下调烟粉虱MED隐种BtabCSP6表达对番茄黄化曲叶病毒传播的影响
2022-11-28卢丁伊慧张战泓张德咏章松柏史晓斌
魏 艳,刘 勇,叶 倩,卢丁伊慧,张战泓,张 卓,张德咏,章松柏,史晓斌,*
(1.长江大学,农林病虫害预警与调控湖北省工程技术研究中心,湖北荆州 434025;2.湖南省农业科学院植物保护研究所,长沙 410125;3.湖南省农业科学院蔬菜研究所,长沙 410125)
番茄黄化曲叶病毒(tomato yellow leaf curl virus,TYLCV)是威胁番茄生长的主要病毒之一(刘馨等,2015),隶属于双生病毒科(Geminiviridae)菜豆金黄花叶病毒属(Begomovirus)(刘佰明等,2021),是具有孪生颗粒形态的单链环DNA病毒。TYLCV的发病症状主要表现为新叶先发病,叶片皱缩变小并向上卷曲,叶缘黄化,植株生长缓慢,无法正常开花结实并且影响果实色泽、口感和营养成分,使番茄的品质大大降低(王贤等,2021),素有“番茄癌症”之称。TYLCV在田间极易与番茄褪绿病毒(tomato chlorosis virus,ToCV)复合侵染(汪怡蓉等,2019),造成更严重的致病性,加速宿主植物死亡,不及时防治将会给农业生产造成重大损失(廖锦钰等,2021)。自然条件下TYLCV只能通过烟粉虱Bemisiatabaci传播,病毒泛滥往往伴随着烟粉虱的大暴发,因此控制烟粉虱种群数量可以有效减缓病毒传播。
烟粉虱属于半翅目(Hemiptera)粉虱科(Aleyrodidae),是危害番茄、黄瓜、辣椒、茄子、甘蓝等农业经济作物生长的主要害虫之一,其繁殖周期短、速率快,适应环境能力强,可以全年繁殖(李英梅等,2020)。烟粉虱可以携带并传播的植物病毒超过200种,为害600多种寄主植物(陈文斌等,2021),吸食植物汁液的同时传播植物病毒,造成寄主植物生长缓慢、无法正常结实,严重时导致植物死亡(刘国霞等,2021)。烟粉虱以持久性的传毒特性传播TYLCV:吸食带毒番茄植株后,病毒粒子通过唾液管和消化系统释放到血淋巴,最后再次回到唾液腺,在体内长时间保留。当带毒烟粉虱在未感染TYLCV番茄植株上取食时,体内的病毒粒子随着唾液通过口针一起进入植物体内,导致未感染TYLCV的番茄染病。此外,烟粉虱取食过程中分泌的蜜露会附着在叶片表面诱发煤污病(刘国霞等,2021)。因此,在防御手段上应该做好媒介昆虫的防治,切断传播途径(苑丽彩等,2016)。
化学感受蛋白(chemosensory proteins,CSPs)是一类小分子的可溶性蛋白,有4个保守的半胱氨酸形成两个非连锁的二硫键,能识别并结合周围环境中的信号分子(Calvelloetal.,2005),再传递给相应的受体,帮助昆虫感应外界信号。这类蛋白最初在美洲大蠊Periplanetaamericana的再生足上检测到,随后发现于黑腹果蝇Drosophilamelangaster的触角,之后苜蓿盲蝽Adelphocorislineolatus的不同感觉器官包括触角、唇须和喙等部位也发现了这类蛋白(Guetal.,2012),甚至少量分布于非感觉器官中。从病毒传播的角度出发,探究影响病毒传播时发挥作用的昆虫化学感受蛋白,发掘潜在的分子机制,可以有效控制病毒传播。因此,研究化学感受蛋白可以作为设计和开发新型害虫管理策略的重要分子靶标(Zhangetal.,2016;Wangetal.,2019)。本研究旨在探明烟粉虱的化学感受蛋白基因在雌雄成虫传毒过程中的表达差异,探明差异基因对病毒传播的影响,为控制病毒发生寻找新途径。
1 材料与方法
1.1 供试烟粉虱MED隐种
本实验使用的烟粉虱MED隐种成虫由湖南省农业科学院植物保护研究所提供,饲喂在棉花植株上,并用纱笼隔绝。养虫室温度保持在26±2℃,相对湿度控制为60%±10%,光周期设定14L∶10D。番茄(钻红美娜)种植的温室条件:温度26±2℃,相对湿度75%±5%,光周期16L∶8D。
1.2 烟粉虱MED隐种成虫获毒
使用TYLCV侵染性克隆方法获得带毒番茄植株:TYLCV侵染性克隆是由浙江大学生物技术研究所周雪平教授课题组构建并提供,为上海TYLCV分离株(TYLCV-SH2)。将含有TYLCV侵染性克隆的农杆菌Agrobacteriumtumefaciens菌液在YEP固体培养基上进行划线操作,28℃恒温培养箱(冠森生物科技有限公司,上海)倒置培养48 h;挑取单菌落,接种于含有卡那霉素(50 μg/mL Kan+)和利福平(25 μg/mL Rif+)的YEP液体培养基中,28℃恒温摇床200 r/min过夜培养,收集农杆菌菌液至50 mL无菌离心管中,离心机(Eppendorf,德国)转速调至4 000 r/min,室温离心10 min,去除上清液,底部菌体用无菌去离子水重新悬浮,使用分光光度计调节OD600为1.0,室温静置4 h后进行接种。用一次性无菌注射器吸取1 mL接种液注射至番茄真叶及茎杆中,8 h黑暗处理后转至温室继续培养,约3周后观察发病症状并通过PCR检测感染情况。
微虫笼收集未感染病毒的烟粉虱MED隐种成虫,夹在已感染TYLCV的番茄植株叶背面饲喂48 h,收集取食后的烟粉虱,使用蛋白酶K、树脂溶液提取单头烟粉虱MED隐种DNA。根据Taq DNA Polymerase (Mg2+Plus Buffer)试剂盒(南京诺唯赞生物科技股份有限公司,南京)的操作说明书,使用TYLCV特异性引物(廖锦钰等,2021)(表1),进行PCR扩增,反应体系:10×Taq Buffer (Mg2+plus) 2.5 μL,TYLCV正反向引物(10 μmol/L)各1 μL,dNTP Mix (10 mmol/L each) 0.5 μL,Taq DNA Polymerase (5 U/μL) 0.5 μL,模板DNA 1 μL,ddH2O 18.5 μL。扩增程序:95℃ 3 min;95℃ 15 s,56℃ 15 s,72℃ 1 min 30 s,35次循环;72℃ 5 min。扩增产物进行1%琼脂糖凝胶电泳检测,检测结果正确的PCR产物送至公司测序并进行序列比对,鉴定TYLCV。
从饲养的烟粉虱MED隐种中随机抽取20头进行种群鉴定,首先使用蛋白酶K、树脂溶液提取单头烟粉虱MED隐种DNA,使用2×Taq Plus Master Mix Ⅱ (Dye Plus)试剂盒(南京诺唯赞生物科技股份有限公司,南京)并结合特异性引物(刘微等,2019)(表1)进行PCR扩增。PCR反应体系:2×Taq Plus Master Mix Ⅱ (Dye Plus) 10 μL,上下游引物(10 μmol/L)各1 μL,模板DNA 1 μL,ddH2O 7 μL。扩增程序:95℃ 5 min;95℃ 15 s,53℃ 45 s,72℃ 1 min,35次循环;72℃ 10 min。扩增产物进行酶切,AseⅠ酶可将烟粉虱MED隐种的PCR产物酶切成两条链,烟粉虱MEAM1隐种的PCR产物则不能被切开(刘微等,2019)。
1.3 烟粉虱MED隐种成虫体内CSP基因相对表达量测定
人工自制吸虫管分别收集50头未感染和感染TYLCV的烟粉虱MED隐种雌雄成虫并提取RNA,根据HiScript®ⅡQ RT SuperMix for qPCR (+gDNA Wiper)试剂盒的说明书 (南京诺唯赞生物科技股份有限公司,南京)逆转录合成cDNA并定量至200 ng,使用AceQ®qPCR SYBRGreen Master Mix试剂盒(南京诺唯赞生物科技股份有限公司,南京)参考CSP基因序列(Zengetal.,2019),使用Primer Premier 5软件设计qBtabCSP1-8特异性引物(表1),于qTOWER3G荧光定量PCR仪(耶拿,德国)进行RT-qPCR。PCR反应体系:Nuclease-free Water 8.2 μL,正反向引物(10 μmol/L)各0.4 μL,cDNA 1 μL,5×HiScript Ⅱ qRT SuperMix Ⅱ 10 μL。扩增程序:95℃ 5 min;95℃ 10 s,60℃ 30 s,95℃ 15 s,40次循环;60℃ 1 min,95℃ 15 s。以未感染TYLCV的雌雄成虫体内BtabCSP1-8基因表达量为参照,采用比较周期阈值法(2-ΔΔCt)计算烟粉虱MED隐种雌雄成虫体内BtabCSP1-8基因的相对表达量。
1.4 RNAi效果测定
取100头未感染TYLCV的烟粉虱MED隐种成虫RNA为模板,按照1.3节的方法逆转录合成cDNA。合成dsGFP(GenBank 登录号:U70496.1)(对照组)的正反向引物(Luetal.,2021),使用Primer Premier 5软件设计合成dsBtabCSP6(处理组)的正反向引物(表1),使用Phanta®Max Super-Fidelity DNA Polymerase试剂盒(南京诺唯赞生物科技股份有限公司,南京)进行扩增,反应体系:ddH2O 10 μL,2×Phanta Max Buffer 12.5 μL,dNTP Mix (10 mmol/L each) 0.5 μL,正反向引物 (10 μmol/L)各0.5 μL,cDNA 0.5 μL,Phanta®Max Super-Fidelity DNA Polymerase 0.5 μL。扩增程序同1.2节,扩增产物纯化回收后使用pEASY®-T1 Cloning Kit试剂盒(北京全式金生物技术股份有限公司,北京)连接到pEASY-Blunt克隆载体获得重组子,转化到Trans1-T1感受态细胞(北京全式金生物技术股份有限公司,北京)中得到转化子,提取克隆质粒。用T7 RiboMAXTMExpress RNAi System试剂盒[普洛麦格(北京)生物技术有限公司]将目标DNA片段合成为dsRNA。纳米滴分光光度计[赛默飞世尔科技(中国)有限公司,上海]测定dsRNA的浓度和纯度。合成的dsRNA与15%的蔗糖混合配制成为饲喂液(dsRNA定量400 ng/μL,每管200 μL饲喂液),吸50头烟粉虱MED隐种雌成虫饲喂dsGFP(对照组)和dsBtabCSP6(处理组),48 h取样进行RT-qPCR,检测BtabCSP6的表达量(方法同1.3节),技术重复3次,生物学重复3次。
表1 引物信息Table 1 Primer information
1.5 BtabCSP6 RNAi后烟粉虱MED隐种获毒率和传毒率的测定
1.5.1获毒率的测定:选择未感染TYLCV的雌成虫,按照1.4节的步骤饲喂dsGFP(对照组)和dsBtabCSP6(处理组),参考1.1节的方法获毒不同时间(6,12,18,24,48和72 h),每个时间段各30头,检测不同时间段单头雌成虫获毒情况,根据每个时间段获毒雌成虫数量计算获毒率。获毒率=感染TYLCV烟粉虱总数/每组用于实验烟粉虱总数(30头)×100%。生物学重复3次。
1.5.2传毒率的测定:吸取1.4节饲喂dsGFP(对照组)和dsBtabCSP6(处理组)后48 h的烟粉虱MED隐种雌成虫,参考1.1节的方法获毒后,每组分别吸取1,5,10,25和50头烟粉虱,共5组,置于不同微虫笼中,微虫笼夹在长势相同的未感染TYLCV的5株番茄植株上传毒。每组实验番茄总数为25株,一周后移去微虫笼,番茄植株继续培养两周,使用1.1节的方法检测感染TYLCV番茄植物株数量,计算对照组与处理组的传毒率。传毒率=感染TYLCV番茄植株数量/每组用于实验的番茄植株总数(25株)×100%。生物学重复3次。
1.6 数据分析
用IBM SPSS Statistics 21(SPSS Inc.,Chicago,IL,美国)进行统计分析,采用独立样本T检验分析方法比较感染TYLCV的雌性与雄性成虫之间BtabCSP基因表达量差异显著性,RNAi处理前后不同时间段之间获毒率、不同数量级传毒率的差异显著性;使用作图软件 SigmaPlot 12.50和Graph Pad Prism 8作图。
2 结果
2.1 感染TYLCV后烟粉虱MED隐种体内BtabCSP1-8基因表达量变化
RT-qPCR结果表明,与未获毒成虫相比,获毒雌成虫中BtabCSP3和BtabCSP6表达量变化最为明显;雄成虫中BtabCSP4和BtabCSP6表达量变化最为明显,均可考虑为与TYLCV相关的差异表达基因。与获毒烟粉虱MED隐种雄成虫相比,获毒烟粉虱MED隐种雌成虫有4个基因(BtabCSP2,BtabCSP5,BtabCSP6和BtabCSP8)表达显著上调(P<0.05);与获毒烟粉虱MED隐种雌成虫相比,雄成虫有3个基因(BtabCSP1,BtabCSP3和BtabCSP7)表达显著上调(P<0.05),其中BtabCSP3与BtabCSP6表达量在雌雄成虫间差异极显著(P<0.001)。BtabCSP6在雌成虫中的表达量显著高于在雄成虫中的(P<0.001),且与未侵染TYLCV的雌雄成虫相比,在侵染TYLCV的雌雄成虫体内均表达量差异显著(P<0.05),最终挑选BtabCSP6进行后续实验(图1)。
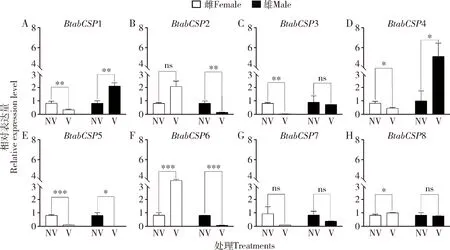
图1 TYLCV侵染后BtabCSP1-8 (A-H)在烟粉虱MED隐种雌雄成虫中的相对表达量Fig.1 Relative expression levels of BtabCSP1-8 (A-H) in female and male adults of Bemisia tabaci MED after TYLCV infectionNV:TYLCV未侵染烟粉虱TYLCV-noninfected B. tabaci;V:TYLCV侵染烟粉虱TYLCV-infected B. tabaci. 图中数据为平均值±标准误;柱上符号表示两组间的差异显著性(*P<0.05;**P<0.01;***P<0.001;nsP>0.05)(独立样本T检验)。下图同。Data in the figure are mean±SE.Symbols above bars indicate significance of difference between two groups (*P<0.05;**P<0.01;***P<0.001;nsP>0.05)(Independent samples T-test).The same for the following figures.
2.2 RNAi干扰BtabCSP6后基因表达量变化
结果显示,与饲喂dsGFP的对照组比较,饲喂dsBtabCSP6 48 h后雌成虫体内BtabCSP6的表达量显著下降(P<0.001)(图2),证明RNAi实验成功降低了基因的表达,干扰效果良好。
2.3 RNAi干扰BtabCSP6对烟粉虱MED隐种雌成虫获毒率和传毒率的影响
结果显示,与饲喂dsGFP的对照组比较,饲喂dsBtabCSP6 18-72 h的烟粉虱MED隐种雌成虫获毒率(图3:A)与传毒率(图3:B)均显著降低(P<0.05),其中,获毒率随着时间的延长呈上升趋势,对照组从48 h获毒率75%到72 h获毒率82%,RNAi干扰BtabCSP6后48 h获毒率为53%,比对照组24 h的获毒率高4%,在72 h后烟粉虱获毒率为59%,相比对照显著降低了23%(P<0.05)。传毒率随着番茄植株上烟粉虱MED隐种雌成虫数量的增多呈上升趋势。使用1头烟粉虱传毒时,对照组和处理组传毒率均低于10%;用5头烟粉虱传毒时,对照组和处理组传毒率分别为29%和16%;用10头烟粉虱传毒时,处理组的传毒率比对照组低23%;用25头烟粉虱进行传毒时,对照组传毒率达到92%,相比处理组的高出25%,二者差异极显著(P<0.01);用50头烟粉虱传毒时对照组传毒率几乎可达到100%,而处理组传毒率仅为80%,二者差异显著(P<0.05)。

图2 饲喂dsGFP和dsBtabCSP6后48 h烟粉虱MED隐种雌成虫中BtabCSP6相对表达量Fig.2 Relative expression level of BtabCSP6 in female adults of Bemisia tabaci MED at 48 h after feeding dsGFP and dsBtabCSP6

图3 饲喂dsGFP和dsBtabCSP6后48 h烟粉虱MED隐种雌成虫的获毒率(A) 和传毒率(B)Fig.3 Virus acqusition rate (A) and transmission rate (B) of female adults of Bemisia tabaci MED at 48 h after feeding dsGFP and dsBtabCSP6
3 讨论
本研究采用RT-qPCR检测烟粉虱MED隐种成虫感染TYLCV后体内BtabCSP基因表达量,结果显示不同BtabCSP基因的表达量差异较大(图1),这可能是不同基因参与不同功能造成的。例如,CSP在烟粉虱触角中表达与其参与挥发物的识别和检测有关,在胸部、足以及腹部的表达与觅食场所选择有关(Zengetal.,2019);此外,RNAi实验证明CSP除了参与昆虫嗅觉感知外还与昆虫发育有关,并且可以作为效应蛋白触发植物生理防御反应、作为润湿剂降低溶液表面张力(Liuetal.,2016;Zengetal.,2019)。本研究还发现不同BtabCSP基因在雌雄成虫之间差异较大(图1),这种差异首先会被认为是生殖因素的影响所致,因为雌成虫具备生殖产卵的特点且为二倍体,相比单倍体的雄成虫来说,核DNA含量也更加丰富,转录组分析结果也证明了较多功能性基因会在雌成虫中高表达(Guoetal.,2014),但本研究结果更倾向于感染TYLCV后BtabCSP基因表达量的变化。已有研究表明,烟粉虱雌成虫在叶背面的刺探时间高于雄成虫(卢少华等,2015),且无论是MED或MEAM1隐种,雌成虫的摄食能力均优于雄成虫,从而造成烟粉虱MED隐种雌成虫在TYLCV的获得和传播方面也更加有效(Guoetal.,2014)。因此,本研究以未感染TYLCV的烟粉虱雌雄成虫中BtabCSP基因的表达量作为参照,分析感染后BtabCSP基因表达量的差异(图1),确定感染TYLCV后雌成虫体内表达明显上调,且与雄成虫之间也有差异表达的基因,探明其对病毒传播是否造成影响。结果显示BtabCSP6在雌雄成虫之间的表达量具有显著差异,且在雌成虫中高表达(图1),因此推测BtabCSP6更可能成为烟粉虱MED隐种雌成虫获得与传播TYLCV过程中发挥作用的关键基因。烟粉虱雌成虫在生存能力、传毒效率、寄主适应性等方面均优于雄成虫,且个体数量会直接影响种群密度,这也是TYLCV会随着烟粉虱种群数量上升而大暴发的重要原因。探究雌成虫传播病毒过程中发挥作用的关键基因,并对其进行干扰,从而减少烟粉虱种群数量,阻碍病毒传播并减缓病毒蔓延,为防治番茄病害提供新的思路和理论依据,但其内在的分子机制仍需要深入研究。
本研究合成dsGFP和dsBtabCSP6并与15%的蔗糖混配为饲喂液饲喂烟粉虱MED隐种雌成虫,干扰靶标基因BtabCSP6的表达(图2),分析干扰前后烟粉虱对TYLCV的获毒率(图3:A)与传播率(图3:B)的结果可知,烟粉虱MED隐种雌成虫在不同获毒时间段的获毒率均比饲喂dsGFP的对照组低,且在48 h存在显著差异,不仅如此,获毒的烟粉虱MED隐种雌成虫在饲喂dsBtabCSP6 48 h后,不同数量烟粉虱的传毒率相比对照也大大降低,虽然干扰BtabCSP6并不能完全阻断病毒传播,但仍然考虑BtabCSP6对传毒获毒造成了一定影响,实验结果符合预期,证明BtabCSP6可以影响TYLCV的传播,BtabCSP6可能是与TYLCV传毒获毒相关的基因,但不一定是最关键的基因。之前研究表明,TYLCV侵染烟粉虱后,病毒粒子可以在体内循环至血淋巴,且存在于整个生命周期,植物感染TYLCV后防御反应被抑制,营养条件的改变反而更利于烟粉虱种群增长,因此,烟粉虱获毒2 d再传毒,侵染率基本可以达到100%(丁天波等,2021)。采用相同的RT-qPCR分析方法发现,烟粉虱MED隐种尤其是雌成虫,获得和传播TYLCV的能力均显著高于MEAM1隐种,表明病毒传播能力的差异不仅与生物型相关,还受性别的影响(Guoetal.,2014)。本研究中BtabCSP6对传毒获毒造成了一定影响,据报道,在昆虫味觉感受器中高度表达的CSP在摄食中可以作为疏水性营养物质的增溶剂以及口器表面活性剂,减少吮吸过程中的压力(Sánchez-Graciaetal.,2009),考虑本实验获毒率和传毒率的降低,其原因可能是干扰BtabCSP6后烟粉虱刺吸次数减少造成的。另有研究表明,在斜纹夜蛾Spodopteralitura中肠中高表达的CSP可能在特化和适应不同生态系统中发挥功能(Yietal.,2017),中肠是病毒在体内运输的一道重要屏障,也是病毒能否在昆虫体内存留的关键,因此BtabCSP6也可以考虑是否在中肠高表达,进而影响昆虫的获毒与传毒。本实验推测BtabCSP6是与TYLCV传毒获毒相关的基因,其抑制病毒传播的机理还有待深入挖掘。BtabCSP6有望成为减缓病毒传播的新靶标基因,RNAi也可以考虑用做控制烟粉虱MED种群的新方法,成为杀虫剂的替代品且对植物无毒害作用。
现在已经筛选出烟粉虱MED隐种中的许多基因可能成为RNAi的作用靶标,如Hormone-receptor-likein78 (BtabHR78),Ultraspiracle(USP),Nuclearreceptorsubfamily5groupAmember1 (NR5A1),Hormonereceptor-likein39 (HR39),Estrogen-relatedreceptor(ERR)和Pannier(PNR)(Heetal.,2020);很多基因成为RNAi的靶标候选基因:乙酰胆碱受体α亚基、α葡萄糖苷酶、水通道蛋白、热休克蛋白、海藻糖酶和海藻糖转运蛋白基因等(Vyasetal.,2017)。干扰BtabCSP6表达可以从传播过程阻碍病毒获取和扩散,因此化学感受蛋白除了可以识别并结合周围环境中的信号分子使机体做出相应的反应外,还可以与烟粉虱体内相应的受体相互作用控制病毒在体内的传导。本实验发现BtabCSP6影响了TYLCV的传播,未来可以定位与BtabCSP6相互作用的受体,阐明CSPs与病毒之间相互作用的分子机制,开发新的生物杀虫剂来防治烟粉虱。
