protein ratios that exceed the target diet,including predators and mountain gorillas, which are annually confined for 8 months to a low-fruit diet comprised of 30%protein (Simpson & Raubenheimer, 2012).
Niche breadth is considered an important predictor of extinction risk, with dietary specialists being more vulnerable than generalists (Machovsky-Capuska et al., 2016). Our application of multi-level niche theory to rhesus macaques demonstrates that the concept of generalism vs. specialism is complex, and dependent not only on the number of foods exploited, but also the compositional disparity among those foods and their temporal patterns of availability. A priority is to apply these concepts more broadly to understand how different dimensions of niche breadth relate to extinction risk.
SCIENTIFIC FIELD SURVEY PERMISSION INFORMATION
Permission for field surveys in the Wangwu Mountain area was granted by the Taihangshan Macaque National Nature Reserve in Jiyuan, Henan Province, China. This research complied with the protocols approved by the appropriate wildlife conservation committees of China and adhered to the legal requirements of China.
SUPPLEMENTARY DATA
Supplementary data to this article can be found online.
COMPETING INTERESTS
The authors declare that they have no competing interests.
AUTHOR CONTRIBUTIONS
Z.W.C., D.R., and J.Q.L. conceived the ideas and designed the methodology; Z.W.C., Q.S., and Y.Z. collected the data;Z.W.C. and Y.F.Z. performed the laboratory protocols; Z.W.C.,D.R., J.B.Y., Y.L.D., C.R.R., and H.J.S. analyzed the data;Z.W.C. and D.R. led the writing of the manuscript. Z.W.C. and D.R. revised the manuscript. All authors read and approved the final version of the manuscript.
ACKNOWLEDGMENTS
We thank the Jiyuan Administration of Taihangshan Macaque National Nature Reserve for permission to conduct field work and logistic support throughout this study. We are grateful to Mr. Guo-Liang Zhao for assistance with field work. We thank Prof. Shi-Xin Zhu (School of Life Science, Zhengzhou University) for plant species identification and Prof. Zhen-Long Wang for helpful suggestions on the study. We also thank Prof. Bao-Guo Li, Prof. Song-Tao Guo, and Dr. Rong Hou for their assistance with nutritional analyses.
杂志排行

Zoological Research的其它文章
- Optimization of sgRNA expression strategy to generate multiplex gene-edited pigs
- Virome in healthy pangolins reveals compatibility with multiple potentially zoonotic viruses
- Chronic lithium treatment ameliorates ketamineinduced mania-like behavior via the PI3K-AKT signaling pathway
- A glimpse into the biodiversity of insects in Yunnan: An updated and annotated checklist of butterflies(Lepidoptera, Papilionoidea)
- Unveiling the functional and evolutionary landscape of RNA editing in chicken using genomics and transcriptomics
- Animal models of Alzheimer’s disease: Applications,evaluation, and perspectives