应用仿生模式分析光环境对蜘蛛抱蛋光合特性及生物量分配的影响1)
2022-11-28谭锦豪于耀泓罗晓茼
谭锦豪 于耀泓 罗晓茼
(广东省森林植物种质创新与利用重点实验室(华南农业大学),广州,5106421)
王冉 王玥 苏艳 何茜
(河南省退耕还林和天然林保护工程管理中心) (广东省森林植物种质创新与利用重点实验室(华南农业大学))
岭南(我国南方五岭以南地区的统称)地区以低山丘陵为主,具有丰富的林地资源和温暖湿润的气候,适宜大量推广林下种植[1],其中林药复合经营模式是最具特色的模式之一[2]。林药复合生态系统中,光照被认为是最重要的单一限制性因子,林分冠层结构的异质性影响太阳辐射进入林内,改变林下的光照强度和光质[3-4]。北美云杉(Piceasitchensis)林分内垂直高度的光强和光质随着叶面积指数上升,林下光量子通量密度、红光和蓝光下降,而远红光增加[5]。Wei et al.[6]分析了6种森林类型下辽东楤木(Araliaelata)的碳水化合物含量发现,随着郁闭度增加,林下相对红光比值降低,促进辽东惚木叶片淀粉积累。郑芬等[7]以不同林龄的天然常绿阔叶林为研究对象,通过研究冠层结构与林下光环境的关系,发现随着冠层叶面积指数增加,红光与远红光的比值下降,蓝光与红光比值增加。
植物改变自身光合特性和生物量分配格局是植物种群适应光环境的重要途径,对植物在不同光环境中存活和提高其自身竞争力至关重要[8];阴生植物在林下低光环境表现耐阴的生态策略,影响着植物的生长和光合特性[9]。Calzavara et al.[10]对4种耐阴植物的光合特性进行了研究,发现阴生植物可以通过改变叶表型形态特征(如叶长、叶宽、叶面积)增加光子捕获能力,增加叶绿素质量分数提高光能利用能力,同时降低光补偿点和光饱和点适应弱光环境;He et al.[10]对比了中国亚热带常绿森林中57种主要木本植物的光合特性和叶片寿命,结果表明耐阴植物在弱光环境表现叶片寿命更长。吴统贵等[11]研究了香樟(Cinnamomumcamphora)林下栽培的红茴香(Illiciumhenryi)光合特性发现,随着林分郁闭度增加,红茴香通过增加表观量子效率、降低光补偿点和光饱和点以适应弱光环境。
蜘蛛抱蛋(AspidistraelatiorBlume)为单子叶植物纲、百合科(Liliaceae)、蜘蛛抱蛋属(Aspidistra)的植物,别名一叶兰、蛇退、万年青等,分布于我国长江以南地区[12],常生于荫蔽、湿度大的天然林下。其根状茎是一种传统民间草药,具有活血止痛、清肺止咳等功效,具有较高的药用价值[13]。目前关于光照强度和光质对植物影响的研究以人工光源为主[14-16],而关于野外自然光对植物生长的研究较少。基于此,本研究选取不同郁闭度的红锥混交林下种植蜘蛛抱蛋,对蜘蛛抱蛋的光合特性和生物量分配策略进行研究,以期揭示蜘蛛抱蛋在荫蔽环境下的生理生态适应性策略,为选择蜘蛛抱蛋仿野生栽培模式提供技术支持。
1 研究地概况
研究地位于广东省龙眼洞林场帽峰山工区,海拔470 m,隶属亚热带季风气候,年平均气温为21.5~22.2 ℃,年平均年降水量为1 800~2 000 mm,年降水时间150 d左右,年日照时间1 473~1 925 h。研究地属低山丘陵,土壤类型为赤红壤,土层较厚。
2 研究方法
2.1 试验地设置
研究林分为35年生的红锥混交林,主要树种有红锥(Castanopsishystrix)、构树(Broussonetiapapyrifera)、木荷(Schimasuperba)、山油柑(Acronychiapedunculata)。通过目测法和半球摄影法选择不同郁闭度(CD)的林分,林下种植蜘蛛抱蛋,试验设置3个处理:试验地A郁闭度≤0.6,试验地B郁闭度为0.6~0.8,试验地C郁闭度≥0.8(见表1)。2018年6月,在每个试验地种植苗高一致,且生长良好的蜘蛛抱蛋幼苗,栽植株行距30 cm×30 cm,每个处理设置3个小区,小区面积为400 m2。

表1 不同试验地及林分特征
2.2 冠层结构参数
2018年5—6月选择阴天或晴天日落后,使用Nikon D750数码相机外接鱼眼镜头摄取红锥混交林冠层结构,用三脚架将设备水平放置离地面1.3 m处,镜头方向向北,避免灌木和人员进入拍摄区。每个小区在中心点和对角线四分位处各拍摄3张冠层图像,每个小区共拍摄15张。拍取的图像采用Gap Light Analyzer 2.0(GLA)图形处理软件分析林冠影像,得到林冠郁闭度(CD)、叶面积指数(LAI)、林下直射光量(Tdir)、林下散射光量(Tdif)和林下总光照量(Ttot)。
2.3 林下光环境
2018年11月和2019年2、5、8月选择连续的晴天,在09:00—11:00,使用植物光照分析仪(PLA-30,中国)测定林下光环境。每个样地以“S”型选取9个位置对林下光环境进行测定,每个位置重复3次测定。测定时将仪器水平放置于垂直地面50 cm处,定时测量3 min,取其平均值。测定过程避开人员、灌木以及林内光斑。测定光谱波长380~780 nm的光量子通量密度(PPFD),按照不同波段分为红、绿、蓝三种光质,红光波长为600~780 nm,绿光为500~600 nm,蓝光为380~500 nm。分别计算红光、绿光和蓝光占总光照光量子通量密度的百分比,即红光比(RRP)、绿光比(RGP)和蓝光比(RBP)。
2.4 蜘蛛抱蛋生长及生物量
2018年10月—2019年10月,每月从各小区选取生长基本一致的蜘蛛抱蛋幼苗20株,测定其株高、叶长、叶宽及分枝数。
2019年10月,于每个试验地采集30株幼苗(每个小区各10株),带回实验室,杀青后烘干至恒质量。测定根生物量(RB)、茎生物量(SB)、叶生物量(LB)和总生物量(TB)。计算各部位占总生物量的比值:叶生物量比(LBR)、茎生物量比(SBR)、根生物量比(RBR)。
2.5 蜘蛛抱蛋叶片形态参数
2019年10月,每个小区采集10株蜘蛛抱蛋的叶片,使用叶面积扫描仪(LA-S,中国)测定叶面积参数。测定指标为单株总叶面积(LA)、比叶面积(SLA)、叶长(LL)、叶宽(LW)。
2.6 叶绿素质量分数的测定
2019年10月使用乙醇浸泡法测定蜘蛛抱蛋叶绿素质量分数。称取剪碎的新鲜叶片0.2 g于95%乙醇浸泡24 h至叶片变白,将提取液倒入比色杯中,在665、649、470 nm波长下测定其消光度值,以95%乙醇为空白对照,计算得到叶绿素a质量分数(Chl a)、叶绿素b的质量分数(Chl b)、类胡萝卜素质量分数(Car)和总叶绿素质量分数(Chl a+b)。每个处理重复3次。
2.7 光响应曲线测定
2018年11月和2019年2、5、8月选择连续晴天,在09:00—11:00,使用Li-6400便携式光合作用分析系统(Li-Cor,Inc,USA)测定蜘蛛抱蛋的光响应曲线。设定红蓝光源控制光合有效辐射(PAR)范围在0~2 000 μmol·m-2·s-1,将光合有效辐射梯度设置为:2 000、1 800、1 500、1 200、1 000、800、600、400、200、100、50、20、0 μmol·m-2·s-1,每个梯度测定时间为2~3 min,测定时叶片温度为25 ℃,CO2摩尔分数固定为400 μmol·mol-1。每个小区随机选择3株,叶片选用植株顶部的成熟功能叶。每种植物用饱和光强进行光诱导直至净光合速率(Pn)稳定(大约10 min),使用光响应曲线自动测试程序测定植物光合参数(净光合速率(Pn)、胞间二氧化碳摩尔分数(Ci)、气孔导度(Gs)、蒸腾速率(Tr))。光能利用效率(LUE),LUE=(Pn/PAR)×100%[17]。
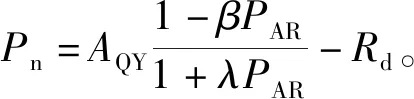
2.8 数据处理
使用SPSS 20.0软件完成数据基本描述统计、单因素方差分析、Duncan多重比较及相关性分析。Origin Pro 2021作图。试验数据表示为平均值±标准偏差。
3 结果与分析
3.1 冠层结构参数及林下光环境差异
3.1.1 冠层结构参数
由表2可知,试验地A郁闭度为0.49~0.61,试验地B郁闭度为0.68~0.78,试验地C郁闭度为0.83~0.86,试验地C显著高于试验地A和试验地B(P<0.05)。试验地C的叶面积指数最大(2.37),分别为试验地A和试验地B的1.85和1.62倍,但试验地A和试验地B叶面积指数差异不显著(P<0.05)。林下直射光量(Tdir)、林下散射光量(Tdif)和林下总光照量(Ttot)试验地A最大,试验地C最小,且差异显著(P<0.05),即随着郁闭度增加,林下直射光量、林下散射光量和林下总光照量减少。

表2 不同试验地冠层结构参数
3.1.2 不同季节林下光照强度
由表3可知,3个试验地林下光量子通量密度(PPFD)季节变化规律为:由冬季到秋季逐渐上升,秋季时最高,冬季时最低。试验地A和试验地B秋季的林下光量子通量密度显著高于冬季(P<0.05),但试验地C在4个季节的林下光量子通量密度之间差异不显著。在同一季节中,林下光量子通量密度最大为试验地A,最小的为试验地C,即随着郁闭度增加,林下光量子通量密度显著降低;秋季时,试验地A的林下光量子通量密度达到最大(51.48 μmol·m-2·s-1),分别为试验地B和试验地C的1.26倍和2.93倍。

表3 不同试验地林下光量子通量密度的季节变化
3.1.3 不同试验地的林下光质
由表4可知,随着郁闭度增加,林下光质呈红光下降,绿光上升的趋势。试验地C红光比值为15.27%,显著低于试验地A(16.21%)和试验地B(16.49%),但试验地A和B之间差异不显著。试验地C的绿光比值显著高于A和B(P<0.05)。3个试验地蓝光比值都在6%~8%,即郁闭度对蓝光比值的影响不显著。
3.2 蜘蛛抱蛋生长及生物量分配
3.2.1 蜘蛛抱蛋株高和分枝数
由图1可知,株高随着时间的增加而增加,3个试验地株高生长呈逐渐上升的趋势,其中前3个月株高增长迅速,后增速下降至平缓。3个试验地分枝数整体呈现先增加后降低的趋势,10—12月时增速缓慢,到2—5月增速最大,后增速逐渐放缓,到8—9月时分枝数呈下降的趋势。不同试验地,株高、分枝数试验地C最大,试验地A最小,即随着郁闭度增加,蜘蛛抱蛋株高和分枝数都表现出增加的趋势。

表4 不同试验地光质比较

图1 不同试验地苗木株高和分枝数
3.2.2 蜘蛛抱蛋生物量分配
由表5可知,蜘蛛抱蛋总生物量随着郁闭度增大而增加,不同试验地蜘蛛抱蛋总生物量试验地C最大,试验地A最小。郁闭度对蜘蛛抱蛋叶、茎和根生物量也产生显著影响(P<0.05)。试验地B和试验地C叶干质量显著高于试验地A(P<0.05),但试验地B和试验地C间差异不显著;试验地C茎生物量最大(6.37 g),显著高于试验地A和试验地B(P<0.05);3个试验地蜘蛛抱蛋根干质量在2.6~3.2 g,差异不显著。
由表6可知,不同试验地的植株体内生物量分配存在差异。3个试验地的叶生物量比差异不显著。茎生物量比在3个试验地差异显著(P<0.05),其中试验地C最大,试验地B次之,试验地A最低。而但根生物量比的结果相反,试验地A最大,试验地B次之,试验地C最低。

表5 不同试验地蜘蛛抱蛋的生物量

表6 不同试验地蜘蛛抱蛋生物量分配
3.3 蜘蛛抱蛋叶片形态参数
由表7可知,郁闭度对蜘蛛抱蛋单株叶面积有显著影响(P<0.05),其中试验地C最大,试验地B次之,试验地A最低。比叶面积随着郁闭度增加呈现“增加”的趋势,但不同试验地之间差异不显著。3个蜘蛛抱蛋单叶叶长在37~44 cm,叶宽在9.5~13 cm,不同试验地无显著影响。

表7 不同试验地蜘蛛抱蛋叶片形态参数
3.4 蜘蛛抱蛋叶绿素质量分数
由表8可知,蜘蛛抱蛋叶绿素a、叶绿素b、类胡萝卜素和总叶绿素随着郁闭度增加而增加,试验地C最大,试验地A最小,试验地C的叶绿素a、叶绿素b、类胡萝卜素和总叶绿素分别是试验地A的1.76、1.03、1.32、1.44倍,分别是试验地B的1.37、1.41、1.25、1.38倍,但试验地A和试验地B之间差异不显著。3个处理w(叶绿素a)∶w(叶绿素b)为1.72~1.75,差异不显著;w(类胡萝卜素)∶w(总叶绿素)试验地A和试验地B显著高于试验地C。

表8 不同试验地蜘蛛抱蛋叶叶绿素质量分数
3.5 光合特性及光能利用效率
3.5.1不同试验地蜘蛛抱蛋光响应曲线及光合特性季节变化
由图2、表9可知,不同处理下的蜘蛛抱蛋光响应曲线趋势一致,即在光强为0~800 μmol·m-2·s-1时逐渐上升,至光补偿点(LCP)后光合速率(Pn)达到最大值,随后开始下降至趋于平缓。通过拟合曲线计算光响应参数,不同试验地的蜘蛛抱蛋表观量子效率(AQY)、暗呼吸速率(Rd)、最大光合速率(Pn,max)、光饱和点(LSP)和光补偿点(LCP)显著(P<0.05)。随着郁闭度增加,最大光合速率呈现逐渐增加的趋势,表观量子效率、暗呼吸速率、光饱和点和光补偿点整体呈现下降的趋势。比较3个试验地不同季节下的最大光合速率(Pn,max),均为秋季最大,试验地B和试验地C的最大光合速率显著高于试验地A。3个试验地的光饱和点为800~1 200 μmol·m-2·s-1,光补偿点为1.5~10 μmol·m-2·s-1,试验地A的光饱和点和光补偿点整体高于试验地B和试验地C。

表9 不同试验地蜘蛛抱蛋光合特性参数

图2 不同试验地蜘蛛抱蛋光响应曲线季节变化
3.5.2 不同试验地蜘蛛抱蛋光能利用效率
由图3可知,3个试验地蜘蛛抱蛋的光能利用效率的光响应过程基本相似,为明显的单峰曲线。低光强(PAR<50 μmol·m-2·s-1)条件下,随着光合有效辐射的增强,光能利用效率急剧上升,对光强变化响应敏感;在PAR=50 μmol·m-2·s-1光强范围附近达到峰值,此后光能利用效率逐渐下降;当PAR>500 μmol·m-2·s-1时,光能利用效率逐渐趋于平缓。光能利用效率在低光强时,试验地C在全年光能利用效率整体大于试验地A处理,但两种处理间差异不大,表明蜘蛛抱蛋能够高效地利用弱光。

图3 不同试验地蜘蛛抱蛋光能利用效率的季节变化
3.6 各指标的相关性
3.6.1 光强、光质与冠层结构参数相关性
由表10可知,林冠郁闭度、叶面积指数与光量子通量密度、红光比都呈显著负相关(P<0.01),但与绿光比、蓝光比呈显著正相关(P<0.01)。林下直射光量、林下散射光量和林下总光照量与光量子通量密度、红光比呈显著正相关(P<0.01),与绿光比、蓝光比呈显著负相关(P<0.01)。
3.6.2蜘蛛抱蛋生物量分配、叶形态参数与光强、光质相关性
由表11可知,蜘蛛抱蛋总生物量与光强、光质相关性不显著。但光质影响茎生物量分配,红光比与茎生物量、茎生物量比呈极显著负相关(P<0.01),绿光比与茎生物量、茎生物量比呈极显著正相关(P<0.01),蓝光比与茎生物量比呈极显著正相关(P<0.01)。蜘蛛抱蛋单株总叶面积受光强光质影响,单株总叶面积与光量子通量密度呈显著负相关(P<0.05),与绿光比呈极显著负相关(P<0.01);但比叶面积、叶长、叶宽与光强、光质相关性不显著。
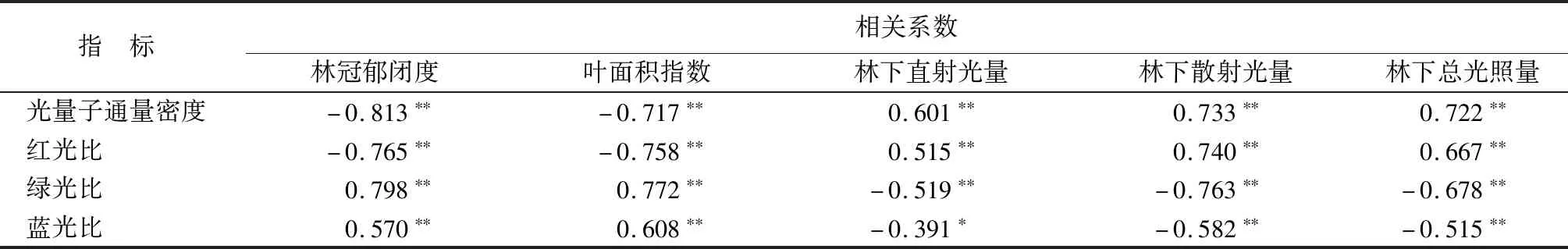
表10 光强、光质与冠层结构参数的相关系数
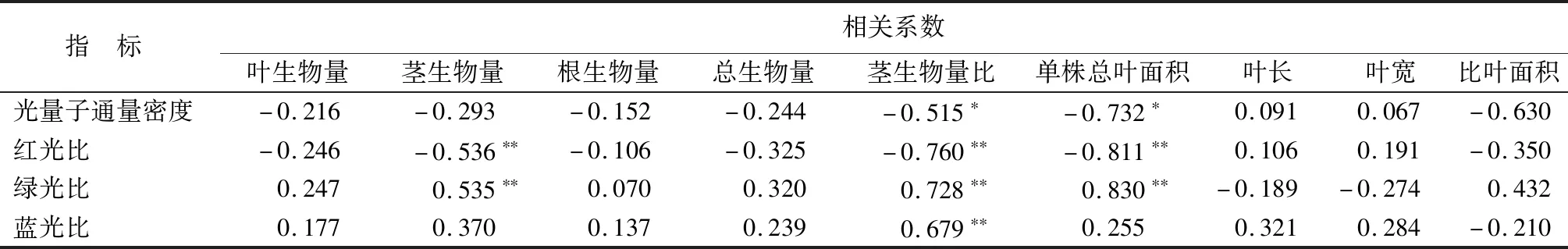
表11 蜘蛛抱蛋生物量分配、叶形态参数与光强、光质的相关系数
3.6.3蜘蛛抱蛋光合特性和叶绿素质量分数与光强、光质的相关性
由表12可知,蜘蛛抱蛋光合特性中表观量子效率、最大光合速率与光强、光质具有相关性。表观量子效率、最大光合速率与光量子通量密度、红光比呈负相关,与绿光比、蓝光比呈正相关;对叶绿素而言,蜘蛛抱蛋叶绿素质量分数(Chl a、Chl b、Chl a+b)和类胡萝卜素与光量子通量密度、红光比呈极显著负相关(P<0.01),与绿光比、蓝光比呈极显著正相关(P<0.01)。
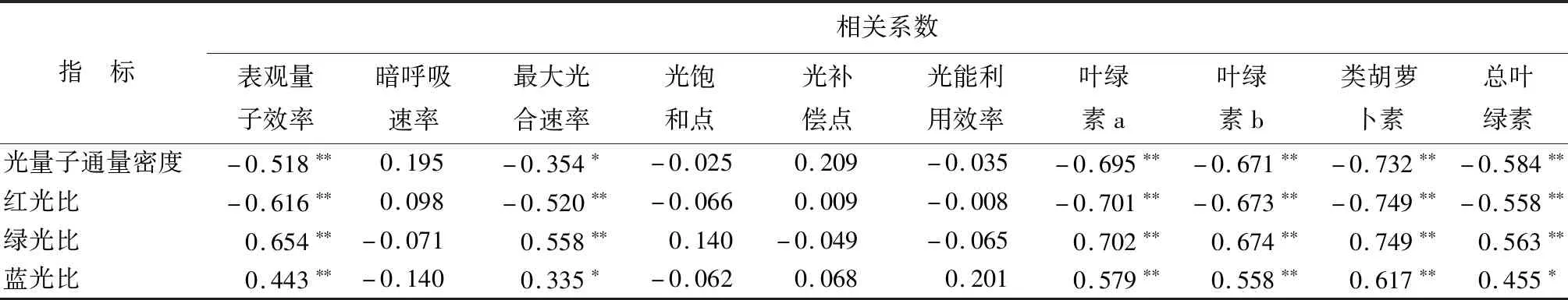
表12 蜘蛛抱蛋光合特性和叶绿素与光强、光质的相关系数
4 结论与讨论
4.1 冠层结构对林下光强光质的影响
冠层结构是森林生态系统与外界大气环境接触最紧密的介质,十分敏感且活跃[19]。森林植被组成影响森林冠层结构,冠层结构特征的差异影响森林生态系统能量循环的过程,尤其对太阳辐射截获的影响尤为明显[20]。林分结构的异质性影响着林下光环境,对光强光质有显著的影响[21]。本研究中,林下光量子通量密度减少程度随郁闭度增大而增大,郁闭度为0.6的林分,林下光量子通量密度比郁闭度为0.7时高0.69倍,但比郁闭度为0.8的林下光量子通量密度高2倍,表明随着叶面积增加,冠层对太阳辐射消光能力随之增加,这与黄润霞等[22]研究结果一致;不同处理间光量子通量密度季节变化也不一致,试验地A和试验地B冬季光量子通量密度最低,且与秋季差异显著,但试验地C林下光量子通量密度为13~18 μmol·m-2·s-1,季节间无显著差异,说明红锥混交林郁闭度≥0.8时,林下光量子通量密度趋于稳定。本研究冠层结构对林下光质的影响主要以对红光和绿光的影响为主,随着郁闭度增大,林下红光呈下降的趋势,绿光呈上升趋势,对蓝光无显著影响;同时,郁闭度0.6~0.7的红光比和绿光比没有显著差异,这与郑芬等[7]研究红锥林混交林光质时,郁闭度0.6的不同林龄林分红光比和绿光比存在差异的结果不一致,原因是林分组成及林龄差异导致的冠层消光系数差异。本研究中红光比与蓝光比、绿光比与蓝光比差异显著,光质的差异影响着林下植物的生长和叶片形态。
4.2 光强光质对蜘蛛抱蛋生长及生物量分配的影响
环境因子的累积效应对植被的生长生理产生巨大的影响[23],林下生长的植物以受光强、光质的影响尤为突出。植物利用太阳光谱波段为380~700 nm进行光合作用,植物对CO2固定能力体现叶片形态特征和植株生物量分配特征[24]。处于弱光环境中,植物通过增加叶面积以提高光能吸收率、捕获率及利用率[25]。本研究中,光环境的差异对蜘蛛抱蛋株高、单叶生长和比叶面积无显著影响,但对分枝数影响显著,光量子通量密度和红光比与叶面积指数呈极显著负相关,即叶面积指数随着光量子通量密度和红光比降低而增加。弱光环境的蜘蛛抱蛋通过增加分枝数和延长叶片寿命的方式增加叶片总面积,从而提高净光合生产力,与有关[26]研究结果一致[26-27],说明植物在不同光环境受到的影响可以反映到叶片表型形态上,并通过改变叶片形态以适应环境。当植物受到光照条件限制时,植物将更多的资源分配到地上部分,通过增加叶生物量分配以获取更多光照资源[27]。蜘蛛抱蛋生物量特征与环境光强、光质有显著相关性,光量子通量密度与蜘蛛抱蛋的生物量呈显著负相关,说明弱光环境促进生物量增加,生物量增加部分主要是叶生物量的增加。对于药用部位根状茎而言,光质对其的影响更大,红光比与茎生物量呈负相关,而绿光比与茎生物量呈正相关。本研究说明光质影响药用植物产量,降低红光和增加绿光可提高蜘蛛抱蛋药用部位的产量。
4.3 光强光质对蜘蛛抱蛋叶绿素质量分数的影响
叶绿素吸收光合作用的光能与光合速率直接相关。w(叶绿素a)∶w(叶绿素b)比值与光吸收能力的平衡有关,w(类胡萝卜素)∶w(总叶绿素)的比值反映了植物叶片光吸收和光损失之间的关系[28]。在试验过程中,随着光照强度的增加,蜘蛛抱蛋的叶片颜色由深绿色变为浅绿色到黄绿色,叶片灼烧现象清晰可见,在林窗处的叶片出现许多棕色斑点,并且叶片出现灼烧脱落的现象。而在弱光环境的蜘蛛抱蛋表现出更长叶片寿命和更高的叶绿素质量分数,来最大限度降低能量消耗,提高净光合生产力,这与前人的研究一致[29]。当郁闭度≥0.8时,蜘蛛抱蛋叶绿素质量分数最高,蜘蛛抱蛋的叶绿素质量分数(Chl a、Chl b)随着光照强度增加而显著降低,试验地A的叶绿素质量分数只有试验地C的70%。对于耐阴植物,强光可能会使光合系统严重损害或者完全失活。本文通过研究蜘蛛抱蛋的光合特性发现,光量子通量密度降低使蜘蛛抱蛋叶绿素质量分数提高,植株体内的叶片中色素蛋白复合体增加,提高了吸收与转化光能的能力。一般情况下,强光环境生长的植物比弱光环境有更高的w(叶绿素a)∶w(叶绿素b)比值,这与PSII对光子捕获能力有关,从而改变了植物的羧化效率[30]。但发现w(叶绿素a)∶w(叶绿素b)比值在不同光环境水平下无显著差异,说明蜘蛛抱蛋不能调整叶绿素a和叶绿素b的相对质量分数以适应高光环境。而其在强光环境中通过增加w(类胡萝卜素)∶w(总叶绿素)的比值来降低叶子的光吸收,以保护光合系统免受损害,提高对高光的适应能力[31]。
4.4 光强光质对蜘蛛抱蛋光合特性的影响
植物光合作用是将光能转化为化学能的过程[32],植物光合作用的表征是植物光响应曲线的拟合,光响应曲线描述了植物光合作用的速率如何随光增加而变化,表现了叶片的光合特性信息[33]。当叶片吸收光不能被最佳利用时,植物会受到光抑制,会导致光合作用效率和光氧化能力降低[34]。在本试验中反映为蜘蛛抱蛋最大光合速率随着光量子通量密度增加而降低。表观量子效率是揭示植物光化学效率的重要参数,反映了植物对弱光的利用能力[35]。蜘蛛抱蛋在郁闭度≥0.8的表观量子效率为0.05~0.42,在郁闭度≤0.6的表观量子效率为0.01~0.05,表明蜘蛛抱蛋对光十分敏感,对弱光有较强的利用能力,而强光会造成蜘蛛抱蛋光抑制。植物暗呼吸效率表现植物呼吸消耗的能力。随着光量子通量密度下降,蜘蛛抱蛋暗呼吸效率也降低,有利于提高植株净光合生产力,这与吕钦扬等[36]对八角林下金花茶的研究结果一致。光补偿点和光饱和点反映植物对光环境的适应能力,光补偿点较低的植物适应荫蔽环境生长。蜘蛛抱蛋的光补偿点和光饱和点随着郁闭度增加而降低,通过降低光补偿点和光饱和点来提高对弱光的能力。光能利用能力(LUE)是评价植物光能利用能力的重要参数,更是影响生态系统生产力的重要指标。3个不同处理的蜘蛛抱蛋光能利用能力在光合有效辐射为50 μmol·m-2·s-1时达到峰值,且随着光量子通量密度下降蜘蛛抱蛋光能利用能力更高。表明蜘蛛抱蛋为极耐阴植物,高光通过降低叶绿素质量分数导致叶片捕获光子能力下降,从而抑制其光合能力;另一方面,高光可能导致PSII结构损坏,减少核酮糖二磷酸缩化酶活性,增加暗呼吸和光呼吸效率,进一步使净光合速率下降。
光决定着植物生存、生长和分布,植物形态和生理可塑性归因于对光照适应能力。本文通过对不同光环境处理下蜘蛛抱蛋的生长生理进行比较,发现随着冠层郁闭增大和叶面积指数增加,林内光量子通量密度降低,林下生长的蜘蛛抱蛋通过增加总叶面积和提高叶绿素质量分数适应弱光,同时增加表观量子效率,降低光饱和点和暗呼吸速率来提高弱光利用能力,促进植株生物量的增加;红光比值下降和绿光比值上升有助于提高叶绿素质量分数、总叶面积和药用部位生物量。大量研究表明,逆境能促进植物次生代谢产物的增加[37-39],因此未来需测定药用成分含量,以分析不同光环境对蜘蛛抱蛋生长和药用成分含量的影响,为探索林下仿野生种植提供理论指导。
