浙江省地方菜豆种质资源鉴定与遗传多样性分析
2022-11-29汪宝根董君暘李素娟鲁忠富吴晓花李国景吴新义
汪宝根,董君暘,汪 颖,李素娟,王 尖,鲁忠富,吴晓花,李国景,吴新义,,*
(1.浙江省农业科学院 蔬菜研究所,农业农村部南方豆类蔬菜种质创新与分子育种重点实验室(部省共建),浙江 杭州 310021; 2.浙江农林大学 园艺科学学院,浙江 杭州 311300; 3.浙江省农业科学院 省部共建农产品质量安全危害因子与风险防控国家重点实验室,浙江 杭州 310021)
菜豆(PhaseolusvulgarisL.)(2n=2x=22)隶属豆科(Fabaceae)菜豆族(Trib.PhasoleaeDC.)菜豆属(PhaseolusL.),是世界范围内最重要的豆类作物之一。野生菜豆起源于美洲,经长期驯化和地理隔离后,形成了安第斯和中美洲2个多样性中心。栽培菜豆主要包括2种类型,即以干籽粒做粮食或饲料的粮用型菜豆(dry bean)和以嫩荚做菜用的菜用型菜豆(snap bean)。菜豆约在15世纪传入我国,别名四季豆、刀豆、春分豆、梅豆、芸豆、玉豆等,既是我国重要的食用豆类,也是我国广泛栽培的重要豆类蔬菜[1]。据联合国粮农组织2020年统计,我国粮用型菜豆的种植面积约为74.21万hm2,产量约占全球粮用型菜豆总产量的五分之一;菜用型菜豆的种植面积约为66.61万hm2,产量位居全球第一。
我国菜豆种质资源丰富,国家种质资源库目前收集保存菜豆资源超过4 900份,大部分为地方品种[2]。这些资源中仅有部分材料调查了基本农艺性状,对其遗传多样性、亲缘关系研究较少。张赤红等[1]利用36对SSR引物对332份国内普通菜豆、16份国外普通菜豆和29份野生菜豆的遗传多样性进行了分析,发现国内普通菜豆的遗传多样性大于国外普通菜豆和野生菜豆,其中贵州、云南、黑龙江等省普通菜豆的遗传多样性较为丰富;Zhang等[2]使用30个SSR标记研究了229份中国菜豆地方品种的遗传多样性,并将之分为安第斯基因池和中美洲基因池两大类,每一类又含有2个亚类;Wu等[3]对683份菜豆种质进行全基因组重测序,构建了包含480万个SNP的高密度、高精度的单倍型图谱,主成分分析和系统进化树分析均支持将该群体分为安第斯基因池和中美洲基因池两大类,进一步可以将之分为5个亚群;彭丽娟等[4]利用7对SSR和13对ISSR标记分析了70份菜豆品种的遗传多样性,系统进化树分析将之分为4个类群7个亚类。
开展地方种质资源精准鉴定与遗传多样性分析有利于梳理现有育种资源,挖掘优异种质,为适合区域性消费需求的品种选育提供第一手资料。菜豆为自花授粉二倍体,基因组大小约600 Mb[5]。随着菜豆基因组的释放和基因组测序技术的快速发展,单核苷酸多态性标记(single nucleotide polymorphism,SNP)由于其数量多、多态性丰富逐步成为菜豆遗传研究的主流标记,如Song等[6]对17份菜豆种质测序后鉴定出992 682个SNP,据此开发了含5 398个SNP的高质量芯片BARCBean6K_3 BeadChip;Wu等[3]通过对683份菜豆种质重测序,鉴定出4 811 097个高质量SNP,尽管这些海量SNP信息为菜豆遗传和育种研究提供了强有力的工具,但在使用SNP标记进行基因型鉴定时,由于芯片造价较高、锚定的SNP无法灵活替换,而重测序技术不仅费用高,在不同材料间对SNP鉴定的重复性也较差,因而在普通育种单位中应用受限。竞争性等位变异特异性PCR(kompetitive allele specific PCR,KASP)技术由于在SNP数目和群体大小上可灵活选择的优势,逐步成为利用SNP标记开展个性化研究的理想方法,如Hurtado-Gonzales等[7]将BARCBean6K_3 BeadChip芯片上的SNP转化为KASP标记用于菜豆抗锈病基因的精细定位。此外,KASP标记在豇豆、瓠瓜等作物上也已经广泛用于遗传多样性分析[8-9]。
菜豆是浙江省优势特色蔬菜,在临安、磐安等地已经形成规模化种植基地,是夏季高山蔬菜的主导产品。浙江省地理形态丰富,自然条件优越,有利于生物多样性的形成和保护。目前在浙江省搜集发现的菜豆地方品种有44份,但这些资源尚未进行系统的鉴定与遗传多样性分析。因此,本研究拟对已经收集到的菜豆种质进行详细的表型精准鉴定,并开发KASP标记鉴定其基因型,评价群体遗传多样性,为地方特色种质的鉴定和有效利用提供参考。
1 材料与方法
1.1 试验材料
供试材料含有44份浙江菜豆地方种质,其中28份为“第三次全国农作物种质资源普查与收集行动”所收集,其余16份为本课题组历年来搜集得到,具体编号、名称和基本信息见表1。
1.2 农艺性状调查
试验于2018—2019年连续2 a在浙江省农业科学院海宁杨渡科研创新基地进行,采用完全随机区组设计,2次重复,每个材料每个重复种植2列,搭人字架引导植株攀爬,畦宽连沟1.5 m,株距40 cm,行距75 cm,每份材料种植20穴,田间肥水管理和生产上保持一致。
参照《菜豆种质资源描述规范和数据标准》[10],每份材料随机选取10株进行农艺性状调查,对株型、小叶叶型、叶色、始花节位、嫩荚长、嫩荚宽、荚型、嫩荚横切面、嫩荚主色、单荚重、种子形状、种皮色、千粒重共13个重要性状进行调查。取2次重复的平均值作为年度鉴定结果,把2年鉴定结果的平均值作为最终鉴定结果。调查嫩荚性状时每份材料至少取10个处于商品成熟期的鲜荚,利用尺子精确测量荚长、荚宽,称重后计算单荚重。分别对幼苗、花、商品嫩荚、叶片、植株茎节、种子等进行拍照。
1.3 KASP标记开发
根据Song等[6]报道的BARCBean6K_3 BeadChip芯片,按照SNP在基因组上的位置均匀选择185个SNP转化为KASP标记。KASP引物设计采用LGC公司的KrakenTM软件,根据目的SNP位点上下游DNA序列,每个位点设计3条引物,包括一条反向通用引物和2条正向特异引物,2条正向特异引物分别添加FAM和HEX荧光序列标签。
1.4 DNA提取和基因型检测
采用CTAB法提取种质的基因组DNA,稀释至10 ng·μL-1后利用浙江省农业科学院公共实验室的IntelliQube高通量基因分型检测平台进行SNP基因型检测,单点反应体积为1.6 μL,其中,样品DNA 0.8 μL,2×Master mix和Primer mix混合后的体积为0.8 μL。PCR扩增程序为95 ℃ 15 min;94 ℃ 20 s,61~55 ℃ 60 s(Touch-Down PCR,0.6 ℃每个循环),10个循环;94 ℃ 20 s,55 ℃ 60 s,26个循环。试验结果采用IntelliQube平台自带软件分析。
1.5 数据统计
试验数据统计分析采用Excel 2010软件,群体遗传多样性分析采用Tassel 5.0软件。
2 结果与分析
2.1 浙江地方菜豆种质分布
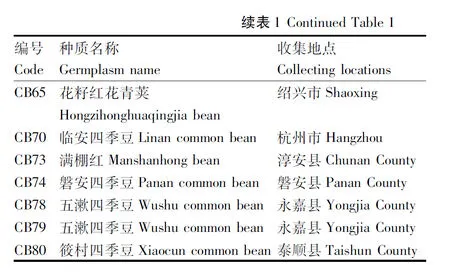
从44份种质的搜集地点分布情况来看,全省11个地级市中,除舟山外,其余10个地级市均收集到菜豆地方品种(图1),其中,杭州市收集到18份地方品种,数量位居全省首位,丽水和衢州也是收集到地方品种较多的地级市,分别收集到8份和6份,湖州、嘉兴和宁波均只收集到1份材料。从材料发现的地理环境来看,大部分材料位于山区,海拔较高,人口密度较小。这些材料均为农民自己留种繁殖,无野生型材料,而且均为菜用型菜豆。
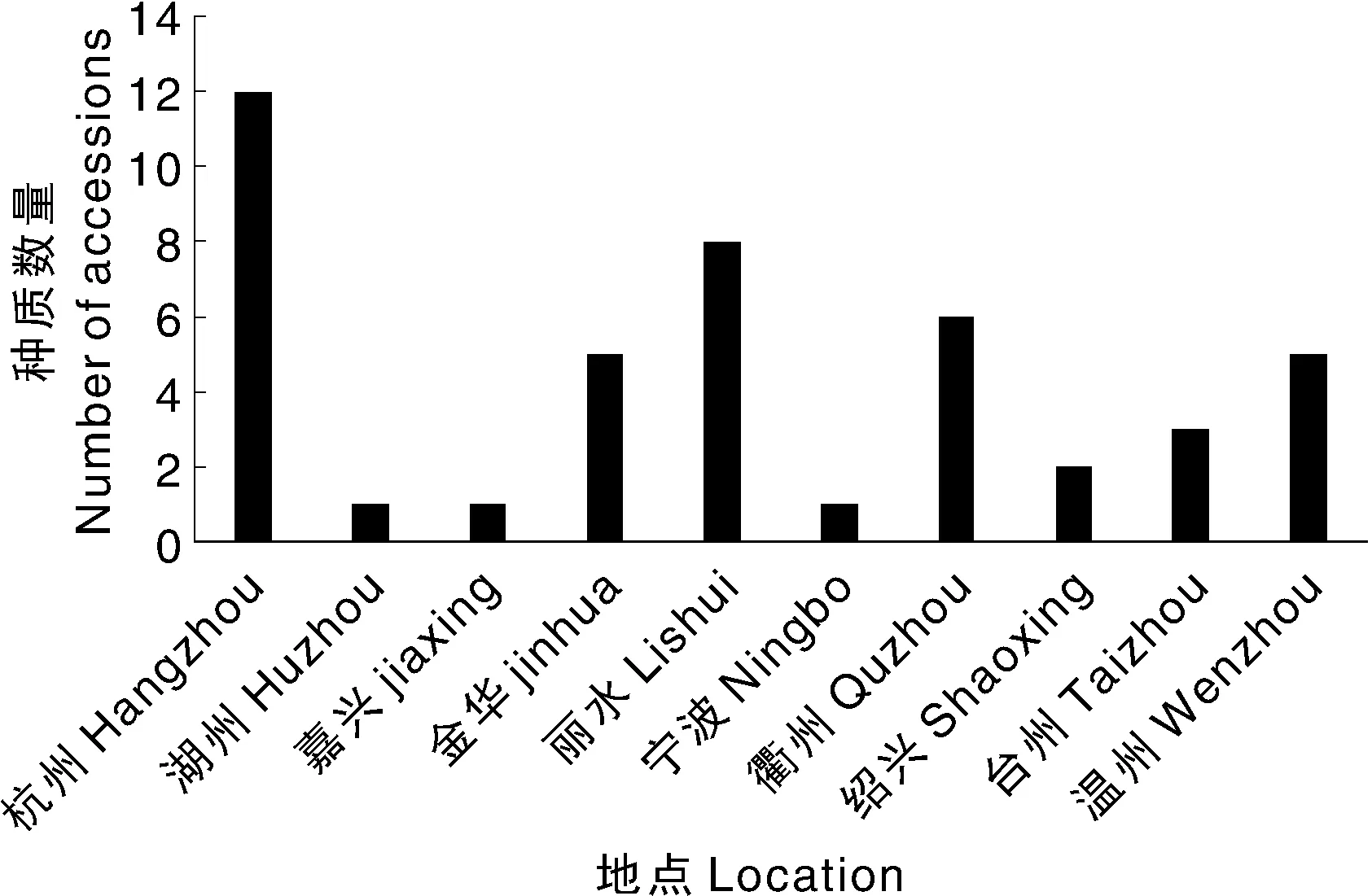
图1 浙江省地方菜豆种质分布Fig.1 Distribution of common bean germplasm in Zhejiang Province
2.2 浙江省地方菜豆种质形态多样性
通过对44份菜豆种质连续2 a的观察,发现这些种质在形态特征上变异丰富(图2)。44份种质中有2份材料为矮生类型,其他材料均为蔓生类型,说明浙江省菜豆种质以蔓生类型为主;在小叶叶形上,18份种质的叶形为近菱形(占比40.9%),26份种质的叶形为近圆形(占比59.1%);叶色呈现出浅绿色、绿色和深绿色,分别包含12、23和9份种质;嫩荚荚型以长扁条为主,有31份种质(占比70.5%),此外短扁条荚型的有2份,短圆棍型1份(花籽红花青荚),弯扁条型有3份,弯圆棍型4份,长圆棍型3份;嫩荚横切面呈现出“∞”字形、近圆形、桃形和长梨形,分别含有1、4、30、9份种质;嫩荚以绿色为主,含有24份种质(占比54.5%),黄绿色有1份,浅绿色有15份,绿白色有3份,紫红色有1份;种子形状以肾形和卵圆形为主,分别含有22份(占比50%)和20份(占比45.5%),还有2份为圆形;种皮色变异丰富,其中3份为白色,5份为茶褐色,6份为褐色,7份为黑色,2份为红褐色,1份黄色,2份黄绿色,3份乳白色,15份为双色。
2.3 主要农艺性状
除形态学特征外,本研究还调查了始花节位等5个重要农艺性状,发现44份种质在这些性状上呈现较大的多样性(表2)。从次数分布情况上看,这些性状在群体上分布均接近正态分布(图3)。44份种质的始花节位变异幅度为2.3~7.2,平均始花节位为3.9,其中,始花节位小于4的有31份种质(占比70.5%),始花节位大于5的有9份种质(占比20.5%),说明大部分种质开花节位较低,结荚较早。嫩荚长为11.8~21.3 cm,平均荚长为16.1 cm,有16份种质的荚长小于15 cm,3份种质的荚长超过20 cm,说明地方品种的豆荚普遍较短;嫩荚宽为0.7~1.7 cm,平均嫩荚宽为1.1 cm,有19份种质的荚宽小于1.1 cm;单荚重为6.0~16.6 g,平均单荚重为11 g,有22份种质(占比50%)的单荚重小于平均值,3份种质的单荚重超过15 g,说明地方品种的单荚重均较小;千粒重为144.56~528 g,平均值为271.6 g,千粒重小于250 g的种质有15份,大于400 g的种质有2份。综合44份种质的形态学和主要农艺性状发现,无法根据表型直接区分这些资源,需要进一步根据基因型研究其遗传多样性和亲缘关系。

表2 四十四份种质在5个农艺性状上的表现
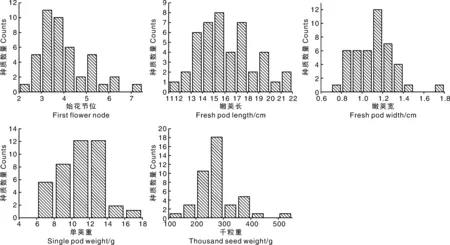
图3 五个主要农艺性状在44份种质中的分布频率Fig.3 Distribution frequencies of five traits of 44 common bean accessions
2.4 KASP标记开发
为了分析44份材料的亲缘关系,本研究从已经发表的BARCBean6K_3 BeadChip芯片[6]上按照SNP在基因组上的位置均匀选择185个SNP转化为KASP标记,经在44份种质中扩增后发现,有97对KASP标记扩增失败或在44份种质中无多态,有88对KASP标记在44份种质中扩增出2种等位变异信号(图4,表3)。这88对标记分布在11条染色体上,其中5号染色体上的标记数量最少,只有4个,8号染色体上的标记数量最多,达到12个(表4)。88个标记中有45个标记在所有种质中得到成功扩增,其余43个标记在群体中的缺失率最小为0.02,最大为0.30,各标记在群体中的多态性信息量值(PIC)变异幅度为0.044~0.726,平均PIC为0.25,最小等位变异频率(MAF)变异幅度为0.023~0.455,MAF平均值为0.12(表5)。

A,一种基因型;B,两种基因型。蓝色信号代表HEX等位变异,红色信号代表FAM等位变异。A, Single genotype; B, Two genotypes. Blue signal represented HEX allelic variation, red signal represented FAM allelic variation.图4 KASP标记在44份种质中的扩增信号Fig.4 Amplification signals for the KASP markers in 44 accessions

表3 本研究开发的88对KASP标记基本信息

续表3 Continued Table 3

续表3 Continued Table 3

续表3 Continued Table 3

表4 不同染色体上有多态标记的分布情况

表5 八十八个KASP标记在44份种质中扩增结果

续表5 Continued Table 5
2.5 浙江菜豆种质遗传多样性分析
利用44份种质的基因型数据进行主成分分析(PCA),结果显示,该群体大致可以分为2个主成分,其中以PC1为主,包含38份种质,PC2仅包含6份种质(图5)。系统进化树分析显示,44份种质大致可以分为2个类群,其中类群Ⅰ含有18份种质,类群Ⅱ含有26份种质(图6)。对比2个类群的基本性状发现,2份矮生型种质均在类群Ⅰ中,在小叶叶形、叶色、荚形上,2个类群间无明显的分化。在荚形上,2个类群都以长扁条为主,但类群Ⅰ还有其他荚形,变异较为丰富;在嫩荚横切面上,类群Ⅰ以长梨形和桃形为主,占比38.9%和50%,但类群Ⅱ以桃形为主,占比达到80.8%;在嫩荚主色和种子形状上,2个类群间亦无明显差异。在种皮色上,类群Ⅱ以双色为主,而类群Ⅰ的变异较为丰富,几乎含有所有类型的种皮颜色;在嫩荚节位、嫩荚长、嫩荚宽、单荚重上,类群Ⅰ的均值都略大于类群Ⅱ的均值;在千粒重上,2个类群间呈现显著差异,类群Ⅰ的千粒重较大,平均值为310.4 g,类群Ⅱ的千粒重普遍较小,平均值为244.7 g,因此,44份种质的类群分化与千粒重高度相关。这也与菜豆种质可以依据籽粒大小分为大粒类型(千粒重大于400 g)、中粒类型(千粒重为250~400 g)和小粒类型(千粒重小于250 g)的研究结果一致[11]。
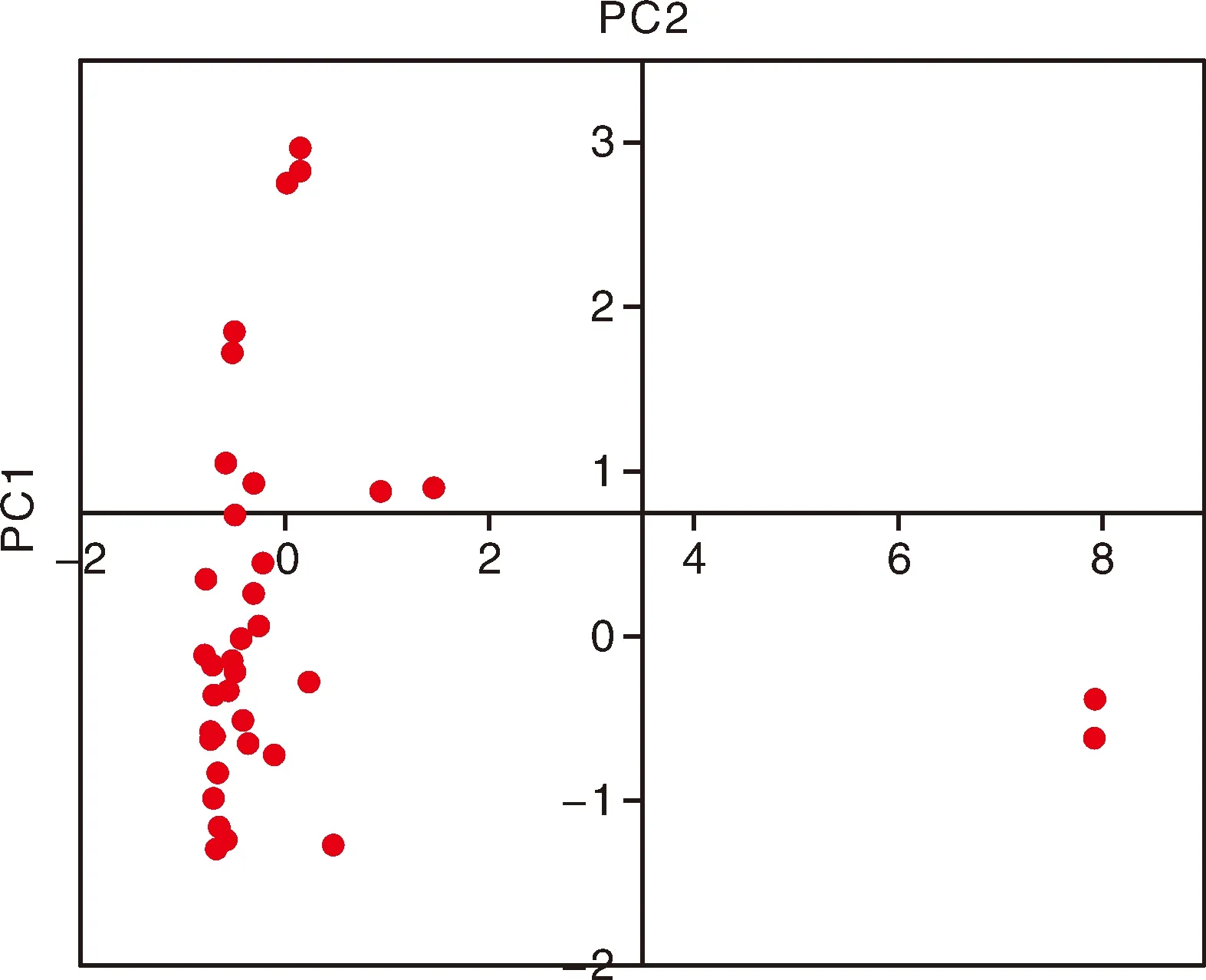
图5 四十四份菜豆种质的PCA分析结果Fig.5 PCA analysis result in the 44 common bean accessions
依据不同种质的千粒重,可以将类群Ⅰ进一步细分为3个小类(图6),其中类群Ⅰ-Ⅰ包含6份种质,千粒重变异幅度为353~528 g,平均千粒重为398.6 g,该类群与PC2中的种质完全一致;类群Ⅰ-Ⅱ含有4份种质,平均千粒重为349.3 g;类群Ⅰ-Ⅲ含有8份种质,平均千粒重为310.4 g,其中一份来自金华的磐安四季豆千粒重较小,只有154 g。类群Ⅱ可以进一步划分为2个小类,其中类群Ⅱ-Ⅰ含有15份种质,平均千粒重为231.8 g;类群Ⅱ-Ⅱ含有11份种质,平均千粒重为262.5 g,但其中一份来自杭州的临安四季豆千粒重较大,达到358 g。
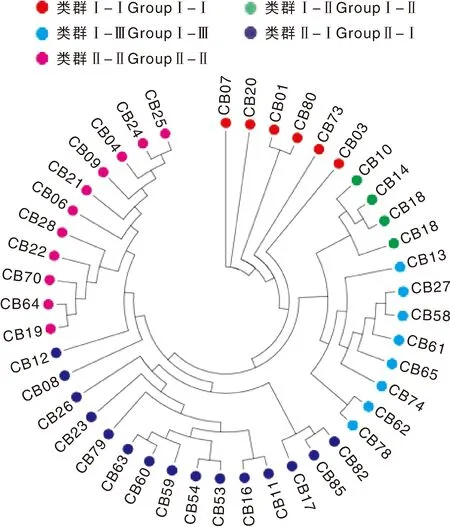
图6 四十四份菜豆种质的系统进化分析Fig.6 Phylogenetic tree analysis in the 44 common bean accessions
3 讨论
本研究使用的44份地方品种均为菜用型菜豆,其中28份种质为“第三次全国农作物种质资源普查与收集行动”所收集,这说明浙江省菜豆种质以食荚类型为主。通过对44份种质13个基本性状的调查,发现浙江省菜豆地方品种在生长习性、发育特性、嫩荚商品性及产量相关性状上均存在丰富的遗传变异,意味着地方资源中蕴含着优异的自然变异,可供育种利用。
依据种子大小等性状差异将普通菜豆分为安第斯和中美洲两大基因池[11]。千粒重超过400 g的大粒品种为安第斯基因池,千粒重小于250 g的小粒品种和千粒重介于250~400 g的中粒品种来源于中美洲基因池。利用种子蛋白、同工酶、形态学和DNA标记进行种质多样性分析得到的结果也支持两大基因池的划分[12-14]。我国普通菜豆是15世纪直接从美洲引入,可能首先在贵州、云南等省引种驯化,然后向东北传播,贵州、云南等省可能是普通菜豆的次级起源中心[1]。我国菜豆地方品种既有来自安第斯基因池的,也有来自中美洲基因池的,且安第斯基因池种质的遗传多样性高于中美洲基因池种质[2]。本研究中,44份菜豆种质的千粒重最小为144.56 g,最大为528 g,千粒重超过400 g的种质仅有2份,分别来自温州和杭州地区,说明我省菜豆种质主要以小粒和中粒类型的中美洲基因池为主。系统进化树分析可以将44份种质分为2大类,类群Ⅰ的平均千粒重为310.4 g,属于中粒类型,类群Ⅱ的平均千粒重为244.7 g,属于小粒类型。根据千粒重大小又可将类群Ⅰ细化为3个小类,类群Ⅱ细化为2个小类。尽管依据表型性状无法准确区分不同种质资源,但可以根据基因型了解不同种质的亲缘关系,进而结合其表型性状为育种中亲本选配提供理论依据。由于我省菜豆种质均为菜用型,因此本研究重点关注了荚部性状,分析发现,嫩荚宽、单荚重在两大类群间无显著性差异,嫩荚长在两大类群间虽有显著性差异,但平均荚长仅相差2.0 cm。因此,对于菜用型菜豆育种,可以适当的引入安第斯基因池的优异基因,通过亚种间杂交,选育突破性新品种。
本研究根据已经发表的菜豆BARCBean6K_3 BeadChip芯片中SNP序列信息,将185个SNP转化为KASP标记,其中88个KASP-SNP标记成功的用于44份种质的遗传多样性分析,其余的97对KASP-SNP标记中,4对引物在群体中扩增失败,可能是该SNP不适合转化成KASP标记,93对标记在群体中无多态,可能是群体太小,44份种质在该SNP位点上的等位变异一致。本研究是国内首次利用KASP标记研究菜用型菜豆遗传多样性的报道。无论如何,通过将芯片上SNP转化为KASP标记,为菜豆遗传多样性分析、基因定位和未来的分子育种提供了充足的标记资源。本研究通过对浙江地方菜豆种质的搜集和鉴定,梳理了浙江省菜豆种质基本信息,进一步丰富了我国菜豆表型信息数据库和遗传多样性,为地方资源的高效利用和新品种选育提供了科学依据。
