胶瘤菌属和孔生胶瘤菌
——中国新记录属、种
2022-11-16朱学泰冶晓燕范佳馨蒋长生
朱学泰, 杜 璠, 冶晓燕, 范佳馨, 蒋长生
( 1. 西北师范大学 生命科学学院, 兰州 730070; 2. 临夏回族自治州农业科学院, 甘肃 临夏 731100; 3. 连城国家级自然保护区管理局, 甘肃 永登 730333 )
胶瘤菌属(新拟)(Carcinomyces)由Oberwinkler和Bandoni于1982年建立,最初置于异担子菌纲(Heterobasidiomycetes)、胶瘤菌科(新拟)(Carcinomycetaceae)中,所属目当时未确定(Oberwinkler & Bandoni, 1982)。该属真菌通常寄生于某些担子菌子实体或植物上(Fungsin et al., 2002;Crous et al., 2019),常导致宿主的局部组织肿瘤状生长,形成透明至半透明胶质状菌瘿结构,新鲜时呈无色至淡黄色;其菌丝末端可同时形成分生孢子梗和担子。
孔生胶瘤菌(新拟)(Carcinomycespolyporina)发表于1970年,最初被置于银耳属(Tremella)中(Reid, 1970);1982年,Oberwinkler和Bandoni因其具菌瘿状的子实体,将其并入了胶瘤菌属(Oberwinkler & Bandoni, 1982);但是1986 年Ginns将胶瘤菌属的模式种Carcinomyceseffibulatus并入了链孢耳属(Syzygospora)中(Ginns, 1986),自此研究者就胶瘤菌属是否成立产生了争议,孔生胶瘤菌的归属问题也悬而未决。
近年来,银耳纲的分子系统发育分析研究结果显示,Carcinomyceseffibulatus(Ginns & Sunhede, 1978)、Carcinomycesarundinariae(Fungsin et al., 2002)和孔生胶瘤菌,应隶属于担子菌门(Basidiomycota)、银耳纲(Tremellomycetes)、银耳目(Tremellales)、胶瘤菌科、胶瘤菌属,与链孢耳属亲缘关系较远(Liu et al., 2015;Crous et al., 2019)。Crous 等(2019)依据分子系统学和形态学的证据,发表了物种Carcinomycesnordestinensis,所构建的系统发育树也支持胶瘤菌属成立,隶属于银耳目、胶瘤菌科。
胶瘤菌属真菌此前在我国未见有报道。本研究对采集自我国的1号该属真菌标本进行了形态解剖学与分子系统学研究。
1 材料与方法
1.1 标本信息
所研究标本于2019年8月1日采自甘肃省兰州市永登县,连城国家级自然保护区内棚子沟,针阔混交林中,地理位置为102°44′55.257″ E、36°37′42.288″ N,海拔1 941 m,主要植被为青海云杉(Piceacrassifolia)、青杄(P.wilsonii)、红桦(Betulaalbosinensis)等。标本采集号为ye629,标本馆保藏号为HKAS 115765。
1.2 形态学观察
使用Panasonic lux 10相机拍摄生态照片;用解剖镜(Motic SMZ-171)观察干标本的颜色、形状等;制作徒手切片,使用5%的KOH溶液制作水封片,在光学显微镜(ZEISS AX10)下观察显微结构,用Canon 70D相机拍摄显微照片。分别选取20个成熟的分生孢子和担孢子进行测量,长、宽以(a) b~c (d)表示,a和d分别表示测量的最小值与最大值,b、c表示90%的置信区间;孢子长宽比用Q表示,Qm= Q的均值 ± 标准差(Wu et al., 2016)。
1.3 分子系统学分析
用CTAB法提取标本的总DNA,对ITS(internal transcribed spacer内转录间隔区片段)进行扩增,引物为ITS5/ITS4(ITS5:5′-GGAA GGTA AAAG TCAA GG -3′,ITS4:5′- TCCT CCGC TTAT TGAT ATGC -3′)(White et al., 1990)。PCR产物由北京奥科鼎盛生物科技有限责任公司测序,所得的基因序列用BioEdit进行手动校对调整(Hall, 1999),在NCBI中进行BLAST比对,最终将准确的序列信息提交至GenBank库中。
参考Liu 等(2015)的研究结果,从GenBank数据库中选取相关的ITS序列,加上本研究获得的序列构建矩阵。使用RAxML 8.1.24软件基于最大似然法ML(maximum likelihood)构建系统发育树,碱基替代模型为GTRGAMMAI,其余参数设置均为默认值(Stamatakis, 2014);使用MEGA 7.0.26软件基于邻接法NJ(neighbor joining)进行系统发育分析;Bootstrap均重复1 000次以获得统计学支持。
所选序列的物种名称、来源地及GenBank登录号见表1。

表 1 构建系统发育树的ITS序列信息Table 1 Informations of ITS sequences used to construct phylogenetic tree
2 结果与分析
基于ITS 序列,采用最大似然法和邻接法构建系统发育树,二者的拓扑结构基本一致,仅支持率略有差异,最大似然法构建的系统发育树见图1,ML和NJ的支持率BP(Bootstrap percentages)分别用BP1和BP2表示。结果显示,本研究的序列与来自西班牙的孔生胶瘤菌标本AM20序列聚在一起,获得很高支持率(BP1=100, BP2=100),且与包括模式种Carcinomyceseffibulatus在内的胶瘤菌属物种以高支持率(BP1=99, BP2=98)聚为一支。

分支上从左到右分别为最大似然法(ML)和邻接法(NJ)经1 000次重复后的Bootstrap支持率。The Bootstrap percentages (BP) of maximum likelihood (ML) and neighbor joining (NJ) analyses from 1 000 replicates are shown respectively from left to right on the major branches resolved.图 1 基于ITS序列采用最大似然法构建的分子系统发育树Fig. 1 Molecular phylogenetic tree based on an ITS dataset using maximum likelihood
基于标本HKAS 115765进行的形态特征研究,结果如下。
寄生于多孔菌的子实层上(图2),形成半透明至透明的胶质状菌瘿,新鲜时稍具乳白色,干燥后变薄膜状,呈土褐色至黑褐色。分生孢子梗与分生孢子常见;分生孢子梗短棒状;分生孢子长椭圆至纺锤形,光滑,薄壁,无色,(3.5) 4.4~5.4 (8.0) × (1.5) 2.5~3.0 (4.5) μm,Qm= 1.80 ± 0.12。担子近球形,9.8~12.8 × 8.8~11.0 μm,具纵向分隔;小梗2个或4个,长可达8 μm;担孢子球形至近球形,无色,(4.8) 5.0~5.5 (6.0) × (4.5) 4.6~4.8 (5.1) μm,Qm= 1.10 ± 0.01,可萌发后形成分生孢子(图3)。有锁状联合。

A. 宿主菌的背面观; B. 宿主菌的腹面观; C. 宿主菌子实层及生长于其上的胶质状孔生胶瘤菌菌瘿。a. 胶质状的孔生胶瘤菌菌瘿。标尺: A, B=5 mm,C=2 mm。A. Dorsal view of parasitifer; B. Ventral view of parasitifer; C. Hymenium of parasitifer and the gelatinous mycocecidium of C. polyporina on it. a. Gelatinous mycocecidium of C. polyporina. Bars: A, B=5 mm, C=2 mm.图 2 孔生胶瘤菌及其宿主菌的宏观形态Fig. 2 Fruitbody of Carcinomyces polyporina and its parasitifer
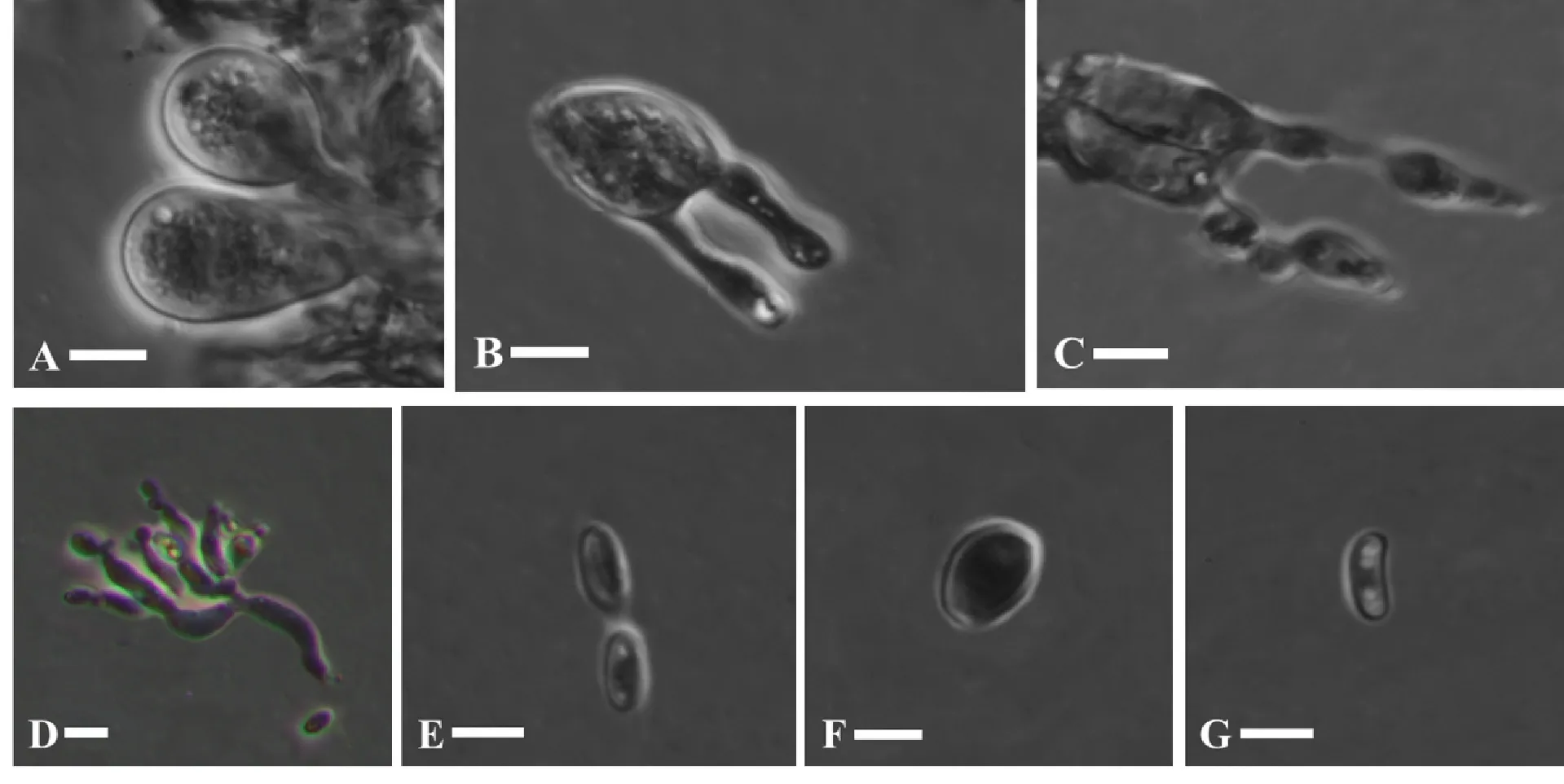
A. 孔生胶瘤菌的原担子; B. 孔生胶瘤菌纵裂的成熟担子; C. 孔生胶瘤菌的担孢子萌发; D. 孔生胶瘤菌的分生孢子梗; E. 孔生胶瘤菌的椭圆形分生孢子; F. 孔生胶瘤菌的近球形担孢子; G. 宿主菌的腊肠形担孢子。标尺=5 μm。A. Probasidium of C. polyporina; B. Basidium with longitudinally septation of C. polyporina; C. Basidiospore germination of C. polyporina; D. Conidiophore of C. polyporina; E. Elliptic conidia of C. polyporina; F. Subglobose basidiospore of C. polyporina; G. Botuliform basidiospore of parasitifer. Bars=5 μm.图 3 孔生胶瘤菌和宿主菌的显微结构Fig. 3 Microstructure of Carcinomyces polyporina and its parasitifer
综合形态特征与分子系统学研究结果,确认该标本为孔生胶瘤菌:
Carcinomycespolyporina(D. A. Reid) A. M. Yurkov, 2015
=TremellapolyporinaD. A. Reid, Trans. Br. Mycol. 1970
分布:英国(Reid, 1970;Roberts, 2007)、北美(Setliff, 1982)、西班牙(Millanes et al., 2011)、波兰(Karasiński & Wokowycki, 2015)、中国。
3 讨论与结论
本研究基于分子系统学与形态学的研究结果证实采自甘肃连城国家级自然保护区的标本HKAS 115765为Carcinomycespolyporina,是我国新记录属、种。Carcinomyces源于希腊文Καξχο,意为“癌症、肿瘤”,又因其胶质状的子实体,故将其中文名拟为“胶瘤菌”;其种加词“polyporina”表示该物种常寄生于多孔菌类真菌的子实层上,所以本文将该种的中文名拟定为“孔生胶瘤菌”。王耀等(2010)报道链孢耳菌(Syzygosporamycetophila)在中国的分布时,曾提及该物种所属的科Carcinomycetaceae,但并未给出相应的中文名称,本文根据词语含义,将其中文名拟定为“胶瘤菌科”。
系统分类研究方面,胶瘤菌属的模式种Carcinomyceseffibulatus曾被研究者置于链孢耳属中(Ginns,1986),链孢耳属目前隶属于线黑粉菌目(Filobasidiales)、线黑粉菌科(Filobasidiaceae)(Millanes et al., 2011;Liu et al., 2015),Index Fungorum系统目前采信这一处理, 将胶瘤菌属放置于线黑粉菌目、线黑粉菌科(http://www.indexfungorum.
org/Names/NamesRecord.asp?RecordID=823057)。但是,Liu 等(2015)构建的银耳纲系统发育树显示,Carcinomyceseffibulatus、C.arundinariae和C.polyporina以高支持率单独聚为一支,形成银耳目中的1个单属科——胶瘤菌科;链孢耳属的模式种Syzygosporaalba则位于线黑粉菌目、线黑粉菌科的分支上,与胶瘤菌属亲缘关系较远。
形态特征方面,胶瘤菌属物种大多寄生于担子菌的子实体上,在宿主上形成菌瘿,担子近球形或圆柱形,成熟后具有纵向或十字形分隔;线黑粉菌科物种多寄生在地衣上,担子管状或长棒状,无隔。目前,分子系统学和形态解剖学的证据均显示,胶瘤菌属应隶属于银耳目、胶瘤菌科,而非线黑粉菌目、线黑粉菌科,Index Fungorum系统中胶瘤菌属的系统发育位置相关信息需要进行订正。
胶瘤菌属目前确认的物种仅4种:Carcinomyceseffibulatus(模式种)(Ginns & Sunhede, 1978)、C.arundinariae(Fungsin et al., 2002)、C.nordestinensis(Crous et al., 2019)和孔生胶瘤菌(C.polyporina)(Reid, 1970)。作为寄生型的真菌,这些物种的寄生方式复杂而独特,如C.effibulatus目前仅发现寄生在栎金钱菌(Collybiadryophila)的子实体上(Ginns, 1986);Carcinomycesnordestinensis被发现寄生于红心凤梨(Bromeliaantiacantha)的叶片上,同时是紫花风铃木(Handroanthusimpetiginosus)的内生菌(Crous et al., 2019);孔生胶瘤菌的已知宿主有奶油波斯特孔菌(Tyromyceslacteus)和波状薄孔菌(Antrodiasinuosa)(Reid, 1970;Roberts, 2007)。本研究标本的宿主总DNA未能获得,仅根据子实体擦伤变蓝、担孢子腊肠形等形态特征,推测是波斯特孔菌或其近缘类群,准确的鉴定还有待更多标本的获得和研究。
孔生胶瘤菌此前在欧洲、美国均有报道(Reid, 1970;Setliff, 1982;Roberts, 2007;Millanes et al., 2011),此次在我国的发现,是该物种在东亚地区的首次报道。随着真菌多样性研究的广度和深度不断拓展,孔生胶瘤菌和胶瘤菌属其他物种必然会在更多的地方被发现,其分布规律和生态特征也会更加明晰。
