持续性水分胁迫对冬小麦光合特性及产量的影响*
2022-11-10马雅丽郭建平刘文平
马雅丽 郭建平 栾 青 刘文平 李 蕊
1 山西省气候中心,太原 030006 2 中国气象科学研究院,北京 100081 3 南京信息工程大学气象灾害预警预报与评估协同创新中心,南京 210044 4 山东省威海市气象局,威海 264200
提 要:通过设置对冬小麦起身—灌浆期不同阶段的充分灌水试验,即分别在起身期(T1)、起身—拔节期(T2)、抽穗—灌浆期(T3)进行充分灌水处理,模拟自然状态下土壤从水分饱和到产生干旱的整个过程,并以起身—灌浆初期各发育期充分灌水处理为对照(CK),研究持续性水分胁迫对冬小麦光合作用及产量的影响,为冬小麦春季大田生产中选择充分灌溉时机提供参考。结果表明:持续性水分胁迫下冬小麦旗叶的叶绿素含量、净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)和胞间CO2浓度(Ci)均有所下降,且光合日变化峰值不同程度提前。T1、T2、T3处理的冬小麦Tr和Ci在不同发育阶段均比CK差异显著,但T1、T2、T3处理之间在灌浆期差异不显著。在T3受到持续性水分胁迫,即使后期复水,相比仅在T2受到持续性水分胁迫来说,其叶绿素含量、Pn和Gs降低更明显。冬小麦在T3受到持续性水分胁迫,造成冬小麦穗数和穗粒数减少最明显,对产量影响最大。
引 言
干旱是世界上危害最广泛、最严重的自然灾害之一,中国是干旱灾害发生频率最高且影响最严重的国家之一(马柱国等,2018;张强等,2020)。干旱也是全球小麦生产中备受关注的主要气象灾害,华北平原是中国冬小麦主产区,严重水分胁迫往往是造成冬小麦减产的重要原因(曹雯等,2019)。不同时段出现不同程度水分胁迫,尤其是返青以后的水分胁迫,会对冬小麦株高、叶面积、干物质累积、根冠比等农艺性状及产量造成不同程度的影响(霍治国等,2001;杨贵羽等,2006;姚宁等,2015;信志红等,2019;吕丽华等, 2014),归根到底是由于水分胁迫影响了冬小麦植株光合作用,导致干物质累积和分配发生变化,影响生长发育过程和最终产量。作物干物质90%以上来自光合作用,光合作用是物质生产和产量形成的重要生理过程,是揭示作物对不同生境反应机制的重要途径。干旱的识别很多建立在气象因子上(金燕等,2018;左冰洁和孙玉军,2019;姜丽霞等,2021),而农作物主要是通过吸收土壤中的水分维持正常的生长发育(郭建平等,2020),随着气候变化背景下农田生态系统水分供需矛盾日益凸显(张丽敏等,2021;马雅丽等,2020),作物干旱通常以土壤水分和作物蒸散为主要因子(谢五三等,2019)。在不同灌水处理下系统研究不同时段水分胁迫对冬小麦光合特性和产量的影响,对理解作物抗逆机理、指导抗旱决策意义重大。
国内外学者针对水分胁迫对光合特性的影响开展过大量研究,对深入理解作物抗旱机理具有重要作用。在不同水分胁迫对冬小麦光合参数影响方面,张建军等(2008)得到抽穗后长期水分胁迫使净光合速率明显下降,抽穗后适度的水分胁迫虽然使气孔导度和蒸腾速率下降,但复水后二者均明显提高。宁东峰等(2010)研究表明,轻度水分亏缺条件下,气孔导度、蒸腾速率下降,而光合速率得以维持,单叶水分利用率提高,但生育前期重度水分亏缺对后期旗叶光合速率有显著影响。米慧聪等(2017)发现适度水分亏缺对旱作小麦穗部净光合速率影响较小。轻度水分胁迫条件下叶片净光合速率的降低与植株叶片气孔关小以及CO2摄入量减少有关,但重度水分胁迫时光合速率下降主要受生化代谢过程限制(Flexas and Medrano,2002;薛崧等,1992)。在水分胁迫对光合参数日变化影响方面,裴冬等(2006)得出水分胁迫使冬小麦光合速率的峰值提前出现,蒸腾速率比光合速率对水分胁迫的反应更敏感,更易受气孔调节的影响。在水分胁迫对光合产物和产量影响方面,胡程达等(2014)发现干旱胁迫时间延长至扬花期后,前期的自身调节并不能阻止重度干旱对冬小麦的伤害,干旱会对整株水平的光合产物累积、分配和利用产生影响。严韬等(2019)研究表明返青期—成熟期充分供水的冬小麦旗叶光合参数和产量最高,开花—成熟期轻度干旱及全生育期轻度干旱的冬小麦光合特征参数与产量均显著降低。马青荣等(2020)试验表明,不同干旱条件下冬小麦根系活力和产量呈显著性相关,水分利用效率呈显著性负相关,重大干旱年在拔节期灌水可达到抗减损节水灌溉效果。
上述研究多集中在阶段性水分胁迫和不同亏缺灌溉对冬小麦的光合特性及产量的影响方面,但实际生产中通常是在春季充分灌溉一到两次,也相当于春季的透墒雨。在冬小麦返青以后营养生长期或生殖生长期充分灌溉(或有透墒雨)的条件下,其余时段的持续性水分胁迫对华北地区冬小麦不同生育期的光合特性及产量的研究较少。本文拟通过对冬小麦起身期、起身—拔节期、抽穗—灌浆初期不同时期进行充分灌水处理,研究不同阶段持续性水分胁迫对冬小麦光合特性、产量及产量构成要素的影响,揭示光合作用对持续性水分胁迫的响应机制以及对产量形成的作用机理,为华北地区冬小麦合理灌溉提供参考。
1 材料与方法
1.1 试验设计
试验于2017年10月至2018年6月在河北省定兴县固城镇中国气象局生态环境与农业气象试验站开展。在移动式遮雨棚下总共设置12个试验小区,每个试验小区的长宽为4 m×2 m,深度为3 m,四周及底部为水泥筑成,以防水分渗透;小区为原状土壤,田间持水量为22.7%,土壤容重为1.37 g·cm-3,凋萎湿度为5.0%,试验期间土壤水分全部来自灌溉。供试材料为当地冬小麦常规生产品种(郯麦98),于2017年10月6日播种,播种量为20 g·m-2,播种方式为条播,播种行距为25 cm,播种时施底肥75 g·m-2(磷酸二胺,总养分≥64.0%),播种到起身前按照大田常规管理,11月15日浇灌越冬水40 mm,控水试验时间为3月24日(起身期)到6月16日(成熟期),控水试验期间有雨雪时候将移动式遮雨棚遮挡所有小区,其余时间不进行遮挡。灌水方案如表1所示,分别在起身期(T1)、起身—拔节期(T2)、抽穗—灌浆期(T3)进行充分灌水,对照区(CK)是在起身期、拔节期、抽穗期、灌浆期共进行4次充分灌水,每次灌溉量均为100 mm,以上4个处理中每个处理重复3次。根据近30年地面气象观测资料,计算得到当地冬小麦起身、拔节、抽穗、灌浆期各发育期内的平均需水量分别是37.8、80.0、45.5和50.0 mm,而平均降水量分别为8.3、14.1、10.1和13.6 mm,不能满足冬小麦需水,因此灌溉对于华北平原冬小麦是很有必要的。灌溉量100 mm 是通过计算满足小麦需水量以及试验得来的,目的是保证0~100 cm土层的土壤含水量达到饱和,自此之后不再补充水分,保持一段时间无水分供应,模拟自然状态下土壤从水分饱和到产生干旱的整个过程。为反映水分胁迫真实情况,从3月23日至6月13日每隔5 d对每个试验小区测定0~50 cm每10 cm的土壤水分并进行平均,各处理的土壤相对湿度变化见图1,在每次水分处理之前的土壤相对湿度情况见表2。
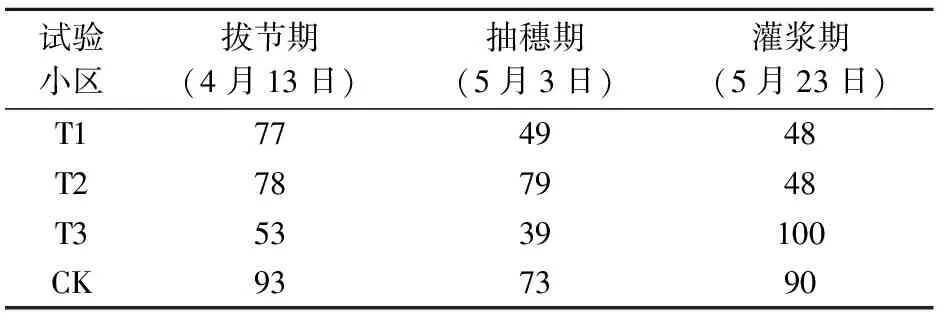
表2 冬小麦水分处理之前土壤相对湿度(单位:%)
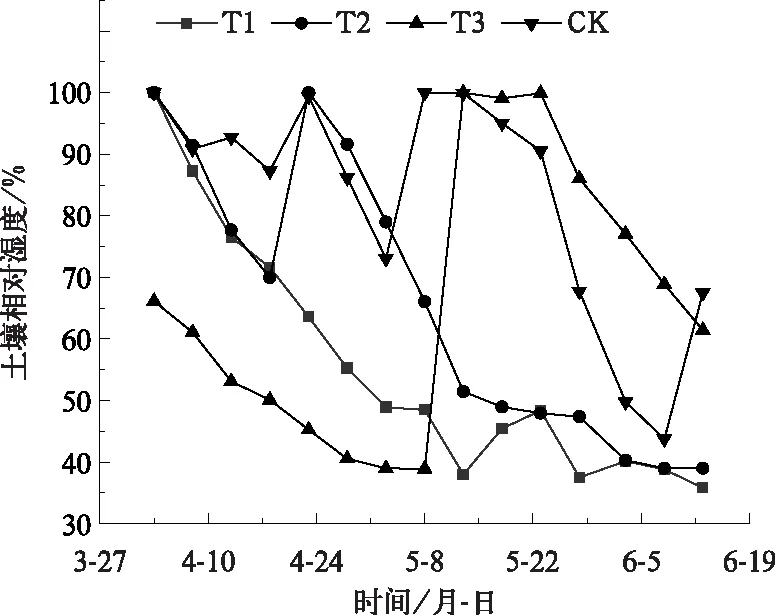
图1 不同水分胁迫处理的土壤相对湿度变化
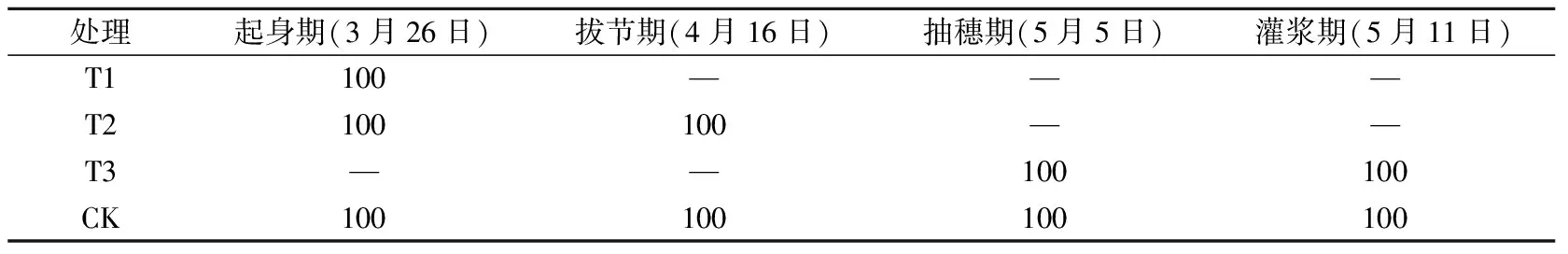
表1 冬小麦各生育期灌水处理(单位:mm)
1.2 测定项目与方法
1.2.1 灌水量
灌水使用智能流量积算仪(精度为0.000 00 m3)定量控制。
1.2.2 土壤湿度
采用土钻法人工取土,0~50 cm每10 cm取一层,烘干称重法计算土壤相对湿度:土壤相对湿度=土壤重量含水率/田间持水量。
1.2.3 旗叶光合参数
在冬小麦拔节期、抽穗期和灌浆期选择晴朗无风的天气,具体测定日期除灌浆期外,其他均为灌水处理前1天,即4月15日、5月4日、5月24日,于07—17时每2 h用便携式光合仪LI-6400测定冬小麦旗叶光合参数,包括净光合速率(Pn,单位:μmol·m-2·s-1)、蒸腾速率(Tr,单位:mmol·m-2·s-1)、气孔导度(Gs,mmol·m-2·s-1)、胞间CO2浓度(Ci,单位:μmol·mol-1)。为保证数据的代表性和有效性,每个试验小区连续测定5片旗叶。将4月15日(拔节期)、5月4日(抽穗期)和5月24日(灌浆期)每次光合测定日07—17时的6次光合参数测定值进行平均,分析不同生育期内光合特性变化。
1.2.4 叶绿素测定
在测光合参数的同日10—11时,用叶绿素仪(SPAD-502)测定旗叶叶绿素含量,每个试验小区连续测定5片旗叶,求其平均值。
1.2.5 产量及构成因素测定
在冬小麦成熟期,每试验小区连续取1 m2冬小麦植株,晒干后测定穗粒数、穗粒重和千粒重,人工收获试验区其余冬小麦进行脱粒、晒干、计产。
1.2.6 数据分析
采用Excel 2016和SPSS16.0 进行数据处理和统计分析,采用Duncan新复极差法(俞世蓉和郭蔼平,1981)进行显著性检验。
2 结果与分析
2.1 不同水分胁迫下冬小麦灌浆期光合参量的日变化特征
根据不同程度持续性水分胁迫造成的光合特性日变化特征,得出各处理冬小麦旗叶的光合特性日变化在灌浆期的差异最显著。由图2可见T1、T2和T3处理的Pn、Gs、Tr和Ci全天几乎低于CK。CK的旗叶Pn、Gs、Tr呈双峰型曲线,其他处理呈先升后降趋势,T1、T2和T3处理的Pn、Gs、Tr日变化曲线的峰值都出现在09时,较CK(第一个峰值在11时)明显提前。Duncan新复极差检验表明,不同水分处理的Pn、Gs、Tr与CK的差异均达到极显著水平(P<0.01),T1与T3处理之间差异不显著(P>0.05),但T1和T3处理的旗叶Pn、Gs、Tr比T2降低更明显,差异显著(P<0.05),说明拔节期之后出现持续性水分胁迫与灌浆期旗叶Pn、Gs、Tr的降低有很大关系,且随着水分胁迫程度的加重,旗叶Pn、Gs、Tr下降越明显,日变化幅度越小。这种差异可能与水分胁迫造成叶片光合的功能过早下降有关,这与肖颖(2002)的研究结果相吻合,即轻度水分胁迫对冬小麦旗叶的净光合速率影响较小,而中度以上胁迫虽在开花前保持较高的净光合速率,但开花后急降。
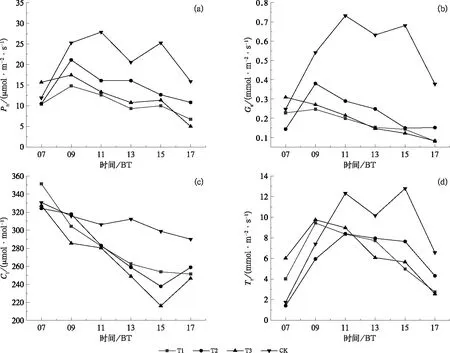
图2 冬小麦灌浆期(a)净光合速率、(b)气孔导度、(c)胞间CO2浓度、(d)蒸腾速率的日变化
2.2 持续性水分胁迫对旗叶叶绿素含量的影响
水分胁迫不仅抑制叶片中叶绿素的合成,也会加速已合成的叶绿素分解,导致叶片中叶绿素含量降低(薛崧等,1992),进而影响植物光合能力。如图3 所示,正常灌水条件下叶绿素含量在生育中后期保持在较高的水平。Duncan新复极差分析发现,拔节期各处理的叶绿素含量差异不显著(P=0.549),说明拔节期轻度水分胁迫对叶绿素含量影响不大。抽穗期各处理叶绿素含量仍处于较高水平,且T3处理的叶绿素含量高于仅在起身期灌水的T1。灌浆期各处理都明显低于CK,各处理间的差异极显著(P<0.01)。T1和T3的叶绿素含量比CK明显减少。

图3 冬小麦旗叶叶绿素含量随生育期的变化
2.3 持续性水分胁迫对旗叶光合特性的影响
2.3.1 净光合速率
从图4可看出,拔节期T3处理的旗叶Pn低于其他处理(P<0.05),说明拔节期轻度水分胁迫可降低旗叶Pn。抽穗期T1和T3处理的旗叶Pn比CK分别下降20.1%和15.1%,说明抽穗前持续性水分胁迫明显降低旗叶Pn,其中T1处理浇灌了起身水,而T3处理起身水和拔节水都未浇,因此T3水分胁迫时间更长,但T3的旗叶Pn大于T1,说明苗期轻度水分胁迫可增强冬小麦的抗旱性。灌浆期各处理旗叶Pn明显较CK降低,T1、T2和T3分别降低51.4%、32.1%和40.3%,随着水分胁迫时间越长,旗叶Pn下降越明显,其中T3处理的旗叶Pn小于T2,说明起身—拔节期比抽穗—灌浆期的持续性水分胁迫使灌浆期旗叶Pn下降更显著(P<0.05)。

图4 冬小麦旗叶净光合速率(Pn)随生育期的变化
2.3.2 气孔导度
气孔是植物叶片内外水汽和CO2交换的通道,既保证了光合作用的进行,也避免了体内水分的过度散失。Schurr et al(1992)的研究认为土壤干旱时,叶水势下降,膨压随之降低而引起气孔关闭。由图5可知,不同水分处理下冬小麦旗叶Gs与CK的差异显著,说明Gs对水分胁迫十分敏感(Davies and Zhang,1991;Schurr et al,1992)。拔节期T3的Gs比CK低20.9%,随着水分胁迫程度的增加,抽穗期T1和T3处理的旗叶Gs比CK分别下降38.8%和40.5%,差异极显著(P<0.01),而T1与T3处理之间的差异不显著(P=0.626)。灌浆期各处理旗叶Gs都比CK显著下降(P<0.01),T1和T3处理的旗叶Gs分别比CK降低57.7%和56.6%,说明前期持续性水分胁迫严重降低旗叶Gs,即使抽穗后复水也对Gs影响不大。
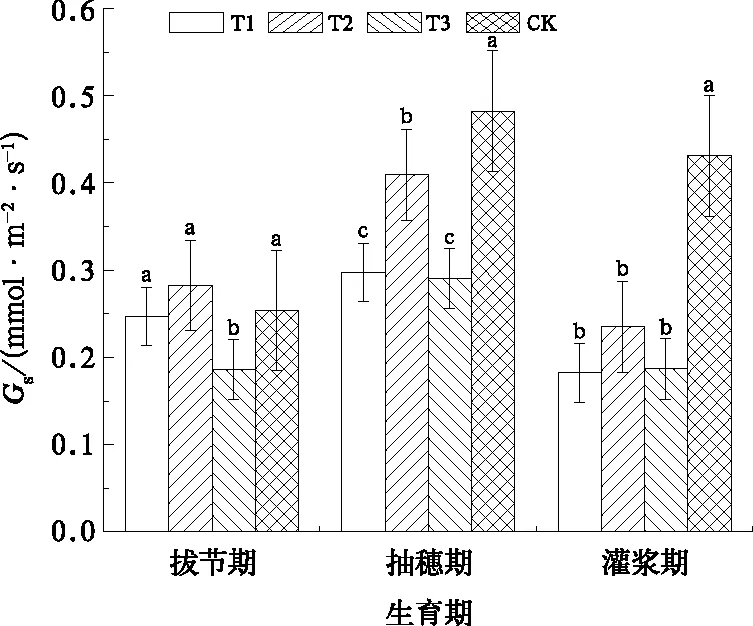
图5 冬小麦旗叶气孔导度(Gs)随生育期的变化
2.3.3 胞间CO2浓度
Ci的变化是分析植物气孔与非气孔限制的基础,是光合过程中CO2的中介,一方面受到作为源的外界CO2浓度和气孔导度的影响,另一方面又受叶片光合消耗的影响(傅伟和王天铎,1994)。由图6可见,拔节期T3受到轻度水分胁迫,Ci比CK明显降低;随着水分胁迫的持续,抽穗期T1和T3分别受到中度水分胁迫和重度水分胁迫,旗叶Ci比CK均有所降低,分别降低10.7%和16.1%。灌浆期T1、T2和T3的Ci比CK分别下降8.0%、9.4%和13.4%,与CK差异显著(P<0.05),但各处理之间差异不显著。
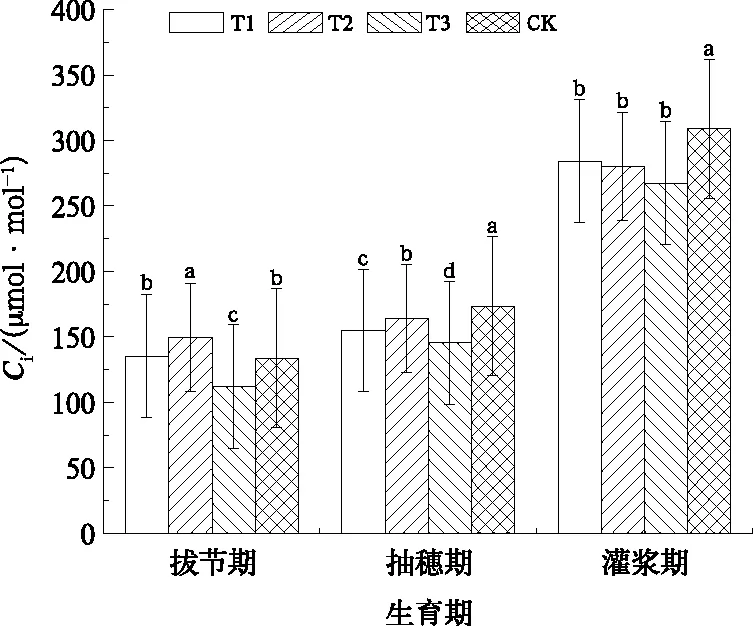
图6 冬小麦旗叶胞间CO2浓度(Ci)随生育期的变化
2.3.4 蒸腾速率
从不同灌水处理对旗叶Tr的影响来看(图7),拔节期T3的Tr明显低于CK,抽穗期各处理的Tr明显增大且差异减小,灌浆期各处理的Tr比CK明显下降,表明水分胁迫的累积效应到后期对叶片蒸腾速率的影响明显显现出来。除CK的Tr随着生育期推进而增大外,不同水分胁迫处理的Tr均为抽穗期最大,灌浆期减少,说明前期水分胁迫使灌浆期叶片早衰,功能下降,受到水分胁迫的植株蒸腾能力越差。CK的Tr在灌浆期迅速增大,但Pn基本没变化,说明冬小麦后期过多灌水反而造成水分散失,对净光合速率提高的促进作用不大。

图7 冬小麦蒸腾速率(Tr)随生育期的变化
2.4 水分胁迫对冬小麦产量及产量构成要素的影响
作物产量由单位面积穗数、穗粒数和千粒重决定,产量构成三要素是评价产量的重要指标。从各处理的籽粒产量看(表3),T1、T2和T3处理比CK处理分别减产50.8%、31.1%和57.0%。T1和T2千粒重比CK分别降低21.9%和14.9%,说明拔节或抽穗后持续性水分胁迫明显影响籽粒灌浆,且水分胁迫时间越长,对籽粒灌浆影响越大。在各处理中T3处理的产量最低,冬小麦穗数和穗粒数分别比CK降低30.6%和47.5%,说明起身到抽穗前持续性水分胁迫严重影响产量,即使后期充分灌水,也无法获得高产。

表3 2018年不同灌溉处理下冬小麦产量及产量构成要素
3 结论与讨论
水分是影响作物生长发育最重要的环境因子之一,水分胁迫不仅影响作物发育,更重要的是影响叶片光合作用和产量形成。光合作用是作物干物质形成的重要来源,持续性水分胁迫一般对作物光合产物形成有很大影响,因此其对作物光合特性和产量的影响特征是抗旱灌溉时机和次数选择的重要参考依据。冬小麦大田生产灌溉多是在春季进行一到两次充分灌溉,而较多的亏缺灌溉研究难有条件在大田应用,因此研究充分灌溉或透墒雨后水分胁迫的持续变化及对光合作用和产量的影响,对大田生产有更实际的参考作用。本试验结果表明,与水分充足处理对照(CK)相比,持续水分胁迫下冬小麦旗叶的叶绿素含量、净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)和胞间CO2浓度(Ci)均有所下降,蒸腾速率相对于净光合速率在灌浆期对水分胁迫的反应更敏感,与裴冬等(2006)、张雅倩等(2011)研究结论一致。在灌浆期各水分胁迫处理的光合日变化曲线中没有CK的明显双峰变化,而是呈单峰变化,且峰值较CK明显提前。在拔节期轻度水分胁迫下叶绿素变化不大,在抽穗期中到重度水分胁迫使叶绿素略有下降,灌浆期三种处理的旗叶叶绿素均明显下降。在起身期(T1)、起身—拔节期(T2)、抽穗—灌浆期(T3)三个阶段充分灌水处理的冬小麦Tr和Ci比CK差异显著,但三种处理之间在灌浆期差异不显著。T3即使后期复水,到灌浆期仍比T2的叶绿素含量、Pn和Gs降低更明显,与宁东峰等(2010)研究结果有一致性。蔡永萍等(2000)研究认为这是营养生长期严重的持续性水分胁迫造成灌浆期叶绿体损坏,使得叶肉细胞利用CO2能力降低所致。拔节期和抽穗期不同程度干旱均显著降低了冬小麦旗叶的净光合速率,而灌浆期灌水对旗叶净光合速率的影响较小,说明灌浆期复水对冬小麦旗叶净光合速率的补偿效应并不显著,可能是因为拔节期的干旱程度超出了水分调亏的合理范围(马富举等,2018)。
光合速率对冬小麦籽粒产量形成有极其重要的影响,尤其是抽穗后冠层叶片的光合速率对籽粒产量的贡献最大(张其德等,2000),本研究中持续性水分胁迫对产量的影响结果表明,起身和拔节期受到持续性水分胁迫的冬小麦,即使后期复水,产量仍明显低于CK,也低于前期充分灌溉而后期持续水分胁迫处理,从产量构成要素分析看出穗数和穗粒数明显减少,李全起等(2011)也得到相同结论。说明营养生长期随着水分胁迫持续时间延长至严重水分胁迫状态,会严重影响冬小麦的孕穗、抽穗,单位面积结实穗数和单株小穗数明显减少,导致严重减产。因此冬小麦起身—拔节期是水分胁迫影响产量最大的时段,起身—拔节期的充分灌溉或充足降水是实现高产的关键。
光合作用是一个很复杂的生理过程,水分胁迫下冬小麦的Pn、Tr、Gs、Ci等光合特性参数除了受水分影响外,还受很多其他环境因素影响,包括温度、大气湿度、外界CO2浓度、总辐射量和辐射强度等,其对冬小麦产量的影响需要进一步研究。
