地下水埋深和季节性干旱对古尔班通古特沙漠南缘梭梭生理和生长的影响
2022-11-26刘深思徐贵青米晓军陈图强
刘深思,徐贵青,*,米晓军,陈图强,李 彦
1 中国科学院新疆生态与地理研究所,荒漠与绿洲生态国家重点实验室,乌鲁木齐 830011
2 中国科学院阜康荒漠生态国家野外科学观测研究站,阜康 831505
3 中国科学院大学,北京 100049
4 乌鲁木齐城北再生水有限公司,乌鲁木齐 830013
干旱荒漠区因降水稀少、蒸发强烈、缺少地表径流,荒漠植被部分或全部依赖地下水生存。地下水是决定荒漠区植被生长、分布和种群演替的主导因素之一[1],而且在干旱区生态水文过程和循环中发挥着不可替代的作用[2]。在全球变化的背景下,极端干旱事件频发与气候变暖加剧了荒漠生态系统的干旱程度[3]。近年来,因工业和农业的生产需求,地下水被大量开采,导致地下水储量减少、地下水位下降和地下水质量变差。与降水相比,地下水是植物群落持续且稳定的水源。然而,地下水埋深的变化会改变生态系统的水分平衡,引起植被群落组成、水分利用策略和功能性状进行调整[4—5]。研究地下水埋深增加引起的地下水水文干旱与植物之间的相互作用对干旱区地下水的可持续利用和生态系统的健康发展至关重要[6]。
水力性状的调节是植物抵御干旱胁迫的重要方面。植物的水分调节包括叶水势、水力导度、水分利用效率、水力安全阈值等[7]。地下水埋深增加引起的水文干旱,造成水分有效性降低,促使植物进行水分调整。但不同的植物对水文干旱的敏感性和适应性具有较大的差异。例如,胡杨(Populuseuphratica)通过提高水分利用效率[8]和叶片最大导水率、降低饱和膨压和膨压消失点渗透势以适应水文干旱[9];古丁氏柳(Salixgooddingii)以较低的黎明前叶水势应对水文干旱,而柽柳(Tamarixchinensis)的水分有效性则没有改变[10]。尽管已有研究揭示梭梭(Haloxylonammodendron)的同化枝水势与地下水埋深呈显著的负相关[11—12],但荒漠植物水力性状在生长季的不同阶段会表现出不同的变化趋势[13],因此深入研究梭梭的水分调节对策仍具有重要意义。
形态调节是植物在干旱胁迫下维持生存的重要生态策略。在器官水平上,根、茎、叶的表型可塑性是植物适应干旱的重要组成部分。在荒漠生态系统中,地下水埋深增加引起的水文干旱是导致植物发生形态调整的关键环境因子。骆驼刺(Alhagisparsifoli)幼苗增加根系生长速率[14]、胡杨降低展叶效率[15]、柽柳增加冠层枯死率[10]的策略来响应地下水埋深的增加。部分旱生植物的叶片为了减少蒸腾面积而退化为膜质鳞片,由同化枝代替叶进行同化作用[16]。此外,安富博等[17]研究表明,地下水埋深增加造成民勤绿洲人工梭梭林当年新枝生长量急剧减少。然而,在极端干旱的条件下,关于荒漠植物叶片生长速率与地下水埋深关系的研究还少有报道。
积累渗透调节物质是植物抗旱的一种重要生理适应机制[18]。有机溶质和无机离子是植物体内两种重要的渗透调节物质,它包括可溶性糖、脯氨酸、钠离子、氯离子等一系列渗透活性分子/离子[19]。发挥渗透调节作用的物质因物种和环境胁迫而不同。梭梭通过积累钠离子以抵御干旱胁迫;白梭梭(Haloxylonpersicum)主要依靠积累可溶性糖来抵抗盐碱的危害[20];胡杨通过积累可溶性糖、蔗糖、脯氨酸、甜菜碱和钾离子等多种渗透调节物质来适应地下水埋深增加引起的水文干旱[21]。从时间尺度上来看,植物体内渗透调节物质不仅具有日变化[22],而且因物候[23]、树龄[24]发生改变。然而,荒漠植物体内渗透调节物质和非结构性碳水化合物对地下水埋深增加的响应特征,尚不清楚。
梭梭,属藜科梭梭属灌木或小乔木,是古尔班通古特沙漠南缘荒漠灌丛植物的建群种之一。它具有耐贫瘠、耐干旱、耐盐碱、抗风沙等优良特性,是荒漠地区防风固沙、保护草场、改善沙区气候的重要植物,在维护区域生态系统结构和功能方面具有很高的价值[11]。早期研究认为梭梭应对水分胁迫(降水及其转化而来的表层土壤水引起的水分亏缺)的一个重要策略是稳定的气孔开度与有效的形态调节[25]。在水分条件较好时,具有高的光合速率、气孔导度和蒸腾速率;水分匮乏时则选择落叶并进入休眠,生长减缓或停滞[26],并认为梭梭主要依赖降水,是雨养旱生植物。但进一步研究发现,在干旱少雨甚至无雨的生长季后期,地下水是主要水源甚至唯一水源[27]。此外梭梭水分吸收策略存在年际尺度的调整,相对于干旱年份,梭梭增加了湿润年份对表层土壤水的利用比例,但表层土壤水对植株水分来源的总体贡献度仍不及深层土壤水和地下水[28]。由此可见梭梭主要依赖地下水,可以称之为兼性潜水植物。当前,地下水埋深增加对梭梭生存造成了严重破坏,而且发现梭梭在不同区域存在大面积的退化死亡现象[29]。本文沿地下水位梯度对古尔班通古特沙漠梭梭的生理生态特征进行研究,旨在阐释梭梭的水分调节对策、形态结构调整和渗透调节过程,这对于充分认识荒漠梭梭林的抗旱生存维持机制具有重要意义,同时有助于荒漠梭梭林的管理和保育。
1 材料与方法
1.1 研究区概况
研究区域位于古尔班通古特沙漠南缘,中国科学院阜康荒漠生态系统国家站沙漠试验区。该区域属典型的温带大陆性沙漠气候,昼夜温差大,夏季炎热,冬季寒冷,平均积雪深度为20—30 cm,年均积雪覆盖时间为100—150 d[30]。年平均气温为6.6℃,年平均降水量约为160 mm,年平均蒸发量约为2000 mm[25]。地势呈南高北低,海拔370—500 m,地貌类型以固定与半固定沙丘、沙垄为主。沙丘顶部是以白梭梭植物群落为代表的荒漠植物群落,盖度为15%—30%;垄间则以梭梭群落为主,盖度为20%—50%[31]。物种丰富度相对较高,群落常伴生有沙漠绢蒿(Seriphidiumsantolinum)、尖喙牻牛儿苗(Erodiumoxyrrhynchum)、条叶庭荠(Alyssumlinifolim)等草本植物。
1.2 实验样地设置
在北沙窝野外试验区,选择5个实验样点组成地下水埋深变化梯度,实验样点分别记为S1、S2、S3、S4和S5(表1)。根据土壤水分状况的季节性差异[11],将所有的实验测量分为两个阶段:生长季前期(4—6月)和生长季后期(7—9月)。生长季前期土壤受到冬季冰雪融水和春季降水的补给;生长季后期气温升高,蒸发加剧,土壤水分亏缺得不到及时补充[32],浅层土壤含水量(0—100 cm)偏低(图1)。在每个样点选择5棵长势良好、株高冠幅相近的成年梭梭测量其生理指标。利用GPS测量仪(ZL180,卓林科技,安徽合肥)测量样点的地理位置和海拔,利用激光测距仪(DL4168,得力,浙江)测量样点地下水埋深信息。
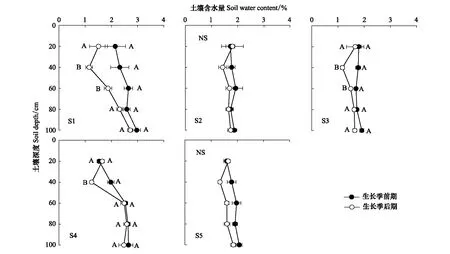
图1 生长季前期和后期土壤含水量的变化
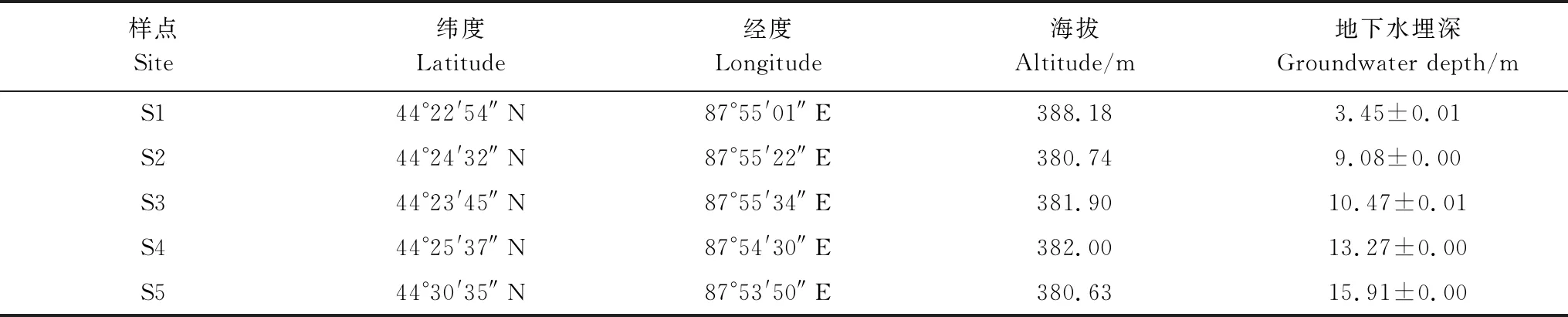
表1 样点地理位置、海拔与地下水埋深
1.3 试验方法
1.3.1 土壤含水量
利用土钻在植物采样点附近采集浅层土壤样品,深度为1 m,土层深度分别为0—20 cm,20—40 cm,40—60 cm,60—80 cm,80—100 cm。采集的土壤样品装入铝盒,用烘干法测量土壤含水量。每个实验样点3个重复。
1.3.2 同化枝水势
利用植物水势压力室(Model 3500, PMS Instrument Company, Albany, USA) 测量梭梭同化枝的黎明前和正午水势。黎明前水势(Ψpd)在日出前30 min测定,正午水势(Ψm)在13:00—14:00测定,每个实验样点8个重复。
1.3.3 同化枝含水量
同化枝含水量采用烘干称重法测定。清晨剪取同化枝样品,将其迅速放入随身携带的冰盒中,带回实验室称取鲜重,然后放入烘箱在75℃下干燥48 h,称其干重,3个重复。同化枝含水量(ABWC)的计算公式为:(同化枝鲜重-同化枝干重)/同化枝鲜重×100%。
1.3.4 同化枝生长速率
梭梭同化枝生长速率在4—8月进行测定。每棵梭梭标记两个同化枝。以白板为底,白板上带有特定面积的黑块,使用相机(EOS550D, Canon Inc., Japan)定期拍摄照片,拍照间隔为10 d或11 d。利用图像处理软件(Adobe Photoshop CS6,USA)去除照片中除实验对象之外的其它枝条。使用CI- 400 CIAS软件(Computer Imaging Analysis Software)批量计算出黑块与同化枝的面积,通过黑块实际面积与计算面积的比例关系计算出同化枝的实际面积[25]。同化枝生长速率(ABGR)计算公式,如下:
式中,St为t时刻的同化枝面积,St+1为t+1时刻的同化枝面积,Δt为两次拍照的时间间隔。
1.3.5 胡伯尔值
在每棵梭梭上剪取一个南面生长的三级枝条。使用扫描仪(Epson Perfection 2400 Photo, Seiko Epson, Japan)对每个枝条上所有的同化枝面积进行扫描。用品红溶液对枝条横截面染色,用游标卡尺测量枝条的边材和心材直径,计算枝条横截面的导水面积。胡伯尔值,即为枝条导水面积与叶片总面积的比值。
1.3.6 渗透调节物质和非结构性碳水化合物
采集当年新生的同化枝,带回实验室。将样品置于65℃烘箱烘干48h后,使用混合震荡型研磨仪研磨成粉。测定指标包括钠离子(Na+)、氯离子(Cl-)、脯氨酸(Pro)、可溶性糖(SS)和淀粉。Na+采用火焰光度计(FP640,上海精科)测定;Cl-采用以明胶-乙醇水溶液为稳定剂的氯化银沉淀法测定[33];脯氨酸采用茚三酮比色法测定[34];可溶性糖和淀粉采用苯酚硫酸法测定[35],非结构性碳水化合物为可溶性糖和淀粉之和。每个指标的测定均为3个重复。
1.4 数据分析
利用SPSS 21.0软件对数据进行统计分析, 利用单因素方差分析(One-way ANOVA)中的Duncan检验分析不同地下水埋深生理生态参数的显著性差异,利用双因素方差分析(Two-way ANOVA)确定地下水埋深、生长阶段以及地下水埋深×生长阶段的交互作用对梭梭生理生态参数的影响;利用独立样本T检验分析同一地下水埋深不同生长阶段生理生态参数的显著性差异;采用Pearson相关分析探讨地下水埋深对梭梭同化枝生长速率的影响。数据分析之前对所有数据进行正态性和方差齐性检验。使用SigmaPlot 12.5软件对数据进行绘图。
2 结果
2.1 不同地下水埋深和生长期梭梭生理生化特征的变化
不同生长阶段梭梭沿地下水埋深增加的水分关系如图2所示。地下水埋深和生长阶段对Ψpd和Ψm有显著的影响(P<0.001)。Ψpd和Ψm随地下水埋深的变化与生长阶段有关。生长季前期Ψpd和Ψm显著高于生长季后期(P<0.05)。随着地下水埋深的增加,生长季前期,S1梭梭Ψpd最高,明显高于S2—S4(P<0.05);S1和S3Ψm明显高于其它3个样点(P<0.05);生长季后期,梭梭Ψpd先降后升,Ψm在较浅的地下水埋深明显高于较深的地下水埋深(P<0.05)。地下水埋深对ABWC产生了显著的影响(P<0.001)。生长季前期和后期,梭梭ABWC先升后降。
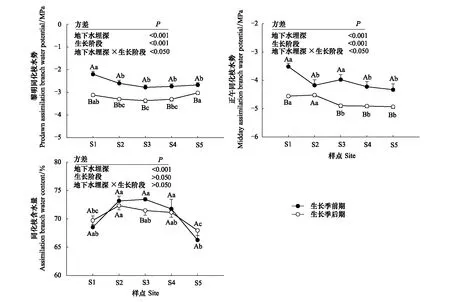
图2 不同地下水埋深下梭梭水分关系的变化
不同生长阶段梭梭同化枝中渗透调节物质和非结构性碳水化合物沿地下水埋深梯度的变化特征如图3所示。地下水埋深对梭梭同化枝中Cl-、SS和碳水化合物(NSC)含量的影响达到极显著水平(P<0.01)。随着地下水埋深的增加,梭梭同化枝中Cl-的含量先升后降;S1—S4同化枝中SS和NSC含量差异不显著, 但在S5 SS和NSC含量显著升高(P<0.05)。生长阶段对梭梭同化枝中Na+和SS含量产生了极显著的影响(P<0.01)。生长季前期梭梭同化枝中Na+含量显著低于生长季后期(P<0.05)。地下水埋深对梭梭同化枝中Pro和淀粉含量的影响与生长阶段有关(P<0.05)。生长季前期,S3梭梭同化枝中脯氨酸(Pro)含量最高,且明显高于其它4个样点(P<0.05); S1—S4同化枝中淀粉含量差异不显著,但在S5淀粉含量明显减少(P<0.05)。生长季后期,S4梭梭同化枝中Pro含量最低,另4个样点Pro含量明显高于S4(P<0.05),且不存在明显差异;同化枝中淀粉含量相对稳定。

图3 不同地下水埋深下梭梭同化枝渗透调节物质和非结构性碳水化合物的变化
2.2 不同地下水埋深和生长期梭梭生长和形态的变化
不同地下水埋深下梭梭的同化枝生长速率如图4所示。地下水埋深对4—5月梭梭ABGR产生了显著的影响,5月地下水埋深较浅的ABGR明显高于地下水埋深较深的地点(P<0.05)。6—8月地下水埋深对ABGR没有显著的影响。不同地下水埋深梭梭胡伯尔值的变化特征如图5所示。地下水埋深、生长阶段和二者的交互作用对梭梭HV的影响均达到极显著水平(P≤0.001)。生长季前期,在较浅的地下水埋深HV差异不显著,在较深的地下水埋深HV显著增大(P<0.05)。生长季后期,在地下水埋深增加的条件下,梭梭HV相对稳定。

图4 不同地下水埋下深梭梭同化枝生长速率

图5 不同地下水埋深下梭梭胡伯尔值的变化
3 讨论
3.1 地下水埋深和季节性干旱对梭梭生理性状的影响
叶(同化枝)水势和含水量不仅能反映植物的水分状态,也是植物进行水分调节的重要表征。冯树林等[36]研究发现,随着干旱胁迫的加剧,荒漠灌木叶水势呈下降趋势。然而,也有学者认为,荒漠灌木的水力行为高度动态,在不同的水分供应状态和温度条件下,其水力调节对策可从部分等水势转变为极端非等水势[13]。本研究中,随着地下水埋深的增加,生长季前期和后期梭梭Ψpd均呈现先降低后升高的变化趋势(图2),表明梭梭Ψpd受到地下水埋深增加引起的水文干旱胁迫,但地下水有效性降低促使梭梭水分利用效率提高、气孔导度降低以及蒸腾速率下降[11—12],这在一定程度上补偿了地下水埋深持续增加对其同化枝水势的负面影响。与较浅的地下水埋深相比,较深的地下水埋深下Ψm明显降低(图2),这与前人[11]研究的结果一致。此外,与生长季前期相比,除S2的Ψm外,生长季后期Ψpd和Ψm均显著降低(图2),这表明梭梭经历季节性大气干旱。梭梭是荒漠环境中的旱生肉质植物,同化枝角质层厚,储水组织发育良好[37]。因此,尽管地下水埋深在15 m以上,生长季前期和后期梭梭ABWC仍高于65%以上(图2),这表明高ABWC是梭梭在干旱生境中长期生存的适应性特征。
渗透调节是植物在细胞水平上减轻干旱胁迫的一种适应策略。渗透调节物质以不同的方式保护植物免受胁迫,如提高细胞的渗透调节能力和保护膜的稳定性。植物在生长过程中对氯的需求量十分少,一般在0.03%—2.0%之间。梭梭同化枝中Cl-含量低,且随地下水埋深的增加先升后降(图3),这表明梭梭同化枝中Cl-的渗透调节能力有限[38],当水分有效性降低到一定程度时,植物体内Cl-向地上部分的迁移会减弱[39]。在模拟干旱的温室条件[40]和极端干旱的自然环境中[20], Na+对梭梭渗透势调节的贡献均高达45%—50%。本文中梭梭同化枝中Na+的含量没有受到地下水埋深的限制,而是因生长阶段而异。在极端干旱的荒漠环境中,Na+是梭梭长期适应干旱的重要渗透调节物质,其含量随干旱胁迫时间的延长而累积。梭梭同化枝中Pro含量在多个样点差异均不显著(图3),说明梭梭同化枝中Pro对渗透调节的贡献较小且有限。吕昕培等[40]在研究梭梭幼苗应对干旱胁迫的渗透调节作用时,也发现Pro对渗透调节的贡献水平十分有限。
有研究发现,在渗透胁迫下,梭梭幼苗同化枝中的SS对渗透调节的贡献较小[40]。但王国华等[38]研究表明,SS是种植年限为5—20年的梭梭抵御干旱胁迫的重要渗透调节物质。树龄的增长促使梭梭体内非结构性碳水化合物(NSC)含量成倍提高[24]。在本试验中,生长季前期与后期梭梭同化枝中SS含量较高,且在S5显著升高(图3)。由此表明,SS是成年梭梭抵御水文干旱的渗透调节物质之一。淀粉作为植物的储存物质,通常在植物遭受干旱时部分转化为SS以供植物应对环境胁迫[41]。这与本研究结果一致,生长季前期梭梭同化枝中淀粉含量在S1-S4相对稳定,但在S5显著下降(图3)。而且,与生长季前期相比,生长季后期同化枝中淀粉含量出现明显的损耗(图3)。这表明梭梭可以将同化枝中的淀粉转化为SS以抵御地下水埋深增加的水文干旱和季节性大气干旱。干旱会改变树木的碳平衡,NSC是在干旱胁迫下衡量碳平衡的重要性状[42]。有研究表明,干旱胁迫下植物NSC含量减少[43],但NSC含量累积和维持[44]的结论也有报道。本文中,地下水埋深持续增加引起的水文干旱没有导致梭梭NSC含量耗尽,反而显著升高(图3)。这意味着梭梭没有受到碳限制,在干旱胁迫下维持较高的NSC含量,这对其水力功能恢复可能十分重要[45]。
3.2 地下水埋深和季节性干旱对梭梭生长和形态的影响
植物生长对水分的变化非常敏感,尤其是荒漠植被。地下水埋深增加引起的水文干旱会改变植物的表型可塑性[46]。有研究表明,干旱胁迫会造成荒漠植物生长缓慢、生物量减少、总叶面积和比叶面积减小[47—48]。早期研究发现,夏季降水的减少会限制梭梭的生长[25]。本研究中,相关性分析表明,4—5月梭梭ABGR与地下水埋深呈显著负相关(P<0.05)。蒋菊芳[49]、陈玲[50]等研究表明,梭梭萌芽期为4月12日,展叶期为4月30日,新枝形成期为5月22日。这意味着在新枝形成期,地下水埋深增加会导致ABGR显著降低。由此可见,梭梭可以通过改变同化枝生长速率来响应地下水水文干旱。研究结果与彭守兰等[14]的结论一致,他们发现疏叶骆驼刺随着潜水埋深的增加,生长缓慢且生物量累积减少,并且认为这种形态的可塑性是荒漠灌木长期适应干旱贫瘠环境的生态策略。
叶片的形态和结构与植物的抗旱机制密切相关,胡伯尔值(HV)被认为是评价植物抗旱性的重要功能性状[51]。在干旱胁迫下,植物通常具有较高的HV以维持叶片水分关系的稳定[52]。在阿塔卡马沙漠牧豆树(Prosopistamarugo)[53]和塔克拉玛干沙漠胡杨[15]的研究中表明,地下水埋深增加会促使HV显著增大。这与本研究结果一致,生长季前期梭梭HV随地下水埋深的增加呈现增大的趋势(图5),这增强了单位叶面积的供水能力;生长季后期梭梭采取有效的形态调整,维持较高的HV值以保证水分运输的有效性(图5)。
4 结论
地下水埋深、生长阶段及其交互作用对古尔班通古特沙漠梭梭的生理生化和生长与形态产生了不同的影响。主要结论为:(1)在生理调节上,梭梭通过先降低后升高Ψpd和积累NSC响应地下水埋深的增加。(2)在生长和形态调整上,梭梭在生长季前期采取降低新枝形成期的ABGR和增大HV的策略响应地下水埋深的增加。(3)与生长季前期相比,梭梭采取降低Ψpd、维持较高的HV、积累Na+和消耗淀粉的策略抵御季节性干旱。(4)在地下水埋深和生长阶段的交互作用下,降低Ψm、维持较高的ABWC和积累SS是梭梭适应大气和地下水水文干旱的结果。由此可见,梭梭生理和形态性状对地下水水文干旱和季节性大气干旱及其交互作用间的响应,既存在一致性,又存在差异性。
