不同杉木无性系叶片解剖结构对大气增温的响应
2022-11-12李林鑫陶长铸林景泉马祥庆吴鹏飞
李林鑫,陶长铸,林景泉,李 明,马祥庆,吴鹏飞,*
1 福建农林大学林学院,国家林业和草原局杉木工程技术研究中心, 福州 350002 2 福建省上杭白砂国有林场, 上杭 364205
气候变化是当代社会发展面临的重大挑战之一,特别是近30年全球大气增温明显,21世纪全球气温将升高1.0—4.0℃,2020年全球多个国家(地区)出现有史以来最热年[1]。高温常伴随着水分短缺,使植物面临高温干旱的复合逆境胁迫,进而影响着植物生长发育概况[2]。Lienin等[3]对植物不同器官特征进行分析后,认为叶片直接暴露于外界环境,是植物长期适应环境过程中可塑性最大的器官,并在环境选择压力下形成不同适应型机制。高温在一定程度上会导致植物叶片细胞扩展、细胞壁变薄,单位面积上细胞数量和细胞层数减少,叶片蒸腾作用加强而出现大量失水现象[4]。然而,大黍(Panicummaximum)[5]、长瓣兜兰(Paphiopedilumdianthum)[6]、蒙古莸(Caryopterismongholica)[7]、杜鹃(Rhododendronspp.)[8]等植物可通过形态解剖结构改变,如增加叶片厚度、角质层厚度及构建不同发达程度的栅栏组织和海绵组织来调整细胞之间的紧密程度,以较好地吸收和保持水分,提高水分传输速率,从而有利于适应高温干旱的环境。叶片对环境的这种可塑性变化也反映出植物形态结构对外界环境因子的响应特征及生理生态适应策略[9]。在当前气候异常变化情况下通过叶片形态解剖结构的可塑性变化,探究植物对大气增温的响应机制至关重要。
杉木(Cunninghamialanceolata)是我国人工林种植面积最大的速生树种,广布于南方17个省(区),是我国重要的森林资源[10]。在全球气候持续变暖趋势下,南方亚热带地区大气温度增幅更是高于全球平均气温,气候变化成为杉木人工林长期健康与稳定生产力相当大的不确定来源[11]。Kang等[12]以不同气候变化情景通过FORECAST模型预测其对杉木人工林长期生长发育的潜在影响,结果表明温室气体CO2的增加可导致杉木水分利用效率增加及年生长期延长,从而对杉木人工林生产力产生了一定的积极影响。唐兴港等[13]通过MaxEnt模型和“ecospat”软件包进一步预测,未来杉木适生区分布受气候变暖影响沿经纬度方向迁移,除中度适生区分布范围呈一定程度扩大外,高度和低度适生区面积呈不断减少的趋势。可见,杉木对大气温度变化尤为敏感,有学者通过土壤电缆增温、开顶箱被动式增温等方式研究大气增温对杉木的影响机制,发现大气增温能显著增加杉木比叶面积和叶含水率,促使叶片生理功能提升,并改变杉木幼苗生长速率[14]。时应贵等[15]研究进一步发现,大气增温使1年生杉木叶片组织密度、比叶质量及2年生杉木叶片面积显著减少,叶功能特性改变明显。但目前相关研究并未深入到叶片解剖结构方面,同时,不同增温方式可能造成研究结果的差异,无法更好的弄清亚热带地区不断增加的大气温度对杉木叶片形态结构与功能特性的影响。此外,杉木分布区域跨度大,不同来源杉木(种源、无性系)对低磷[16]、干旱[17]、氮素[18]等逆境胁迫的响应及养分利用效率机制存在差异,那么,这种差异是否也在杉木叶片解剖结构对大气增温响应机制中体现。近年来,我国南方一些局部地区呈现出连年高温少雨现象,杉木人工林不但面临着林地养分匮乏的困境,大气增温带来的高温干旱复合逆境胁迫对杉木人工林生长发育的影响机制,以及杉木如何调整自身形态结构适应高温干旱的生存策略尚不明确。因此,通过叶片解剖结构探究杉木在大气增温背景下的响应机制及生存策略对杉木人工林长期科学经营具重要意义。
鉴于此,本文以福建省上杭白砂国有林场2年生杉木无性系试验林为研究对象,在造林后对研究地历年气候因子变化进行分析基础上,从叶片解剖结构角度出发,研究10个杉木无性系叶片解剖结构在大气增温下形态特征的差异,以期进一步认识杉木在气候变化下的形态可塑性变化及响应机制,从而更好的为杉木优良品种筛选和人工林栽培管理提供科学依据。
1 研究地概况
试验地设在福建省上杭白砂国有林场(25°01′—25°16′N,116°20′—116°39′E),海拔200—700 m,坡度10—40°,土壤以粗粒花岗岩发育而成的砂质红壤为主,土层厚度60cm以上,林地立地等级以II、III类地为主[19]。2019年3月选择国家林业和草原局杉木工程技术研究中心及福建省洋口国有林场前期筛选的10个健康无病虫害、长势一致的1年生杉木优良无性系苗S23、洋061、W2、P18、S4、P17、P41、P11、S22、P32造林[20—21]。前茬为杉木人工林,林地采伐后进行劈草,带状整地,带宽2.0m,株距1.8m,挖明穴,规格为60cm×40cm×40cm,每个无性系按纵向排列方式顺坡种植,造林后当年进行全面锄草抚育1—2次。林下植被主要有芒萁(Dicranopterispedata)、狗脊(Woodwardiajaponica)、五节芒(Miscanthusfloridulus)等。
基于上杭县气象站(25°03′N,116°25′E)观测数据(国家气象科学数据中心,http://data.cma.cn),研究地2010—2020年气候变化情况如表1所示。其中,年均气温整体上呈上升趋势,2020年出现最大值,为21.08℃,相比2019年增加0.35℃,而年温最大极值每年均超过35℃的高温界限值,2020年达到了36.59℃,相比2019年增加0.5℃。10年内年均降水量为1723.37mm,年均相对湿度为75.86%。从该地区多年气候变化情况来看,该区域气温变化呈明显的增加趋势,在2020年气温出现10年来最大值,降水量相较于前几年有所减少。
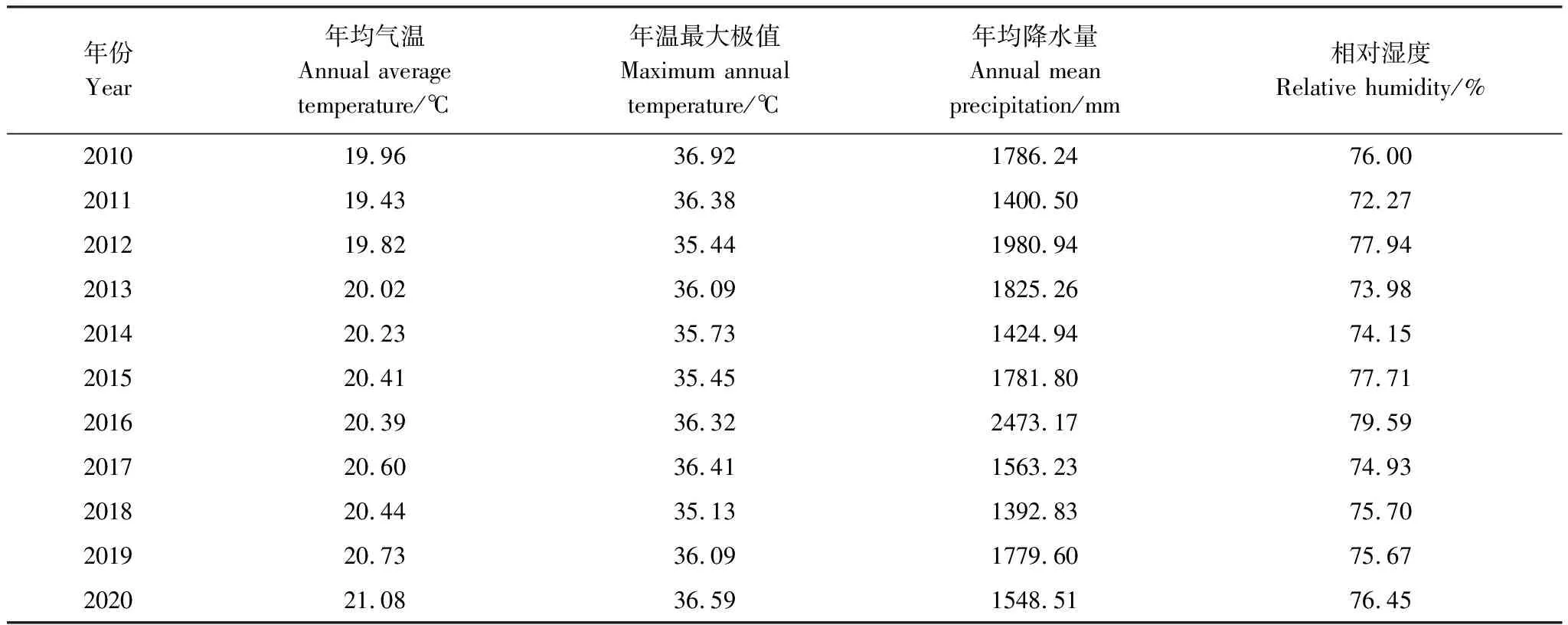
表1 研究地2010—2020年不同气候因子的变化情况
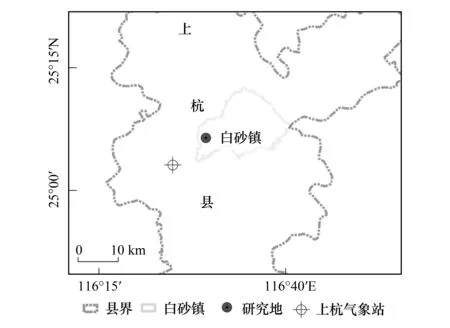
图1 研究地与上杭县气象站的地理分布 Fig.1 Geographical distribution map of experimental field (Baisha) and relationship with the meteorological station(Shanghang)
该气象站海拔205m,与研究地所处地理位置相近,且同属于福建省龙岩市上杭县(图1)。
2 研究方法
2.1 叶片采集与处理
2020年10月,对研究地10个杉木无性系幼林进行每木检尺,测定树高,分别选择平均木3株,剪取每株幼树顶端第一轮侧枝当年新展开的针叶3—4片,要求无病虫害、生长良好。从每片针叶中部切取长0.5cm的针叶小段样品3—5个放入装有FFA固定液(70%酒精90mL:冰醋酸5 mL:福尔马林5mL)的玻璃瓶中,用针筒手动抽气5 min,然后置于装有冰袋的泡沫箱中带回实验室待测。
采用微波石蜡切片技术对针叶样品进行脱水、透明与浸蜡,包埋后经半薄切片机(Leica RM2265)切片(厚为6μm)、粘片与烘片、脱蜡、番红-固绿染色、中性树胶封片[22]。每株平均木制作横向切片2张,即每个无性系制作切片6张,平均每张切片上有叶片横切面20—30个,从中随机选择6个完整样点用Nikon(ECLIPSE E200)电子显微镜拍照并观察。
采用ScopeImage 9.0软件对叶片上表皮角质层厚度、上表皮厚度、下表皮角质层厚度、下表皮厚度、栅栏组织厚度、海绵组织厚度、叶片厚度和维管束厚度进行参数测量。根据参数测量结果按以下公式计算栅海比(P/S)、叶片组织结构紧密度(CTR)、叶片组织结构疏松度(SR),可塑性指数(PI)和叶片变异系数[CV(%)]:
栅海比(P/S)=栅栏组织厚度/海绵组织厚度
(1)
叶片组织结构紧密度(CTR)=(栅栏组织厚度/叶片厚度)×100%
(2)
叶片组织结构疏松度(SR)=(海绵组织厚度/叶片厚度)×100%
(3)
可塑性指数(PI)=(最大值-最小值)/最大值
(4)
变异系数[CV(%)]=(标准差/算术平均数)×100%
(5)
2.2 数据分析
采用SPSS 25.0软件对数据进行统计分析,结果以平均值±标准差表示。利用单因素方差分析(One-way ANOVA)比较不同杉木无性系叶片解剖结构指标之间的差异,Duncan多重比较法用于差异性检验分析(α=0.05),同时采用Pearson相关性分析和主成分分析(principal component analysis,PCA)对不同杉木无性系叶片解剖结构指标、P/S、CTR、SR和气象因子进行分析,并根据叶片变异系数和可塑性指数评价不同杉木无性系对环境表现的潜在适应能力。使用Origin 2018、ArcGIS 10.2软件绘图。
3 结果与分析
3.1 研究区域2020年降水量和气温的月变化
结合当地近10年历史气候特征(表1),2020年是福建省极少见的高温年。该区域5—9月平均气温达到27.13℃,平均降水量只有202.79mm,其中,7月平均气温与月最大极值气温达到最大值,分别为27.25℃和34.52℃,月均降水量变化趋势与之相反,7月份最低,为74.99mm(图2)。从全年各主要气候因子变化趋势可以发现,该区域存在明显的干湿季节变化:秋冬季低温少雨,春季气温逐渐回升,降水量有所增加,但自进入夏季,气温与降水量变化明显,在气温升高的同时降水量呈迅速下降的态势。可见,该区域夏季有明显的高温干旱现象。
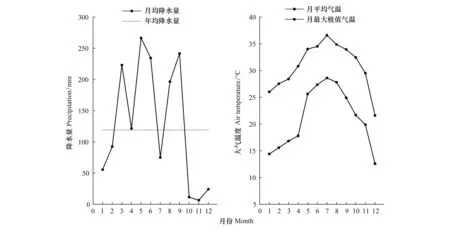
图2 研究地2020年降水量和大气温度的月变化特征Fig.2 Monthly variation characteristics of precipitation and atmospheric temperature in 2020 for the study site
3.2 不同杉木无性系叶片解剖结构特征
从10个参试杉木无性系叶片解剖结构横切面的整体情况来看(图3),其形态结构基本相同,均有明显的栅栏组织和海绵组织分化,为典型的异面叶。叶片解剖结构从上往下,依次为上表皮、栅栏组织、维管束、下表皮和紧邻维管束的海绵组织。表皮外附着明显的角质层,靠近上表皮的长柱形或椭圆形细胞为栅栏组织,层数为1—2层,其长轴与上表皮垂直,越往叶片边缘排列紧密且整齐,第二层细胞体积小于第一层。海绵组织细胞较大,呈长椭圆形,相互之间连接成网状。此外,不同杉木无性系叶片树脂道数量存在差异,无性系S23、洋061、P18、S4、P17、P41、P11、S22只有中间树脂道,而W2、P32存在左右侧树脂道,数量为2个(中间存在1个主树脂道,左右各存在1个侧树脂道,图3)。
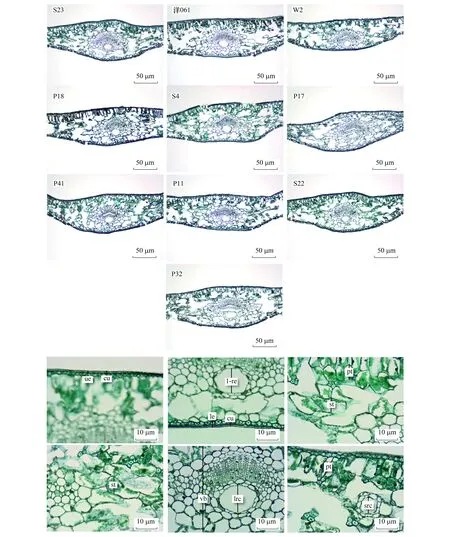
图3 不同杉木无性系叶片横切面解剖结构Fig.3 Anatomical structure of leaf cross section of different Chinese fir clonesS23、洋061、W2、P18、S4、P17、P41、P11、S22、P32(×10)。cu:角质层 Cutical;ue:上表皮 Upper epidermis;le:下表皮Lower epidermis;pt:栅栏组织 Palisade tissue;st:海绵组织 Spongy tissue;vb:维管束 Vascular bundle;lrc:中间树脂道 Intermediate resin canal;src:侧树脂道 Side resin canal(×40)
经方差分析,不同杉木无性系叶片解剖结构指标存在不同程度的差异(图4)。在表皮特征方面,上表皮角质层厚度变幅为1.18—1.59μm,其中P32厚度最大,除与S23、S4、P11、S22差异不显著外(P>0.05),与其它无性系存在显著差异(P<0.05)。上表皮厚度变幅为4.65—5.64μm,洋061厚度最大,与P32的差异达显著水平(P<0.05),前者是后者的1.21倍。下表皮角质层厚度变幅为0.87—1.28μm,S23厚度最大,但与洋061、P41、P11、S22、P32的差异未达显著水平(P>0.05)。下表皮厚度变幅为3.93—4.74μm,不同无性系间差异未达显著水平(P>0.05)。
在叶肉组织方面,栅栏组织厚度变幅为6.12—13.08μm,海绵组织厚度变幅为9.87—16.96μm,所有无性系海绵组织厚度均大于栅栏组织厚度。其中,S23的栅栏组织厚度和海绵组织厚度均小于其它无性系;洋061、W2、P32 的栅栏组织厚度显著大于其它无性系(P<0.05),洋061、W2、P18、P41、P11、P32的海绵组织厚度显著大于其它无性系(P<0.05)。在叶片厚度方面,无性系杉木叶片厚度变幅为89.62—113.21μm,最大的是P32,显著大于其它无性系(P<0.05)。在维管组织方面,维管束厚度变幅为44.62—56.92μm,P32最大,除与S23、S4、P17不存在显著性差异外(P>0.05),均显著大于其它无性系(P<0.05),P18最小。
通过叶片解剖结构指标参数计算出P/S、CTR、SR,由图5可知,P/S值变幅为0.54—0.81,CTR变幅为6%—13%,SR变幅为10%—19%。其中洋061的SR最大,W2的P/S和CTR均大于其它无性系,相比较而言,S23的CTR和SR均最小,P17和S22的P/S同为最小值。
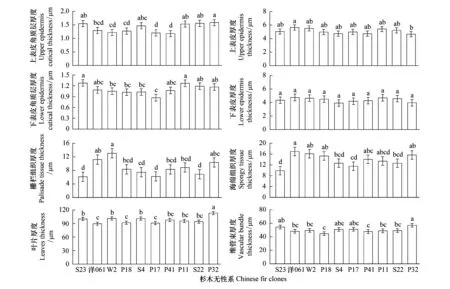
图4 10个杉木无性系叶片解剖结构特征参数Fig.4 Characteristic parameters of leaf anatomical structure of ten Chinese fir clones图中不同小写字母表示无性系间具有显著差异(P<0.05)

图5 不同杉木无性系叶片叶肉组织特征 Fig.5 Leaf mesophyll tissue characteristics of different Chinese fir clones
3.3 不同杉木无性系叶片解剖结构与气候因子的相关性分析
由相关关系分析结果可知(表2),上表皮角质层厚度与下表皮角质层厚度、栅栏组织厚度与海绵组织厚度、叶片厚与维管束厚度呈极显著正相关关系(P<0.01),P/S与栅栏组织厚度、海绵组织厚度均呈极显著正相关关系(P<0.01)。月平均气温与栅栏组织厚度呈极显著正相关关系(P<0.01),月均降水量与栅栏组织厚度、海绵组织厚度、P/S均呈极显著正相关关系(P<0.01),气候因子相互之间也呈极显著正相关关系(P<0.01)。另外,下表皮厚度与上表皮厚度、下表皮角质层呈显著正相关关系(P<0.05),P/S与下表皮厚度呈显著正相关关系(P<0.05),CTR与上表皮厚度、栅栏组织厚度、海绵组织厚度及月均降水量均呈显著正相关关系(P<0.05),而维管束厚度与上表皮厚度呈显著负相关关系(P<0.05)。可见,气温与降水量影响杉木叶片解剖结构变化,但不同解剖结构指标对气候因子的响应存在不同程度的差异。
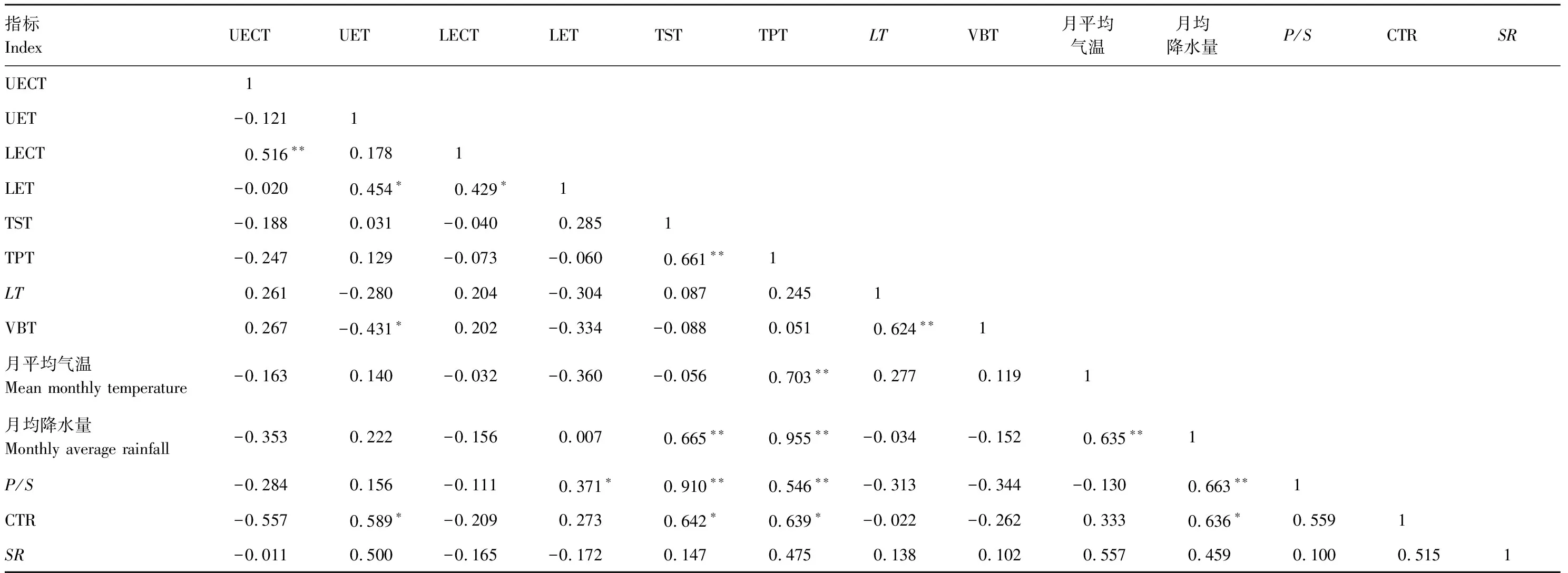
表2 不同杉木无性系叶片解剖结构与气候因子相关性分析
3.4 不同杉木无性系叶片解剖结构的主成分分析
以不同杉木无性系叶片解剖结构指标、P/S、CTR和SR为分析因子进行主成分分析。根据分析结果,由相关系数矩阵得出特征值、贡献率和累计贡献率,依据特征值>1,提取的3个主成分累计方差贡献率为89.449%(表3),可反映出杉木叶片解剖结构特征参数的综合信息。根据各项特征参数在各主成分上的因子载荷分析表明,第1主成分中具有较高载荷值的是海绵组织厚度、栅栏组织厚度、CTR和P/S,主要反映叶肉组织结构特征。第2主成分中具有较高载荷值的是下表皮厚度、叶片厚度、上表皮厚度和维管束厚度,主要反映叶片支撑和输导水分特征。第3主成分中具有较高载荷值的是上表皮角质层厚度和下表皮角质层厚度,主要反映叶片保水和持水能力。由于海绵组织厚度、栅栏组织厚度、CTR和P/S对第1主成分贡献最大,叶片厚度、上表皮厚度和下表皮厚度对第2主成分贡献最大,上表皮角质层厚度和下表皮角质层厚度对第3主成分贡献最大。因此,海绵组织、栅栏组织、叶片厚和表皮组织可视为反映杉木叶片解剖结构特征的主要指标,其次为角质层。
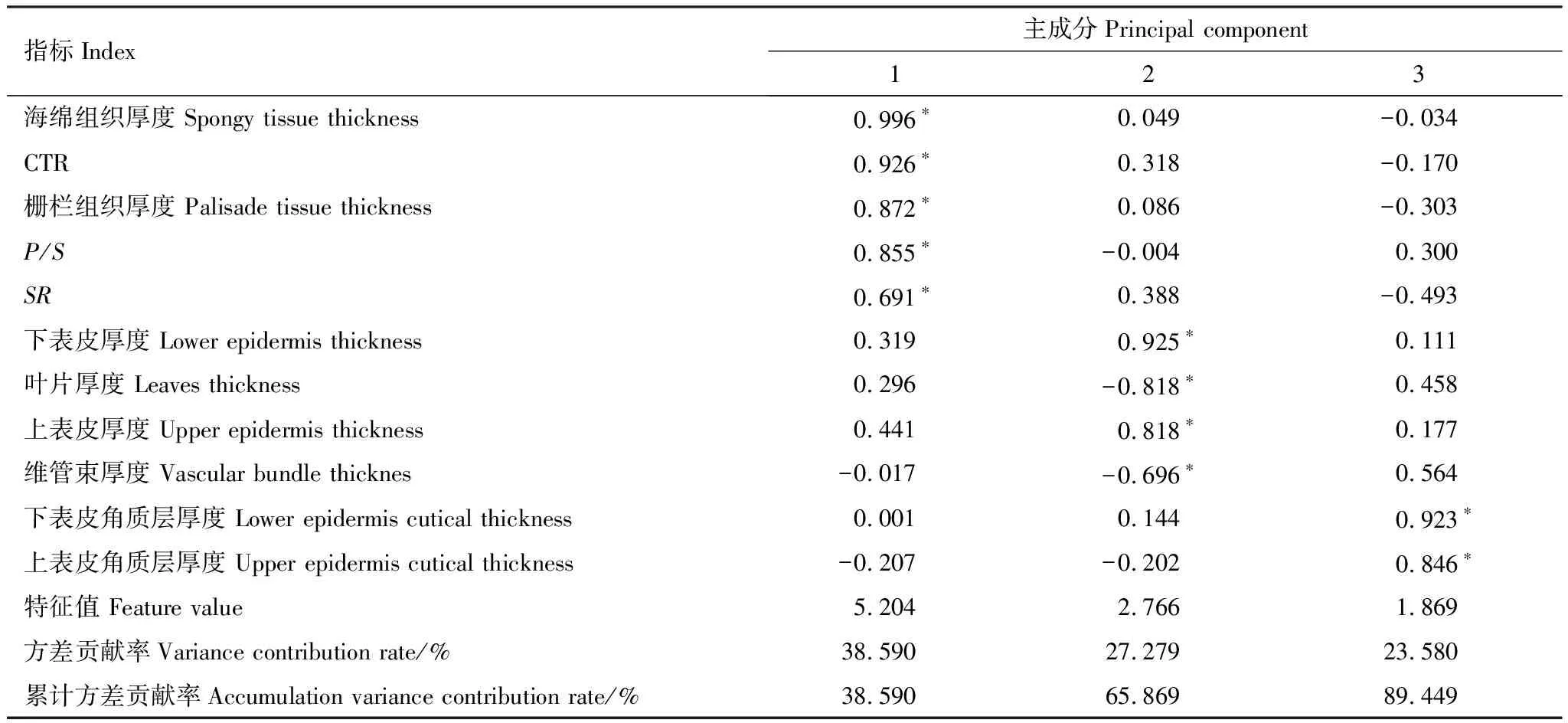
表3 旋转后叶片解剖结构因子载荷矩阵及特征根
3.5 不同杉木无性系叶片解剖结构变异系数和可塑性指数
由表4可知,不同杉木无性系叶片变异系数和可塑性指数存在差异,叶片变异系数在9.28%—24.16%之间,可塑性指数在0.13—0.44之间,但其均值排序结果一致,为W2>P11>S4>P32>P41> P17>P18>S22>S23>洋061。另外,叶片解剖结构指标变异系数大小均值排序为:P/S>CTR>SR>海绵组织厚度>栅栏组织厚度>下表皮厚度>下表皮角质层厚度>上表皮厚度>上表皮角质层厚度>维管束厚度>叶片厚度。
4 讨论
4.1 杉木叶片解剖结构变化与气候因子的关系
气候因子被认为是引起叶片解剖结构可塑性变化的主导因素之一[23]。本文研究发现,月平均气温与栅栏组织呈极显著正相关关系(P<0.01),月均降水量与栅栏组织、海绵组织、栅海比均存在极显著正相关关系(P<0.01),说明水分相比气温更易引起杉木叶片解剖结构变化,但气温和降水量对杉木叶片上表皮厚度、角质层厚度等其余解剖结构指标影响不明显,两者间相关性未达显著水平(P>0.05)。研究表明,叶片栅栏组织是植物进行光合作用的重要场所,海绵组织与气孔器孔下室形成的通气系统有利于植物光合作用及气体交换,而温度和水分是植物光合作用进行的主要条件,对叶片栅栏组织和海绵组织影响较大[24]。另外,研究地2020年夏季有明显的高温干旱现象,杉木在面临大气温度升高的同时,还受环境水分亏缺的影响。因此,相比于其它解剖结构指标,杉木叶片栅栏组织和海绵组织与气温和降水量间的相互关系更加明显。
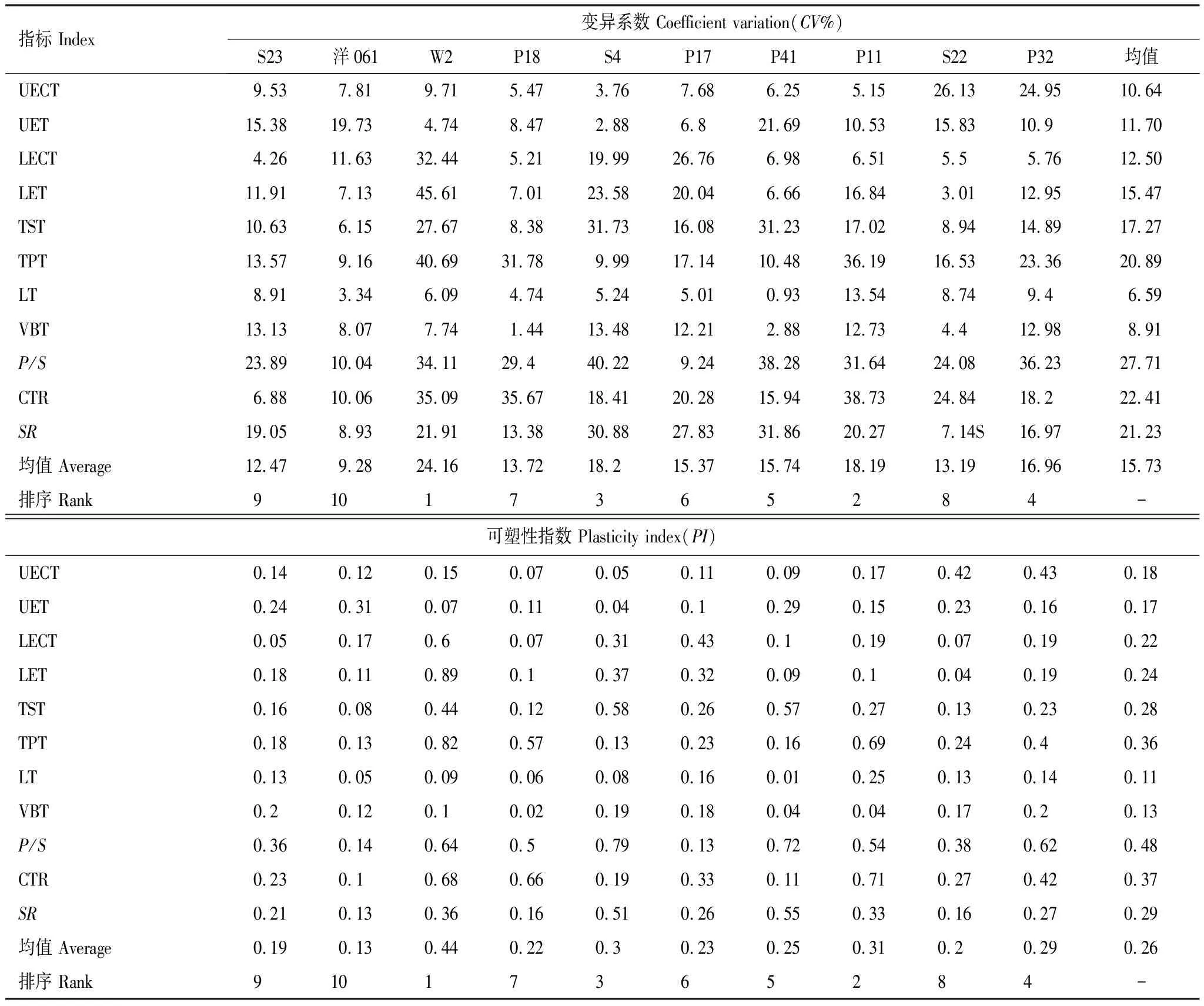
表4 叶片解剖结构的变异系数与可塑性指数
本研究中,叶片厚度与维管束厚度呈显出极显著正相关关系(P<0.01),意味着杉木在叶片厚度增加的同时还趋向于构建发达的维管组织,这种结构有助于叶片在降水量较少环境下贮藏更多水分,并将其尽量多的运输至内部,极大程度上提高叶片水分运输效率[25]。同时这也是很多木本植物在水分缺失下的普遍特征[26]。另外,杉木叶片解剖指标相互之间存在自相关关系,说明杉木叶片组织结构之间存在一定的协同变化关系,这种组织结构方面的协同性进一步表明植物形态结构是在适应环境变化过程中而进化出的生存策略,这种生存策略与叶经济谱概念相吻合[27]。
4.2 不同杉木无性系对大气增温响应的差异
叶片形态解剖结构指标变化在一定程度上反映植物因自身遗传分化而对外界环境因子影响表现出的响应特征[28]。本文研究发现,叶片角质层厚度、栅栏组织厚度等解剖结构指标在杉木无性系间表现出不同程度的改变。研究发现,杉木由于分布区生境差异较大及生态遗传隔绝,在长期系统发育过程中,会形成对生长环境不同需求的遗传变异[29]。因此,这种现象可能是不同杉木无性系在遗传因子影响下对环境大气温度变化的适应而在叶片形态结构上表现出的短期响应特征。在本研究中,杉木无性系叶片解剖结构指标变异系数均值在6.59%—27.71%之间,平均值为15.74%,变异系数最大的是栅海比,最小的是叶片厚度。陈旭等[30]认为,叶片解剖结构指标差异是植物应对环境过滤的结果,其形态结构变异系数大小可反映植物对环境适应形成的变异能力,其值越大,变异性越强。本文研究结果表明杉木叶片解剖结构遗传分化明显,不同无性系选择潜力较大,对环境适应的变异能力也确实表现出一定差异。
众多学者利用可塑性指数来评价植物对环境所表现的潜在适应能力,可塑性指数越大表明其具有更宽的生态幅和更好的逆境耐受性,在优良品种筛选时选择空间也越大[31]。因此,为进一步挖掘对大气增温带来的高温干旱复合胁迫潜在适应能力较强的杉木无性系,本文通过可塑性指数对其响应大气增温的能力做出评价。结果表明,不同杉木无性系叶片可塑性指数均值大小排名为W2>P11>S4>P32>P41>P17> P18>S22>S23>洋061,其中无性系W2叶片可塑性指数为0.44,是无性系洋061的3.38倍。钟悦鸣等[32]发现,荒漠湿地中胡杨(Populuseuphratica)叶片解剖结构相较于绿洲河岸林具有更高的可塑性指数,其更能够适应高温干旱的生长环境。胡营等[33]也认为,在干旱条件下,叶片可塑性指数大的植物能有效地减少水分散失,对水分亏缺的环境具有较强适应能力。因此,从叶片可塑性指数大小的角度来说,无性系W2相比其它杉木无性系具有更强的高温干旱适应能力,可成为未来应对气候持续变暖的良好造林材料。而无性系P11、S4和P32等叶片可塑性指数虽小于无性系W2,但植物对高温干旱环境的适应还包括内在生理特性的改变及生长性状变化,从而形成对大气增温过程的相对稳定性和适应性。因此,这些无性系也为深入研究杉木对大气增温的响应机制提供了优良选择材料。
4.3 杉木无性系叶片解剖结构对大气增温的响应机制
叶片作为植物适应复杂异质环境重要的器官,其组织结构变化是应对环境因子影响的一种重要响应方式,同时也体现出植物在适应环境改变过程中形成的内在响应机制[34]。本文研究发现,10个杉木无性系均为异面叶,综合主成分分析筛选出的海绵组织、栅栏组织、叶片厚度、表皮组织和角质层在生物体中的功能特性可知,10个杉木无性系叶片解剖结构特征表现出抵抗强烈光照和减少水分散发的高温干旱响应机制。但10个杉木无性系叶片解剖结构的特征不同,对高温干旱环境所表现的响应机制也有所差异,如上表皮厚度普遍大于下表皮厚度,而气孔主要分布于下表皮,这在一定程度上缩短了气体运输距离,有利于减少杉木在高温干旱环境下体内能量的消耗[35]。据研究报道,在干旱缺水环境中,植物叶片上表皮厚度越大越利于叶片进行蒸腾作用,同时还能避免光照对叶肉细胞造成的灼伤[36]。因此,这种结构也有利于杉木叶片在强烈光照和水分欠缺条件下与外界进行气体交换,体现出杉木叶片对高温环境的一种响应机制。人们普遍认为结构是功能的基础[37],虽然无性系P32上表皮角质层厚度和下表皮角质层厚度与无性系S23、P11、S22和S4差异均未达显著水平,但其叶片厚度明显较大,这表明无性系P32相比较于无性系S23、P11、S22还通过增加叶片厚度的方式来适应高温干旱环境,这与处于降水量稀缺环境下不同杨树(Populusspp.)无性系[38]、锦鸡儿属(Caragana)[39]植物表现的现象一致。
另外,不同杉木无性系叶片输水组织存在差异,10个杉木无性系除拥有发达的维管组织及中间树脂道外,无性系W2、P32相比于其它无性系,在左右各拥有1个侧树脂道。Rodríguez-García等[40]认为,树脂道数量、维管束厚度与水分运输和生长速率密切相关,环境降雨量的多少对其形成造成重要影响,发达的输水组织结构有利于提高叶片水分运输能力,增强植物在高温干旱环境下的生存能力。但无性系W2、P32树脂道数量增加的这种解剖结构特性是否有利于增强其对高温干旱的适应还有待进一步研究。本研究还发现,参试的10个杉木无性系叶片下表皮厚度差异不显著(P>0.05),这可能是因为植物叶片表皮组织是其抵御外界环境不利因素的重要屏障,在内部水分缺失情况下,表皮细胞越厚能更大程度限制水分蒸散,有利于杉木适应研究地极端的高温环境,这种现象也进一步体现出杉木无性系叶片解剖结构存在趋同进化现象[41]。
5 结论
本文基于当前气候异常变化的特点,分析10个杉木无性系叶片解剖结构对大气增温的响应特征与机制,研究得出:(1)研究地2020年夏季有明显的高温干旱现象,杉木叶片解剖结构在10个参试的无性系间存在不同程度改变,表现出对大气增温响应的形态可塑性差异,从而形成不同的响应机制。(2)气候因子与叶片解剖结构指标之间存在一定相关关系,气温与栅栏组织厚度呈极显著正相关,降水量与栅栏组织厚度、海绵组织厚度、P/S均呈极显著正相关,这表明气温与降水量是引起杉木叶片解剖结构变化的重要气候因子,但其对气候因子的响应程度存在一定差异。(3)不同杉木无性系叶片变异系数与可塑性指数存在差异,对大气增温表现出不同程度的响应,其中无性系W2具有较大的叶片可塑性指数,对高温干旱环境表现出较强的潜在适应能力。(4)不同杉木无性系通过叶片组织结构功能调整与平衡形成各自应对大气增温的响应机制,其中叶片海绵组织、栅栏组织、叶片厚度、表皮组织和角质层为表征杉木叶片解剖结构的典型指标,表现出对高温干旱环境的良好适应性。然而,植物对环境的适应不仅与各器官解剖结构特征有关,还受内在生理生化活动与遗传特性影响,本研究从叶片解剖结构角度对杉木在大气增温下的响应机制进行初步探究,今后应综合考虑其它环境因子对其叠加效应并结合生理生化特性进行深入研究,从而进一步揭示杉木对大气增温的适应机制。
