唑来膦酸对小鼠膝骨关节炎的影响及机制
2022-11-22刘国强王丽梅马贵英刘霞丁冬丁一
刘国强 王丽梅 马贵英 刘霞 丁冬 丁一
1.宁夏回族自治区人民医院骨科,宁夏 银川 750001;2.青岛滨海学院医学院,山东 青岛 266555;3.山东第一医科大学附属省立医院手足踝科,山东 济南 250021
骨关节炎(osteoarthritis,OA)是一种累及全关节的慢性、进行性、退变性疾病,其病理表现不仅包括关节软骨退变和软骨下骨重塑异常,还涉及关节周围肌肉、韧带、滑膜的退变以及血管、神经的异常长入[1],最终导致关节疼痛和功能障碍,严重影响患者的生活质量。随着全球人口老龄化速度的加快,OA的发病率逐年增高[2]。目前,OA的发病机制尚不完全明确,无法实现完全治愈。手术治疗和围术期并发症会对患者的身心造成不良影响,加重患者的经济负担,庞大的患病群体对社会医保造成巨大的冲击[3]。
越来越多的证据表明,软骨下骨重塑在OA的发病机制中发挥重要作用,但具体的作用机制尚不明确[4]。软骨下异常骨重塑与OA的进程相关。正常情况下,软骨下骨重塑是由成骨细胞介导的骨形成和破骨细胞介导的骨吸收维持的动态平衡过程。OA发生后,这一平衡被打破,破骨细胞介导的骨吸收增强,软骨下骨量减少,软骨下骨生物力学及微环境的改变进一步导致骨硬化发生,骨赘形成,软骨下骨弹性模量的改变使关节软骨承受的应力负荷分布不均,加速其退变[5-6]。我们前期的研究证实,Wnt/β-catenin信号参与OA软骨下骨中骨髓间充质干细胞(bone marrow mesenchymal stem cells,BMSCs)的成骨分化[7]。另有研究显示,早期改善软骨下异常骨重塑可以延缓OA进展[8]。Wnt蛋白包含19种不同的配体,他们参与了胚胎发育、干细胞维持、组织稳态和肿瘤形成,在骨与软骨的形成以及骨稳态的调节中发挥重要作用[9]。Wnt5a能够结合不同的受体,调节细胞骨架变化、细胞极化、迁移和粘附,这与炎性和免疫性疾病的发生相关[10]。来源于造血干细胞的破骨前体细胞在巨噬细胞集落刺激因子(macrophage colony stimulating factor,MCSF)和NF-κB受体激活剂配体(receptor activator of NF-κB ligand,RANKL)的刺激下分化为成熟多核的破骨细胞,通过分泌抗酒石酸酸性磷酸酶(tartrate-resistant acid phosphatase,TRAP)、基质金属蛋白酶-9(matrix metalloproteinase,MMP-9)等分解骨基质,完成骨吸收[11]。在OA早期,破骨细胞介导的骨吸收增强,加速了OA进程,但破骨细胞发生异常活化的机制尚不明确。唑来膦酸(zoledronic acid,ZOL)作为第三代双膦酸盐类药物,通过抑制破骨细胞介导的骨吸收治疗骨质疏松取得良好的临床效果[12],但其抑制破骨细胞的作用机制尚不明确,且ZOL对OA的治疗效果尚无定论。本研究观察ZOL对小鼠膝关节OA的治疗效果,并探究ZOL抑制OA软骨下骨吸收的机制,为实现OA的早期治疗和精准治疗提供依据。
1 材料与方法
1.1 实验材料
健康10周龄雄性C57BL/6J小鼠45只,体质量(23.9±1.2)g,由宁夏医科大学实验动物中心提供,实验动物许可证号:SYXK(宁)2020-0001。ZOL(瑞士,诺华制药),荧光显微镜(德国,LEICA DM3000),恒冷式冰冻切片机(德国,LEICA CM1850),Elisa试剂盒(上海联硕),兔抗小鼠一抗:Wnt5a、RANKL、NF-κB(55184-1-AP,23408-1-AP,10745-1-AP,中国,Proteintech),HRP标记的羊抗兔二抗(BA1056,武汉博士德),FITC标记的羊抗兔二抗(F-2765,上海赛默飞),甲苯胺蓝染色试剂盒(北京索莱宝),HE染色试剂盒(北京索莱宝),TRAP染色试剂盒(武汉赛维尔),蛋白提取和浓度检测试剂盒(南京凯基),全自动冷冻研磨仪(上海净信)。
1.2 方法
1.2.1 动物分组及处理 将45只小鼠随机分为sham+生理盐水(normal saline,NS)组、OA+NS组和OA+ZOL组。于内侧半月板失稳(destabilization of medial meniscus,DMM)术后第4、第8周处死小鼠,观察并探讨ZOL对小鼠膝OA的影响及作用机制,实验设计如图1。具体过程:小鼠适应性饲养1周后,用1%戊巴比妥钠(60 mg/kg)腹腔注射麻醉。OA+NS组和OA+ZOL组应用DMM方法建立小鼠膝OA模型[13],切断右膝内侧半月板胫侧副韧带,翻转内侧半月板至股骨近端。sham+NS组仅暴露内侧半月板胫侧副韧带,不予切除,缝合创面。OA+ZOL组予以ZOL腹腔注射(100 µg/kg),每周2次,连续4周。sham+NS组和OA+NS组给予等量NS。

图1 实验设计示意图
1.2.2 Micro-CT检测 术后4、8周将各组的5只小鼠的膝关节标本置于4%多聚甲醛溶液中固定48 h,采用SKYSCAN1076扫描仪(扫描参数:像素尺寸为5µm,峰值管电压为50 kV,电流为250µA,旋转角度为0.6°)进行三维CT成像。胫骨内侧平台负重区域(前后范围1 mm,生长板下方0.2 mm,高度0.5 mm)被确定为感兴趣区(region of interest,ROI)。计算各组骨体积分数(bone volume fraction,BV/TV)、骨小梁厚度(trabecular thickness,Tb.Th)、骨小梁数目(trabecularnumber,Tb.N)、骨小梁间距(trabecular separation,Tb.Sp)和连接密度(connectivity density,CD)。
1.2.3 血清中骨转换标志物检测 各组小鼠于DMM术前及术后2、4、6、8周麻醉后行心脏穿刺采血(采血前禁食6 h),室温静置30 min,4 000 r/min 20 min。分离血清并保存在-80°C冰箱,按照Elisa试剂盒说明测定Ⅰ型前胶原N末端前肽(Nterminal propeptide of typeⅠprocollagen,PINP)和Ⅰ型胶原羧基末端肽(carboxy-terminal telopeptide of type I collagen,CTX-Ⅰ)的浓度。
1.2.4 组织形态学染色 将获取micro-CT图像后的膝关节标本在10% EDTA-2Na中脱钙4周,石蜡包埋,沿胫骨内侧平台矢状面连续切取5µm厚的切片,按照试剂盒说明进行甲苯胺蓝染色、HE染色和TRAP染色。甲苯胺蓝染色:石蜡切片入二甲苯中脱蜡2次,20 min/次,梯度乙醇各1 min/次,自来水洗2 min。0.5%的甲苯胺蓝染液浸染30 min,自来水洗2 min,滤纸吸干水分,丙酮分化至软骨细胞呈蓝紫色,经梯度乙醇脱水后二甲苯透明,中性树胶封片。光镜下拍片后按照改良Mankin评分方法(表1)评估关节软骨的退变程度[14]。HE染色:苏木素染色3 min,自来水冲洗10 min,85%和95%乙醇脱水各5 min,伊红染色5 min。光镜下拍片并测量透明软骨层(hyaline cartilage,HC)、钙化软骨层(calcified cartilage,CC)厚度和关节软骨总厚度(total articular cartilage,TAC)。TRAP染色:切片在TRAP工作液中孵育2 h,蒸馏水冲洗后用苏木素染色3 min,光镜下随机选择切片的5个区域,应用ImageJ 1.48v软件计算TRAP+区域面积比值。

表1 改良Mankin评分
1.2.5 免疫组织化学染色DMM术后4周各组切片经脱蜡、水化后,置于0.01 M柠檬酸盐缓冲液中煮沸15 min,滴加3%的过氧化氢常温下孵育15 min,水洗后滴加1%的山羊血清封闭液并置于湿盒中室温下孵育1 h,在4°C下与兔抗小鼠Wnt5a、NF-κB、RANKL(稀释比例1∶200)孵育过夜。洗涤后分别用HRP和FITC标记的羊抗兔IgG二抗在37°C孵育1 h,洗涤并加入DAB显色后,苏木素和DAPI复染3 min。镜下随机选取切片的5个视野拍照并用ImageJ 1.48v软件计算阳性细胞数目。
1.2.6 Western blot检测术后4周,各组取5只小鼠用过量戊巴比妥钠麻醉处死,去除关节软骨,保留胫骨近端的软骨用于Western blot检测蛋白水平。在自动冷冻样品研磨机中粉碎组织,用含有蛋白酶抑制剂和磷酸酶抑制剂的缓冲液裂解组织。定量蛋白浓度并上样,用10%十二烷基硫酸钠-聚丙烯酰胺凝胶电泳分离蛋白,转膜后用脱脂牛奶封闭2 h以阻断非特异性蛋白结合,然后用兔抗小鼠Wnt5a、RANKL、IκBα、NF-κB工作液(稀释比例1∶2 000)孵育过夜。HRP标记的羊抗兔IgG二抗(稀释比例1∶10 000)孵育1 h,PBST洗膜后进行ECL发光,β-actin作为内参蛋白,Image Lab3.0软件测量灰度值。
1.3 统计学分析
采用GraphPad Prism 7.0统计软件进行统计学分析。计量资料采用均数±标准差(±s)表示,当满足正态分布且方差齐时,多组间比较采用单因素方差分析,不满足时则采用秩和检验。检验水准α=0.05。
2 结果
2.1 ZOL抑制OA小鼠的软骨下异常骨重塑
膝关节micro-CT结果显示,与sham+NS组相比,OA+NS组和OA+ZOL组在DMM术后4周软骨下骨吸收增加(图2A),术后8周骨形成增加(图2B),提示软骨下骨存在异常重塑(图2)。OA+NS组和OA+ZOL组在DMM术后4周时BV/TV、Tb.Th、Tb.N和CD降低,Tb.Sp增加(表2);术后8周时BV/TV、Tb.Th增加,Tb.Sp、Tb.N和CD降低(表3)。OA+NS组比OA+ZOL组变化显著,表明ZOL可以抑制OA早期软骨下异常骨吸收。。

图2 DMM诱导OA后4周和8周时胫骨软骨下骨的矢状位micro-CT图像,比例尺1000µm

表2 术后4周时软骨下骨micro-CT检测结果

表3 术后8周时软骨下骨micro-CT检测结果
2.2 ZOL抑制OA小鼠血清中骨转换标志物的异常改变
Elisa检测结果显示,与sham+NS组相比,OA+NS组在DMM术后4周内血清中CTX-I的含量逐渐增加,4周之后逐渐降低(图3B),PINP的含量逐渐增加(图3A),提示软骨下骨存在异常重塑。ZOL可以抑制OA小鼠血清中骨转换标志物的异常改变,延缓软骨下骨的异常骨重塑。

图3 Elisa检测血清中骨转换标志物的变化
2.3 ZOL抑制OA小鼠关节软骨的退变
DMM可诱导OA小鼠的关节软骨发生退变,并呈时间依赖性。甲苯胺蓝染色显示,与sham+NS组相比,DMM术后4周时OA+NS组的软骨细胞和基质的降解主要发生在软骨浅层,术后8周时,这些变化进一步发展至深层,并形成不规则裂隙(图4A中黑色箭头),逐渐累及全层关节软骨,Mankin评分逐渐增加(P<0.001,表4)。ZOL可以减轻软骨退变的程度,延缓OA进程,降低Mankin评分。HE染色显示,与sham+NS组相比,DMM术后随着OA病程的进展,OA+NS组的HC厚度逐渐降低(图4B中白线),CC随着潮线的复制逐渐增加(图4B中黑线)。ZOL可以减轻和延缓软骨的退变,降低CC/TAC的比值(表5)。

表5 膝关节的CC/TAC检测结果
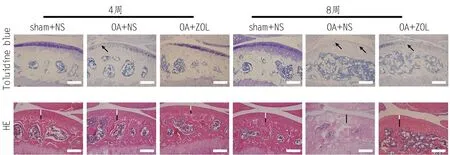
图4 ZOL对DMM诱导的OA小鼠关节软骨退变的影响

表4 膝关节的改良Mankin评分检测结果
2.4 ZOL抑制小鼠OA早期破骨细胞的形成及其相关蛋白的表达
TRAP染色显示,与sham+NS组相比,DMM术后4周时OA+NS组软骨下骨中TRAP+的破骨细胞数量增加,差异有统计学意义(P<0.001),ZOL可以抑制其增加(图5A),与micro-CT和Elisa检测结果相一致,支持ZOL抑制小鼠OA早期软骨下骨中破骨细胞活化造成的骨量减少(表6)。免疫组化染色显示,与sham+NS组相比,DMM术后4周时OA+NS组软骨下骨中Wnt5a+、NF-κB+的细胞比例均显著增加(图5B、C),免疫荧光染色显示RANKL+的细胞具有相同的结果(图5D)(P<0.001),ZOL可以抑制其增加(表7~8),与破骨细胞的变化相一致。

表6 破骨细胞的TRAP染色结果(%)

表7 Wnt5a、NF-κB的免疫组化染色结果(%)

图5 免疫组化检测ZOL对破骨细胞形成及其相关蛋白表达的影响

表8 RANKL的免疫荧光染色结果(%)
Western blot检测显示,与sham+NS组相比,DMM术后4周时OA+NS组软骨下骨中Wnt5a、RANKL、NF-κB的蛋白表达水平均增加,差异有统计学意义(P<0.001),拮抗NF-κB的IκBα蛋白表达水平降低,差异有统计学意义(P<0.01),ZOL可以抑制DMM导致的上述蛋白水平的改变(图6)。

图6 Western blot检测ZOL对破骨细胞形成相关蛋白表达的影响
3 讨论
OA是一种慢性、进行性、退变性疾病,软骨下异常骨重塑在OA的发生和发展中具有重要作用[4]。在OA早期,关节软骨发生退变之前,破骨细胞介导的骨吸收增强,骨量丢失,软骨下骨的显微结构变化导致微环境中生物力学和生物化学信号异常,进而激活骨形成相关的信号通路,造成软骨下骨发生硬化,骨赘形成,软骨退变[6]。OA的这一病理特点为通过早期干预软骨下异常骨吸收进而改善OA的治疗策略提供了依据。作为一种抑制破骨细胞骨吸收的药物,使ZOL通过抑制OA早期软骨下骨的异常骨吸收从而减缓OA进展的治疗策略成为可能。本研究采用DMM法建立小鼠OA模型,通过腹腔注射ZOL,可以有效延缓OA病程,减轻软骨退变程度,这与ZOL抑制Wnt5a/NF-κB信号介导的破骨细胞形成和骨吸收相关。
目前尚无一种OA动物模型能够完全模拟人类退变性OA,因为它是一种随着时间的进展而发生全关节受累的综合病变。尽管有自发性的小鼠OA模型应用于目前的基础研究中,但由于其漫长的病程限制了其在OA治疗中的应用[15]。关节腔注射化学药物的造模方法很难实现对OA各阶段,尤其是OA早期病理机制的研究。创伤后OA(post-traumatic osteoarthritis,PTOA)是OA的一个亚型,其终末转归与特发性OA相似,但OA早期的发生和进展与其他阶段潜在的发病机制和治疗方法是不同的。DMM模型模拟了临床上半月板损伤的病例,为研究OA进程中的结构性和生物性变化提供了可能。既往研究中报道的前交叉韧带离断(anterior cruciate ligament transection,ACLT)模型术后1~2周即可造成软骨下骨吸收增强和关节软骨的退变[6],所以本研究选择造模后2、4、8、12周为取材的时间点。相较于ACLT法所致的OA模型,DMM法创伤较小,出血少,诱发的炎症反应轻,对实验结果干扰小,而且该方法导致的OA进展缓慢,适合OA早、中期的病理机制研究[13]。本研究采用DMM方法构建OA模型,番红O-固绿染色和HE染色结果显示,DMM术后代表关节软骨退变程度的Mankin评分和CC/TAC比值渐进性增加,证实DMM法能够成功建立SD小鼠OA模型。DMM术后4周时,micro-CT结果显示软骨下骨的BV/TV、Tb.Th、Tb.N及CD降低,Tb.Sp升高,TRAP染色结果显示软骨下骨中的破骨细胞数量增加,这说明软骨下骨吸收增强;而ZOL可以减轻软骨下骨量的丢失,抑制破骨细胞的形成和骨吸收,血清中的骨吸收标志物CTX-I的增加进一步应证了这一点。
来源于造血干细胞的单核/巨噬细胞在形成具有骨吸收功能的成熟破骨细胞之前,需要迁移至骨吸收部位,并进一步融合,这一过程与NF-κB信号相关[16]。RANKL与破骨前体细胞表面的RANK结合后激活IκB激酶(IκB kinase,IKK),活化的IKK促使IκBα发生磷酸化并降解,NF-κB失去IκBα的限制后被释放并累积增多,然后转移至细胞核,启动下游信号中与破骨细胞形成和骨吸收相关的基因的表达。Wnt5a介导的非经典信号通路在骨与软骨的形成以及骨稳态的调节中发挥了重要作用,且与巨噬细胞介导的炎性和免疫性疾病的发生相关[9-10]。为了进一步研究ZOL抑制破骨细胞形成和骨吸收的分子机制,本研究检测了OA早期软骨下骨中与破骨细胞形成相关的蛋白表达水平。DMM术后4周,免疫组化结果显示软骨下骨中Wnt5a的表达增加,这与RANKL、NF-κB的增加呈正相关,而ZOL可以抑制Wnt5a、RANKL和NF-κB的增加,这一结果与Western blot的结果一致。上述结果说明,ZOL通过抑制Wnt5a/NF-κB信号减轻OA早期软骨下异常骨吸收,延缓OA病程,减轻关节软骨的退变,为实现OA的早期预防和精准治疗提供了重要依据。
破骨前体细胞的迁移与融合是其分化为成熟破骨细胞的前提,本研究基于动物体内实验探究了ZOL对小鼠膝OA的治疗效果和潜在机制,而ZOL对破骨前体细胞的迁移、融合以及对软骨细胞的直接作用及机制尚需进一步研究。
利益冲突所有作者均声明不存在利益冲突
