锰胁迫对大叶胡枝子种子萌发和幼苗生理生化特征的影响
2022-11-22胡佳瑶王悟敏匡雪韶刘文胜
胡佳瑶,王悟敏,匡雪韶,刘文胜
(中南林业科技大学 生命科学与技术学院,湖南 长沙 410004)
锰(manganese,简称Mn)是植物正常生长发育必不可少的微量元素,直接参与植物光合作用,对叶绿体膜正常结构的维持具有重要意义[1].过量锰则会对植物产生毒害作用,甚至能通过食物链危害人类健康[2].锰矿开采带来经济效益的同时,也造成了大面积的锰污染,严重影响了人们正常的生产生活[3].因而,治理锰矿区污染已刻不容缓.植物修复技术有着成本低、适应范围广、环境扰动小以及治理效果持久等优点而受到国内外的广泛关注,筛选耐锰性较强的植物是利用该技术的关键之一[4].
种子萌发和幼苗生长是植物生长发育过程中最脆弱的时期,该阶段最能够体现植物对逆境的耐受性.植物在受到重金属胁迫时,体内的渗透调节物质(可溶性糖、可溶性蛋白和游离脯氨酸)和抗氧化酶活性(过氧化物酶(peroxidase,简称POD)、超氧化物歧化酶(superoxide dismutase,简称SOD)和过氧化氢酶(catalase,简称CAT))等都会发生一系列变化来缓解胁迫所带来的毒害,而丙二醛(malondialdehyde,简称MDA)是植物细胞膜脂过氧化的最终产物,其含量的高低可用来反映植物被毒害的程度[5].因而,胁迫条件下,植物种子萌发、幼苗生长以及生理生化特征的变化是植物适应环境的重要机制.例如,王波等[6]研究发现,南荻(Triarrhenalutarioriparia)的发芽率、发芽势、发芽指数和活力指数均随铅、镉浓度的增加而下降.曹婧等[7]研究发现,紫花苜蓿(Medicagosativa)的株高、生物量和抗氧化酶活性均随着锰胁迫的增加而先升后降,MDA含量则持续上升.
大叶胡枝子(Lespedezadavidii)为豆科(Fabaceae)胡枝子属(Lespedeza)的一种直立灌木,广泛分布于我国南方地区.该植物抗逆性强,可在多种生境中生长,能生物固氮,是逆境生物治理的重要选择[8-9].笔者对湖南湘潭锰矿废弃地调查显示,大叶胡枝子在该区域生长良好,能够正常开花结实,这说明它具有较强的锰污染耐受能力.因而,该植物是锰尾矿区一种可选择的重要修复植物.目前,大叶胡枝子的相关研究主要集中于其药用价值方面[10-11],关于锰胁迫下该植物适应性机制尚未系统研究,这限制了该植物的进一步应用.
笔者以大叶胡枝子为对象,在不同浓度锰胁迫下开展种子萌发及幼苗生长实验,研究该植物种子的萌发特性以及不同时期幼苗的生理生化特征,探究其在锰胁迫下的生理耐受机制,以期为锰污染区的植物修复提供理论依据.
1 材料与方法
1.1 实验材料
大叶胡枝子种子于2019年11月采于湖南湘潭锰矿区(28°03′E,112°55′N).现场调查发现,该植物可在矿区土壤锰含量达52 319.25 mg·kg-1的土壤中自然生长,其高度达1.7 m,且能够正常开花结实.选择其中50株长势良好的大叶胡枝子,采集其果实并装入布袋带回实验室,于室温保存备用,称量结果显示该植物种子千粒重为9.39 g.
1.2 实验方法
1.2.1 种子萌发实验
根据预实验结果及马彦军等[12]的研究,为提高种子萌发率,挑选大小一致、颗粒饱满的种子采用浓硫酸浸泡法处理种子,即用浓硫酸浸泡种子10 min,破除其硬实性.根据预实验结果并参考潘高等[13]的方法,设置锰胁迫浓度为5 000,15 000 μmol·L-1.由于锰为植物生长的必需元素,其5 μmol·L-1是植物正常生长需要的微量元素水平[14],参照梁文斌、豆长明、吴惠芳等[15-17]的研究,设置锰浓度5 μmol·L-1为对照.利用MnCl2·4H2O设置5(对照)、5 000和15 000 μmol·L-13个锰处理开展相关实验.萌发实验在温度为(26±0.5)℃,12 h光照/12 h黑暗的人工气候培养箱中进行.萌发时,种子放入装有两层滤纸、直径为90 mm的培养皿中,每个培养皿放置25粒种子,每个浓度设有5个重复,共计放置15个培养皿375粒种子.萌发实验开始后,每24 h换1次锰溶液和滤纸,以保持胁迫浓度恒定.每天定时记录种子萌发情况.胚根突破种皮1/2视为种子萌发,连续3 d无新种子萌发即为结束.实验结束后测定大叶胡枝子幼苗的生物量,计算种子发芽率、发芽势、发芽指数、活力指数与根冠比.
1.2.2 幼苗生长实验
将处理后的种子放入装有150 mL基质(V(沙)∶V(珍珠岩)=1∶1)的塑料杯中,然后放入人工培养箱中(与萌发实验相同条件),每天适量浇水,当子叶展开后用1/2 Hoagland营养液培养幼苗.待幼苗长出5~6片真叶时,加入与萌发试验相同的锰浓度处理,继续培养,期间每3 d浇1次Hoagland营养液,分别在胁迫第7,15,30天时对幼苗叶片进行生理生化指标测定.
1.3 测定项目和方法
1.3.1 种子萌发指标测定
参照文献[13]测定:发芽率=10 d正常发芽种子数/总种子数×100%,发芽势=4 d正常发芽种子数/总种子数×100%,发芽指数(GI)=∑(Gt/Dt),活力指数(VI)=GI×h,其中:Gt为在t日的发芽种子数,Dt为发芽天数,h为幼苗高度.
1.3.2 幼苗生物量指标测定
萌发结束后,每个处理随机采集10棵幼苗,用游标卡尺(0.1 mm)测量其芽长和根长.用解剖刀将其分为地上和地下两部分,称量鲜重后装入信封,放进烘箱中烘至恒重(先105 ℃杀青5 min,然后70 ℃继续烘至恒重).根冠比=地下部分干重/地上部分干重[18].
1.3.3 生理生化指标测定
胁迫后每个处理取鲜样0.1 g,每个处理3个重复,生理生化指标用以下方法测定:丙酮浸提法测定大叶胡枝子叶片中光合色素含量;氮蓝四唑法测定叶片中SOD活性;愈创木酚法测定叶片中POD活性(以每分钟OD值变化0.01为1个酶活性单位);紫外吸收法测定叶片中CAT活性(以每分钟OD值变化0.01为1个酶活性单位);蒽酮比色法测定叶片中可溶性糖含量;考马斯亮蓝法测定叶片中可溶性蛋白含量;酸性茚三酮显色法测定叶片中游离脯氨酸含量;硫代巴比妥酸法测定叶片中MDA含量[19].
1.4 数据处理
采用SPSS 22.0进行LSD(p<0.05)多重比较分析.利用Sigma Plot 12.5进行绘图.
2 结果与分析
2.1 萌发及幼苗生长
如表1所示,大叶胡枝子种子的发芽势、发芽指数及活力指数都随锰浓度的升高而显著降低(p<0.05),锰胁迫为15 000 μmol·L-1时,其发芽势低于26.00%,相对于对照减少了34.69%;发芽指数低于21.00,相对于对照减少了17.77%;活力指数低于1.00,相对于对照减少了25.20%.锰胁迫对该植物的种子发芽率无明显影响(p>0.05),均在98%左右.
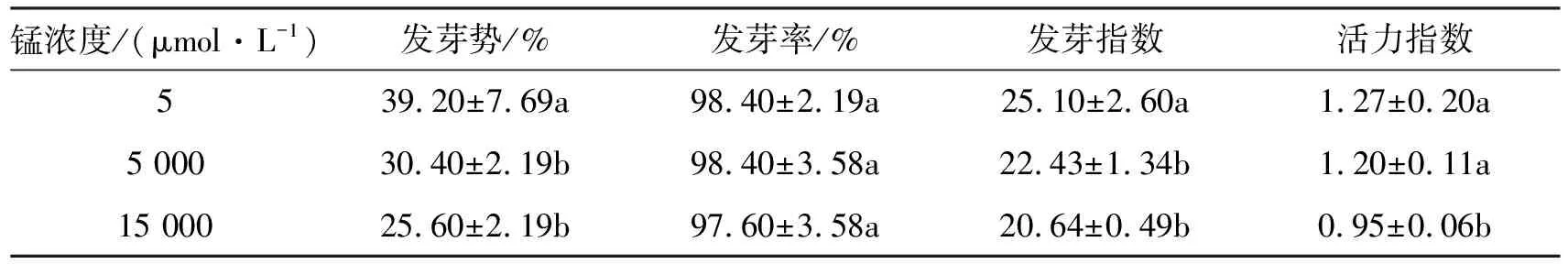
表1 锰胁迫对大叶胡枝子种子萌发的影响
如表2所示,大叶胡枝子幼苗的根长、芽干重、根干重、总干重和根冠比均随锰浓度的升高而先增加后减少,5 000 μmol·L-1处理的锰对该植物幼苗的根长、干重和根冠比有明显的促进作用,其根干重和对照相比差异显著(p<0.05),芽长差异则不显著(p>0.05).锰浓度为15 000 μmol·L-1时,该植物幼苗的芽长、根长和干重相比对照均显著下降(p<0.05),分别减少了24.90%,56.19%,32.52%.
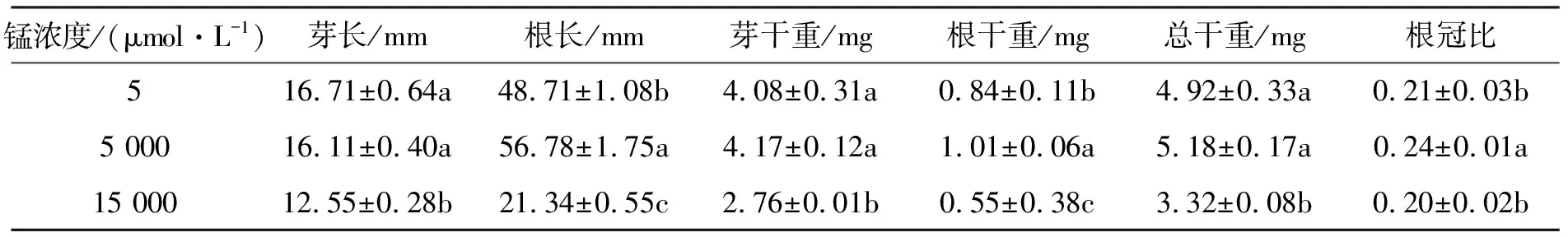
表2 锰胁迫对大叶胡枝子幼苗生长的影响
2.2 光合色素含量
如表3所示,锰处理的不同时间和不同锰浓度对叶绿素a和类胡萝卜素含量有显著影响(p<0.001),且他们的交互作用显著(p<0.001).不同锰浓度对叶绿素b含量有显著影响(p<0.001),不同时间则对其影响不显著(p>0.05),他们的交互作用显著(p<0.05).锰胁迫7 d时,大叶胡枝子幼苗叶片内叶绿素a含量随锰浓度增加而显著下降(p<0.05),分别比对照减少了13.46%和10.20%,叶绿素b含量先减少后增加,类胡萝卜素含量先增加后减少;锰胁迫15 d时,叶绿素a、叶绿素b含量随锰浓度增加而显著下降(p<0.05),类胡萝卜素则先上升后下降;锰胁迫30 d时,叶绿素a、叶绿素b和类胡萝卜素随锰浓度增加而显著下降(p<0.05),且均在15 000 μmol·L-1锰处理达到最低值,比对照分别减少了25.13%,15.38%,21.21%.
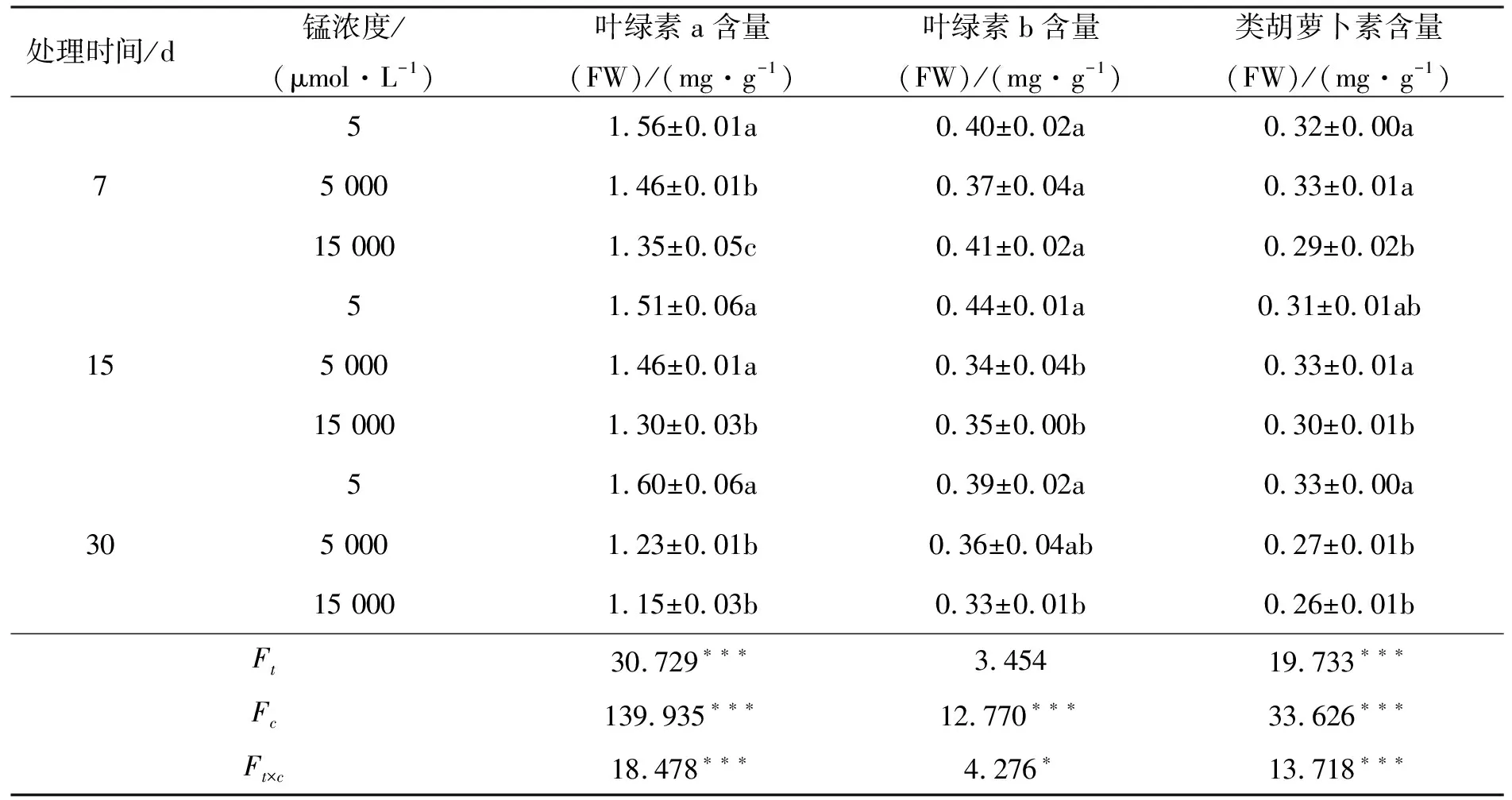
表3 锰胁迫对大叶胡枝子幼苗光合色素含量的影响
2.3 抗氧化酶活性
测定锰处理的不同时间和不同锰浓度时大叶胡枝子幼苗叶片中相应抗氧化酶的活性,结果如图1所示.

图柱上不同小写字母表示同一时间内不同浓度间的比较差异显著(p<0.05).Ft代表处理时间的F值;Fc代表锰浓度的F值;Ft×c代表处理时间与锰浓度交互作用的F值;*代表0.05水平下影响显著,***代表0.001水平下影响极显著.
由图1可知,不同处理时间、不同锰浓度对SOD,POD,CAT活性有显著影响(p<0.001),且他们的交互作用显著(p<0.001).
大叶胡枝子幼苗叶片中SOD活性随时间的推移而升高,且在3个时间段均随锰浓度的增大而显著上升(p<0.05),在15 000 μmol·L-1锰处理15 d时达到最大值.30 d时,5 000 μmol·L-1和15 000 μmol·L-1锰处理的SOD活性相比于15 d的有所减少,分别减少了12.32%和16.99%.
POD活性随时间的推移而逐渐降低,处理7 d时随锰浓度的增加而显著上升(p<0.05),处理15 d和30 d时则随锰浓度的增大而显著降低(p<0.05),在30 d时15 000 μmol·L-1锰处理处达到最低值.30 d时,5 000 μmol·L-1和15 000 μmol·L-1锰处理的POD活性相比于对照有所减少,分别减少了21.52%和49.32%.
CAT活性随时间的推移而先上升后下降,在处理7 d和15 d时随锰浓度的增大而显著上升(p<0.05);在处理30 d时则随锰浓度的增大而先上升后下降,15 000 μmol·L-1锰处理30 d时达到最低值;5 000 μmol·L-1和15 000 μmol·L-1锰处理30 d时的CAT活性相比于15 d时的有所减少,分别减少了9.46%和57.10%.
2.4 渗透调节物质含量
测定处理不同时间和不同锰浓度时大叶胡枝子幼苗叶片中渗透调节物质的含量,结果如图2所示.

图柱上不同小写字母表示同一时间内不同浓度间的比较差异显著(p<0.05).Ft代表处理时间的F值;Fc代表锰浓度的F值;Ft×c代表处理时间与锰浓度交互作用的F值;*代表0.05水平下影响显著,***代表0.001水平下影响极显著.
由图2可知,不同时间和不同锰浓度处理对可溶性糖、可溶性蛋白和游离脯氨酸含量有显著影响(p<0.001),且他们的交互作用显著(p<0.001).大叶胡枝子幼苗叶片可溶性糖随锰胁迫时间的推移而不断升高,随锰浓度的升高而增加,且在处理15 d和30 d时差异显著(p<0.05).在15 000 μmol·L-1锰处理30 d时可溶性糖含量达到了最大值,相比于对照增加了127.00%.
可溶性蛋白含量随锰处理时间的推移而逐渐升高,随锰浓度的升高而增加,且在处理7 d和15 d时差异显著(p<0.05).在15 000 μmol·L-1锰处理30 d时可溶性蛋白含量达到最大值,相比于对照增加了310.34%.
游离脯氨酸含量随锰处理时间的推移而先升高后降低,随锰浓度的升高而增加,且在处理7 d和15 d时差异显著(p<0.05).15 000 μmol·L-1锰处理幼苗叶片的游离脯氨酸含量在处理7 d时达到最大值,相比于对照增加了325.62%,处理30 d时达到最低值,相比于处理7 d减少了43.29%.
2.5 MDA含量
测定处理不同时间和不同锰浓度时大叶胡枝子幼苗叶片中MDA的含量,结果如图3所示.

图柱上不同小写字母表示同一时间内不同浓度间的比较差异显著(p<0.05).Ft代表处理时间的F值;Fc代表锰浓度的F值;Ft×c代表处理时间与锰浓度交互作用的F值;*代表0.05水平下影响显著,***代表0.001水平下影响极显著.
如图3所示,不同处理时间和锰浓度对MDA含量有显著影响(p<0.001),且他们的交互作用显著(p<0.001).大叶胡枝子幼苗叶片中MDA含量随锰处理时间的推移而逐渐升高,随锰浓度的升高而增加,且在处理15 d和30 d时差异显著(p<0.05).30 d时15 000 μmol·L-1锰处理的MDA含量达到了最大值,为61 nmol·g-1,比对照增加了226.20%.
3 讨 论
3.1 种子萌发及幼苗生长
种子萌发和幼苗生长是植物对逆境最敏感的阶段,胁迫条件下的发芽和生长能力可体现该植物的耐受性.该研究中,锰胁迫对大叶胡枝子种子发芽率无明显影响,与张龙冲等[20]对铬胁迫下虎耳草(Saxifragastolonifera)的研究结果一致.说明大叶胡枝子对锰有一定的耐受能力,能在锰含量相对较高的环境下萌发.其原因可能是大叶胡枝子的种子较大,相对于小种子其耐锰能力和发芽能力更强[21].随锰浓度的升高,该植物种子的发芽势、发芽指数和活力指数均降低,与邹璐璐等[22]对镉胁迫下河岸带草本植物的研究结果一致.说明高浓度的锰胁迫对大叶胡枝子种子萌发速度有影响.
研究显示,大叶胡枝子幼苗的生物量随锰浓度的升高而先增加后减少,与陈丽丽等[23]在锌胁迫下稗草(Echinochloacrusgali)幼苗的研究结果一致.说明低浓度的锰能使植物的生物量增加.其原因可能是低浓度的锰促进了植物光合作用,使营养物质生产加快,植物生物量增加.随着锰浓度的增加,该植物幼苗的根受到的抑制作用比芽大.这与肖厚军等[24]对油菜(Brassicarapavar.oleifera)的研究结果一致.其原因可能是根作为植物吸收重金属的主要器官,它与锰溶液直接接触,受到的锰毒害程度大于芽.
3.2 幼苗光合色素含量
叶绿素含量是衡量植物光合作用强弱的重要生理参数,能够体现植物对逆境的耐受性[25].在该研究中,随着锰浓度的增加,大叶胡枝子幼苗叶片内叶绿素a、叶绿素b和类胡萝卜素含量不断降低,且在锰最高浓度15 000 μmol·L-1处理时达到最低值,与张宝成等[26]对锰胁迫下小飞蓬(Erigeroncanadensis)和汪结明等[27]对3种蕨类的研究结果一致,说明锰浓度过高时,超出了植物所能耐受的范围,植物细胞中的叶绿体则会受到毒害,光合能力减弱.其原因是锰是组成叶绿体的必需元素,直接参与植物的光合放氧过程,当锰浓度较高时,植物受到了毒害,抑制了叶绿素的合成和相关酶活性,叶绿素分解的速度加快,叶绿素含量降低.
3.3 抗氧化酶活性
SOD,POD和CAT共同构成了植物体内的抗氧化酶系统,具有清除植物体内活性氧、减轻逆境毒害的重要作用.SOD是植物体内重要的抗氧化酶,SOD活性越高表明植物的抗逆性越强.该研究中,大叶胡枝子幼苗叶片中SOD活性随锰浓度的增高而上升,与Li等[28]对镉胁迫下苇状羊茅(Festucaarundinacea)和李强等[29]对锰胁迫下三色堇(Violatricolor)的研究结果一致.其原因可能是随着锰浓度的升高,植物为应对高锰毒害,SOD活性不断增强以减缓锰胁迫导致的氧化伤害;但随着时间的推移,高浓度的锰胁迫超出了植物所能承受范围,植物内环境紊乱,SOD最适环境遭到破坏,其活性降低.
POD活性随着时间的推移而逐渐降低,在处理7 d时随锰浓度的增大显著上升,在处理15 d和处理30 d时则随锰浓度的增大而降低.这与徐玲玲等[30]在镉胁迫下3种草本植物研究结果一致.其原因可能是在锰含量较高的情况下,随着胁迫时间的增加,植物产生活性氧的速度加快,从而导致植物细胞受到毒害程度加大,POD活性降低.CAT活性随时间的推移先上升后下降,在处理7 d和15 d时随锰浓度的增加而显著上升,在处理30 d时则随锰浓度的增加先上升后下降.这与石贵玉等[31]对镉胁迫下红树(Rhizophoraapiculata)幼苗的研究结果一致.其原因可能是在低浓度锰胁迫下,植物体内活性氧增加,细胞的防御反应开始启动,激活了CAT活性,从而减轻活性氧对植物造成的伤害.随着胁迫时间的增加,高浓度锰所带来的毒害超出了植物所能承受范围,CAT适应环境遭到破坏,导致其活性降低.
3.4 渗透调节物质含量
植物受到胁迫时会产生并积累可溶性糖、可溶性蛋白和游离脯氨酸等渗透调节物质,它们能保持植物细胞正常的含水率与膨压,从而对抗外界的不利条件;其含量越多表明植物的抗逆性越强[32].该研究中,大叶胡枝子幼苗叶片可溶性糖和可溶性蛋白含量在3个不同时间段都随锰浓度的升高而增加,与Dong等[33]对锰胁迫下蓝莓(Vacciniumuliginosum)和Huang等[34]对锰胁迫下构树(Broussonetiapapyrifera)的研究结果一致,说明植物在面对逆境时能够通过提高体内的可溶性糖和可溶性蛋白含量来缓解逆境所带来的毒害.其原因可能是可溶性糖是合成有机溶质的碳架和能量来源,而可溶性蛋白具较强的亲水性,可提高细胞保水性能,使植物对锰的耐受能力增强.
研究中,游离脯氨酸含量在3个时间段都随锰浓度的升高呈现增加的趋势,在处理30 d时15 000 μmol·L-1锰处理组相对于前2个时间段游离脯氨酸含量减少.这与鲜靖苹[35]对盐及重金属胁迫下龟背竹(Monsteradeliciosa)的研究结果一致,说明大叶胡枝子幼苗能通过增加游离脯氨酸等渗透调节物质的含量来减轻逆境所产生的毒害作用.其原因可能是锰能加快植物体内营养物质的生产,使植物幼苗能正常生长.随着胁迫时间的推移,锰破坏了植物体内的平衡机制,植物受毒害程度增大,游离脯氨酸含量则会相应地减少.
3.5 MDA含量
MDA是植物细胞膜脂过氧化的最终产物,在植物受毒害情况下,植物体中活性氧的产生与清除的平衡会被破坏,自由基会大量积累从而造成细胞膜脂过氧化,MDA含量也会随着增加,因而其含量的高低多少代表着植物受毒害强弱以及对胁迫的耐受能力.笔者的研究结果显示,大叶胡枝子幼苗叶片中MDA含量随锰胁迫时间的推移而逐渐升高,随锰浓度的升高而增加.这与李欣航等[36]对锰胁迫下鸡眼草(Kummerowiastriata)和查应琴等[37]对镉胁迫下鸡冠花(Celosiacristata)的研究结果一致,说明在锰含量较高的情况下,随着胁迫时间的推移,植物幼苗叶片的MDA含量会随之增加.其原因可能是锰含量过高,导致大叶胡枝子幼苗叶片细胞膜脂过氧化,使细胞受到了破坏,从而加大了植物受毒害的程度.
4 结束语
锰胁迫对大叶胡枝子种子发芽率无明显影响,其幼苗生物量则均表现出“低促高抑”现象,且根受到的抑制作用比芽大.随着锰浓度的增加,大叶胡枝子叶片叶绿素a和叶绿素b含量降低;SOD活性随锰浓度的增加和胁迫时间的延长而逐渐升高,POD活性在处理15d和30 d时随锰浓度的增加而逐渐降低,CAT活性在处理7 d和15 d时随锰浓度的增加而升高,处理30 d时则是先上升后下降.可溶性糖和可溶性蛋白含量随锰浓度的升高和胁迫时间的延长而增加,游离脯氨酸含量随锰胁迫时间先升高后降低,随锰浓度的升高而增加.MDA含量随锰胁迫强度增加、时间延长而逐渐升高.大叶胡枝子能在锰含量较高的环境萌发生长,它主要通过调节体内渗透调节物质含量和抗氧化酶活性来减轻外界胁迫所带来的毒害,其耐锰能力较强,可作为矿区污染修复的候选植物.
