夜间增温下施硅对稻田CH4好氧氧化及其氮响应的影响
2022-11-09周聪谢晴薛梦琪张耀鸿娄运生贾仲君高霄鹏
周聪,谢晴,2,薛梦琪,,张耀鸿*,娄运生,贾仲君,高霄鹏
(1南京信息工程大学气象灾害预报预警与评估协同创新中心/江苏省农业气象重点实验室,江苏南京 210044;2气象防灾减灾湖南省重点实验室,湖南长沙 410118;3中国科学院南京土壤研究所土壤与农业可持续发展国家重点实验室,江苏南京 210008;4曼尼托巴大学,加拿大温尼伯R3T2N2)
0引言
【研究意义】稻田是陆地甲烷(CH4)排放的重要源。据估算,稻田CH4排放约占全球CH4总排放量的17%(Schimel,2000)。稻田土壤中产生的CH4气体在进入大气之前大部分被氧化去除,是CH4消减的最主要过程(Cai and Mosier,2000)。其中,以好氧CH4氧化作用最重要,对稳定大气CH4含量及减缓全球温室效应具有重要意义。联合国政府间气候变化专门委员会第五次报告显示,近半个世纪全球平均地表温度持续变暖,近地气温增幅呈明显的昼夜不对称性,即夜间增幅显著高于白天(IPCC,2014)。夜间增温对稻田CH4氧化的影响受到人们普遍关注(刘燕等,2020;郭家宏等,2022)。硅是植物的有益元素之一,且是水稻的必需营养元素。叶片的硅化细胞对于散射光的透过量为绿色细胞的10倍,可使其吸收更多的光源,促进光合作用;还可促进碳水化合物的长距离转运,协调植物的库—源关系(高臣等,2011)。因此,研究硅肥对稻田CH4氧化过程及其温度敏感性的影响,对于缓解陆地生态系统典型湿地CH4排放及探索有效的减排措施具有重要意义。【前人研究进展】CH4排放通量是CH4产生过程和氧化过程的净综合结果。一般认为,增温强烈影响水稻的光合速率、光合碳分配、根系分泌及输导组织泌氧等生理过程,对根际土壤微环境(营养水平、酸碱度、氧化还原电位等)产生重要影响,进而对微生物驱动的CH4产生和氧化过程有显著影响效应(Voigt et al.,2017;Zhang et al.,2017;Li et al.,2020;Liu et al.,2020)。Zhou等(2018)研究认为,陆地湿地系统CH4排放量的土壤温度敏感系数(Q10值)介于1.7~28.0,强烈受CH4氧化作用贡献的影响。Wang等(2022)在双季稻田中进行田间开放式增温后发现,增温提高了水稻有效分蘖数和根系生物量,有利于O2从叶片运输到根际中,促进根际土壤的CH4氧化速率,而对CH4氧化菌功能基因pmoA的丰度无明显影响。Yan等(2022)研究发现,增温导致全球湿地系统CH4排放通量增加10.1%,而CH4氧化过程对此增幅的贡献尚不清楚。此外,钢渣硅肥具有提高水稻产量、提升土壤肥力的特性,在日本和韩国等地区已普遍施用(Das et al.,2019)。Lim等(2022)发现,施硅降低了水稻生长季CH4排放量的15%,其原因是钢渣硅肥提高了水稻根际的氧化还原电位,有利于CH4好氧氧化作用的进行;同时钢渣硅肥中含有丰富的Fe3+和Mn4+,可作为电子供体促进CH4厌氧氧化过程(Ali et al.,2008;Wang et al.,2015)。相反,有报道发现,钢渣硅肥促进了泥炭湿地CH4排放,原因是钢渣硅肥中释放出的P、Ca、Mg等元素促进了CH4产生菌的活性(Reithmaier et al.,2017;Hömberg et al.,2021)。Nakamura等(2022)研究发现,施硅对森林湿地土壤的CH4排放通量和产生过程均无影响,暗示施硅对CH4氧化过程也无明显影响。【本研究切入点】目前,有关增温或施硅单因子对稻田土壤CH4排放通量的影响已有报道,但二者耦合对稻田土壤CH4消减过程的影响尚不清楚;且以往的报道大多基于培养箱或玻璃温室进行增温,不能代表大田夜间增温的真实情况。因此,有必要研究增温/施硅对稻田CH4氧化过程的影响效应,阐明施硅条件下增温稻田CH4好氧氧化驱动的固碳特征变化规律。【拟解决的关键问题】采用田间开放式夜间增温系统提供长期增温环境,运用13CH4稳定性同位素标记法研究夜间增温/施硅对稻田土壤CH4好氧氧化过程及其有机碳累积的影响,为探索稻田温室气体减排措施提供科学依据和实践途径。
1 材料与方法
1.1 供试材料
田间试验在南京信息工程大学农业气象试验站进行。试验点属于亚热带湿润气候区,年均降水量1100 mm,年均气温15.6℃。土壤为潴育型水稻土,灰马肝土属,质地为壤质粘土。农作方式为当地典型的稻麦轮作制。供试氮、磷、钾肥料为当地普遍施用的高浓度复合肥(15-15-15)。供试硅肥为钢渣硅肥,取自南京钢铁集团有限公司,该粉状矿渣型硅肥有效硅(SiO2)含量为13.8%。
1.2 试验方法
试验站自2016年6月采用田间开放式增温系统设置长期试验一直持续至今。增温设2个水平,即夜间常温和夜间增温;施硅设2个水平,即不施硅和施硅(SiO2)200 kg/ha。试验共设4个处理,即夜间常温不施硅(CK)、夜间增温不施硅(NW)、夜间增温施硅(NW+Si)及夜间常温施硅(Si)。采用随机区组试验设计,每处理3次重复。小区面积4 m2(2 m×2 m)。在每处理区内采用根袋法,将根际土与非根际土用300目尼龙袋隔开。夜间增温采用被动式增温方式进行,即在试验小区四周搭设不锈钢管支架,将铝箔反光膜架设于支架上,在农作物(水稻/小麦)生长期内,夜间(19:00—6:00,随日长有所增减)用反光膜覆盖农作物冠层,白天将铝箔反光膜卷起。随农作物生长发育进程,及时调整支架高度,使反光膜与农作物冠层保持在0.3 m左右。为保持各试验小区植物受光度的一致性,在常温对照区内也搭设不锈钢管支架,而无反光膜覆盖。采用温度记录仪自动记录作物冠层和土壤5 cm处全生育期温度,数据记录间隔为30 min。前期试验结果表明,与常温对照区相比,水稻冠层和5 cm土层的全生育期平均夜温分别提高1.21和0.62℃,增温效果明显(郑泽华等,2017)。
2020年9月,分别从各处理稻田中采集0~10 cm耕层的根际土与非根际土。一部分鲜土用于测定各试验处理区耕层土壤的基本理化性质;另一部分鲜土用于培养试验,测定稻田土壤的CH4好氧氧化速率。其中,土壤有机碳(SOC)采用H2SO4消煮—FeSO4滴定法测定,可溶性有机碳(TOC)采用TOC分析仪测定,土壤总氮采用凯氏定氮法测定,铵氮(NH4+)和硝氮(NO3-)采用KCl提取—流动分析仪测定,土壤有效磷采用钼蓝比色法测定,土壤速效钾采用火焰光度计测定(Gao et al.,2022)。
1.3 土壤CH4氧化速率及13C有机碳分析
称取2.000 g鲜土放入12 mL的Labco顶空瓶,加入2 mL去离子水后盖好盖子,充分摇晃30 s成均匀泥浆,25℃预培养1 d。然后培养瓶中抽真空—充氩气3次,再置换等量的4 mL高纯O2和1 mL13CH4后进行正式培养(胡敏杰等,2017)。每个瓶子的顶空空间在培养开始时含有约10%的CH4浓度,25℃避光培养。同时,进行3种不同形态氮添加处理,分别为:NH4+-N、NO3--N和尿素(Urea),使瓶中最终N浓度为5 mmol/L。每个稻田土壤样品共计4个培养添加处理,每处理3次重复。采用气相色谱仪(Agilent 7890B)测定0时刻CH4和CO2起始浓度。之后分别在第3、6、9和12 d采气测定培养瓶顶空气体的CH4和CO2浓度。根据CH4浓度随时间的变化计算CH4氧化速率(R)。
式中,C0为零时刻顶空气体中的CH4浓度(μL/L),Ct为培养t时刻后的CH4浓度(μL/L),V为培养瓶体积(L),ρ为标准状态下CH4密度(g/L),t为培养时间(d),m为土壤干土质量(g)。
培养结束后,瓶内加入2 mol/L的HCl溶液进行土壤酸化处理(去除无机碳),然后将土壤泥浆冷冻干燥后测定有机碳含量及其13C有机碳(13C-SOC)丰度值和净增量。
1.4 统计分析
试验数据采用Excel 2016和SPSS 21.0进行整理分析,并对处理间土壤理化性质、CH4氧化速率及其固碳量进行方差分析和多重比较,对CH4氧化速率与土壤理化性质进行相关分析。利用Origin 2021制图。
2 结果与分析
2.1 长期试验处理下稻田土壤的理化性质变化
经过4年多的长期增温/施硅处理后,各试验小区耕层土壤的理化性质发生明显变化。由表1可知,Si处理的稻田耕层土壤有机碳和可溶性有机碳含量分别较CK显著提高10.5%和26.7%(P<0.05,下同);而NW处理的这2个指标则分别较CK显著降低11.9%和9.9%。NW+Si处理的稻田可溶性有机碳含量较CK显著提高16.4%,而有机碳和总氮含量均显著低于CK;Si处理的土壤总氮、氨氮、硝氮含量均显著高于其他处理,而NW处理的氮含量均为最低。施加硅肥处理(Si和NW+Si)的有效磷和速效钾含量普遍高于未施硅肥处理(CK和NW),且Si处理的土壤有效磷和速效钾含量显著高于NW处理。以上结果表明,施加硅肥有利于提升耕层土壤的有机碳、活性碳及磷和钾含量。
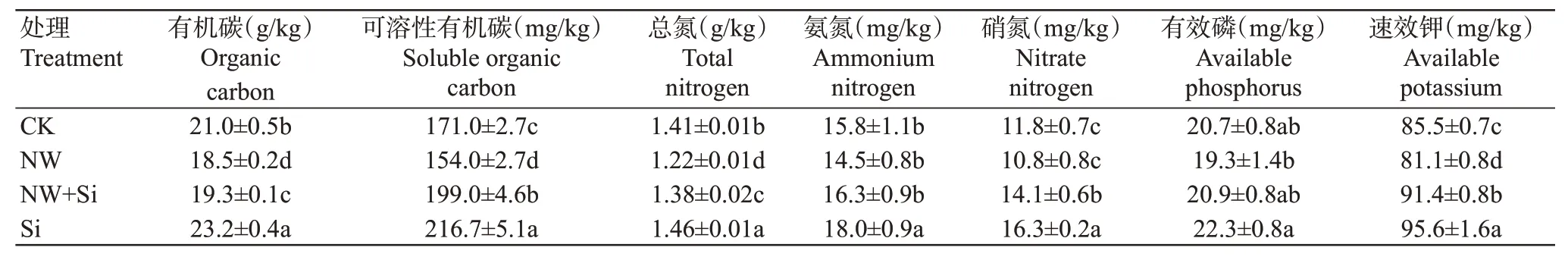
表1 长期试验处理下稻田土壤的理化性质Table 1 Soil physicochemical properties of paddy fields under long-term experimental treatments
2.2 不同处理的稻田土壤CH4好氧氧化速率
由培养期间起始和终止时刻顶空气体CH4浓度变化可计算得到土壤CH4好氧氧化速率。由图1可知,稻田非根际土的CH4好氧氧化速率明显高于根际土,且4个处理间无显著差异(P>0.05,下同),其数值介于16.0~17.0μg/(g·d)。对于根际土而言,4个处理的稻田CH4好氧氧化速率存在显著差异,施硅处理(Si和NW+Si)的氧化速率显著高于未施硅处理(CK和NW),其中Si处理的土壤CH4好氧氧化速率最高,为10.9μg/(g·d),而NW处理则最低,为7.5μg/(g·d)。
将根际土和非根际土的CH4氧化速率分别与土壤(包括根际土和非根际土)理化性质进行相关分析,发现根际土CH4氧化速率与土壤有机碳和总氮含量呈显著正相关,相关系数分别为0.71和0.74;与水溶性有机碳、矿质态氮、有效磷和速效钾含量均呈极显著正相关(P<0.01),相关系数在0.83~0.89。表明增温/施硅处理后,土壤理化性质的变化可能是稻田根际土壤CH4氧化速率产生差异的重要原因。相反,非根际土CH4氧化速率与土壤理化性质之间未表现出显著相关性。
2.3 不同处理土壤的13C-SOC丰度值与净增量
测定CH4氧化速率的培养试验结束后,将培养瓶内的泥浆酸化处理后冷冻干燥,分析其有机碳含量及13C丰度值,进而计算由底物13CH4被氧化并同化进入到土壤中的13C-SOC净增量。由图2可知,所有供试土壤的13C-SOC丰度值均远高于其自然丰度值(1.08%),表明培养瓶内进行了明显的13CH4好氧氧化过程,且有相当一部分底物13CH4被同化进入了土壤中。其中,在非根际土中,施硅处理(Si和NW+Si)的13C-SOC丰度值显著高于未施硅处理(CK和NW),Si稻田土壤13C-SOC丰度值最高,达3.45%,而NW处理则最低,为3.28%。根际土也表现出与非根际土类似的变化规律,且其13C-SOC丰度值总体均明显低于非根际土。
由图2还可看出,在底物13CH4好氧氧化过程中,有一部分13CH4被同化到土壤有机碳中。对于根际土而言,Si处理的土壤13C-SOC净增量为13.6μmol/g,显著高于其他处理。对于非根际土而言,施硅处理(Si和NW+Si)的13C-SOC净增量显著高于未施硅处理(CK和NW),其中NW+Si处理最高,达13.2μmol/g,而CK最低。总体而言,根际土的13C-SOC净增量小于非根际土(Si稻田除外)。
将土壤的13C-SOC净增量与13CH4净消耗量相比,可反映出稻田土壤CH4好氧氧化过程的固碳效率。本研究中发现,施硅处理(Si和NW+Si)的CH4好氧氧化固碳效率为61%~66%,明显高于CK和NW处理的固碳效率(51%~54%),表明施硅有利于CH4好氧氧化过程中有较多的CH4碳源被固定到土壤中,促进土壤有机碳的累积。
2.4 增温/施硅稻田CH4氧化速率对氮输入的响应特征
在测定CH4氧化速率的培养试验中,分别添加NH4+-N、NO3--N和Urea进行培养,以模拟增温/施硅稻田在田间施加化学氮肥情景下CH4氧化过程的变化特征。由图3可知,在添加3种氮形态条件下,4个处理非根际土中的CH4氧化速率均明显高于根际土(NO3--N的Si处理除外),这与无氮添加下的氧化速率特征一致,表明施加氮肥对根际土与非根际土之间的CH4氧化速率差异无明显影响作用。对于非根际土而言,添加NH4+-N、NO3--N或Urea条件下,4个处理的CH4氧化速率差异明显,且均以NW+Si处理的氧化速率最高,达33.6~36.8μg/(g·d)。相对于无氮添加处理,添加NH4+-N和Urea均明显提高了4个处理土壤的CH4氧化速率,且以添加NH4+-N效果更明显。对于根际土而言,添加NH4+-N条件下,CK的CH4氧化速率显著高于NW和Si处理,而NW+Si处理则最低。添加NO3--N条件下,Si处理的CH4氧化速率显著高于其他处理,与无氮添加条件下的速率特征一致。添加Urea条件下,NW+Si处理的CH4氧化速率为18.2 μg/(g·d),显著高于其他处理,而NW稻田则最低。
综合来看,与无氮添加相比,添加氮条件下4个处理的根际土和非根际土CH4氧化速率总体有大幅提升,说明施加氮肥对增温/施硅稻田的CH4好氧氧化具有促进作用。在3种氮添加条件下,NW+Si处理非根际土的CH4氧化速率均显著高于其他处理,且相对无氮添加处理其增幅也最大,说明NW+Si处理非根际土CH4好氧氧化对外源氮输入的响应度最高。
2.5 CH4氧化过程驱动的13C-SOC净增量对氮输入的响应
由图4可知,在添加3种氮形态条件下,4个处理非根际土中的13C-SOC净增量均明显高于根际土(Si处理除外),与无氮添加下的净增量特征一致,表明施加氮肥对根际土与非根际土的13C-SOC净增量差异无明显影响。对于根际土而言,在添加3种形态氮条件下,Si处理土壤CH4氧化过程驱动的13C-SOC净增量显著高于其他处理,与无氮添加处理的变化特征一致。对于非根际土而言,施硅处理(Si和NW+Si)的13C-SOC净增量显著高于未施硅处理(CK和NW),其中NW+Si处理最高,达13.3μmol/g,与不添加氮条件下的净增量基本持平。总体而言,与不添加氮处理相比,添加3种氮条件下4个处理的土壤CH4氧化过程介导的13C-SOC净增量绝对值变化幅度很小,说明施加氮肥对增温/施硅稻田CH4氧化驱动的土壤固碳量无明显影响。此外,添加3种氮条件下,施硅处理(Si和NW+Si)的CH4好氧氧化固碳效率(48%~55%)明显高于CK和NW处理的固碳效率(38%~51%),与无氮添加下固碳效率的规律一致,但施加氮肥降低了增温/施硅稻田土壤CH4氧化驱动的固碳效率。
3 讨论
CH4好氧氧化作用是稻田土壤CH4消减的主要过程,其由好氧甲烷氧化菌在有O2参与条件下催化完成,该微生物以CH4为唯一碳源和能源进行生长。首先甲烷好氧氧化菌利用其所含的甲烷单加氢酶(MMO)将CH4氧化为甲醇,然后在甲醇脱氢酶(MDH)作用下氧化为甲醛,甲醛通过RuMP循环或Serine循环合成细胞物质,或甲醛被进一步氧化为甲酸,最终降解为CO2。由此可见,CH4好氧氧化过程的碳源流向有两个:一方面CH4碳源被同化到土壤微生物中,提高了土壤微生物碳含量,另一方面微生物好氧呼吸将其异化为CO2。本研究中将土壤的13CSOC净增量与13CH4净消耗量相比,计算出试验稻田CH4好氧氧化的固碳效率为51%~66%。与Trimmer等(2015)报道发现英国Chalk河流沉积物的CH4好氧氧化平均固碳效率约为50%的结果相似。然而,有报道发现,在CH4浓度较高的覆盖土中CH4好氧氧化的固碳效率可达69%,而在CH4浓度较低的森林土壤中其固碳效率仅为31%~43%(Roslev et al.,1997)。从功能菌代谢途径来看,以RuMP途径进行碳代谢的II型CH4好氧菌倾向于将较高比例的C-CH4转化到微生物体内,显著高于以Serine途径的I型CH4好氧菌的固碳效率(蔡朝阳等,2016)。可见,稻田中的CH4好氧氧化过程不仅可将CH4转化为增温潜势较小的CO2气体,更重要的是可将一半以上的甲烷碳源同化进入土壤微生物中,促进土壤有机碳累积。
本研究发现,4个处理中非根际土的CH4好氧氧化速率均显著高于根际土,可能与根际微环境中的营养养分水平有关。已有研究发现,水稻光合碳总量的7.6%~23.5%以有机酸(苹果酸、柠檬酸、琥珀酸等)的形式分泌到根际微域中,为微生物生长提供了一个富碳营养区(Wei et al.,2019)。CH4好氧氧化菌具有底物代谢多样性,可以苹果酸、柠檬酸、琥珀酸等作为碳源进行合成代谢(Chen et al.,2019)。因此,根际土壤中丰富的有机酸对CH4好氧氧化菌利用CH4分子产生底物竞争,使得本研究中通过CH4浓度变化计算出的CH4好氧氧化速率可能被低估。不仅如此,水稻根系从根际土壤中强烈吸收营养元素磷和钾等,通常造成根际土壤矿质营养元素的普遍匮乏。新近的研究发现,磷缺乏显著降低水稻根际土壤CH4好氧氧化速率及其相关微生物功能基因pmoA的总丰度,改变了I型和II型CH4好氧氧化菌的相对丰度(Gao et al.,2022)。这种效应在NW处理中表现最明显,即在无氮添加条件下非根际土的CH4氧化速率比根际土高出1.27倍,在添加尿素条件下也表现出相同特征。夜间增温可能刺激了水稻根系分泌较多的有机酸,抑制了CH4氧化菌对CH4分子的利用;同时夜间增温有利于水稻根系生长,促进根系对根际土壤营养元素的吸收,从而对CH4氧化菌的矿质营养元素利用造成不利影响。由此可见,水稻根系生长对根际微环境的养分状态产生重要影响,在此生境中CH4好氧氧化过程对环境条件变化(如增温/施硅)表现出较高的响应性。因此,在研究其他环境要素对CH4好氧氧化过程的影响效应时,选用根际土进行分析可能会得出更科学可靠的结论。
CH4好氧氧化作用是微生物驱动的生化过程,凡是影响微生物活性或代谢的环境因素均会对CH4好氧氧化速率产生影响。其中,温度是CH4好氧氧化过程的重要因素之一(郭家宏等,2022)。本研究发现,无氮添加条件下夜间增温抑制了稻田根际土CH4好氧氧化速率,且该速率与土壤有机碳、总氮和活性氮含量等均呈显著正相关。有报道认为,夜间增温会降低水稻根系活力,减少进入土壤的有机碳量,使得根际土壤中甲烷氧化菌所需养分减少,进而降低甲烷氧化菌活性(Zhang et al.,2020)。而且,稻田夜间温度敏感性系数Q10相对于白天更大,说明在夜间增温后微生物活性受温度影响更大(贾庆宇等,2020)。因此,增温导致土壤理化性质发生改变,进而影响CH4好氧氧化微生物的活性可能是本研究夜间增温抑制CH4好氧氧化的主要原因之一。不仅如此,温度升高会使得土壤中其他微生物活性增强和有机物分解速率增加,导致土壤中O2消耗量增大,CO2浓度上升,养分资源匮乏,诱导底物性竞争,最终抑制CH4的好氧氧化(Zhang et al.,2020)。
硅是水稻生长的必需营养元素,有关硅对稻田CH4好氧氧化过程及其功能微生物影响的报道十分鲜见。一般认为,施加硅肥促进水稻叶片的硅化细胞发育,提高光合速率,可将较多的光合产物以根际分泌物方式释放到根际土壤中(张万洋和李小坤,2020);且对水稻根系生长也有促进作用,可增加水稻根部泌氧能力(Ma,2001)。这将极大改善根际土壤的微生物生长环境,有利于CH4好氧氧化菌活性的提升。本研究发现,施硅稻田根际土壤的CH4好氧氧化速率均高于不施硅稻田,以Si处理最高。此外,施硅与增温处理对CH4好氧氧化速率的影响存在显著的交互作用,表明硅肥对稻田根际土壤CH4好氧氧化过程的促进效应会受到夜间温度变化的强烈影响。尽管如此,NW+Si处理根际的CH4好氧氧化速率显著高于NW处理,表明在夜间增温条件下施硅对CH4好氧氧化速率仍具有促进效应;说明在未来增温背景下施加硅肥可能是促进CH4氧化、缓解CH4排放的有效途径之一。刘燕等(2020)的田间试验结果也表明,在夜间增温条件下,施硅处理的水稻全生育期CH4累计排放量下降39%,说明其CH4好氧氧化速率的提升是全生育期CH4累计排放量下降的主要原因。另一方面,本研究中,Si处理土壤的13C-SOC净增量显著高于其他处理,且施加硅肥处理的CH4好氧氧化固碳效率明显高于不施硅处理。暗示施硅可能改变了土壤中CH4氧化菌的群落结构,更有利于II型CH4好氧菌的生长繁殖,促使较多的CH4碳源被固定到土壤微生物碳中,促进土壤有机碳的累积。
外源氮对CH4好氧氧化过程具有重要影响。本研究中,总体来看,与无氮添加相比,添加3种不同形态氮条件下稻田土壤CH4好氧氧化速率大幅增加,尤其以非根际土更明显。说明施加氮肥可促进稻田的CH4好氧氧化作用。其原因可能是外源氮缓解了微生物细胞生长对氮素的限制,促进了好氧甲烷氧化菌落的活性及相关反应的酶合成。然而有研究表明,由于氮输入的形态、浓度、施肥方法等均会对CH4好氧氧化造成不同影响,所以氮对CH4好氧氧化的研究结果并不一致(Ravishankara et al.,2009;Ettwing et al.,2016)。本研究中,外源氮对稻田CH4好氧氧化速率起促进效应的总体表现为NH4+-N>Urea>NO3--N。有研究认为,NH4+-N可与微生物的CH4单加氧酶结合,从而抑制CH4氧化过程(Schimel,2000)。Cai和Mosier(2000)的研究却发现NH4+-N对CH4氧化的抑制作用随CH4浓度的增加而减弱,最终表现为促进CH4氧化。本研究中,CH4底物浓度超过10%,其较高的浓度可能导致外源氮促进CH4氧化过程。在不同处理中,NW+Si处理非根际土对外源氮添加的响应度最高,表明夜间增温背景下,通过合理配施氮肥和硅肥可能是提高稻田CH4氧化速率并异化为CO2的一条有效途径。值得注意的是,外源氮对增温/施硅稻田CH4氧化驱动的固碳净增量无明显影响,导致固碳效率有所下降。这可能是因为外源氮提高了CH4氧化微生物的代谢活性,而未促进微生物繁殖生长所致。所以,综合来看,施加氮条件下,增温/施硅稻田CH4氧化为CO2的潜势大幅提高,而同化到土壤中的13C-SOC固碳量并无明显变化。
4 结论
长期夜间增温抑制了稻田根际土的CH4好氧氧化速率,同时降低其驱动的固碳效率。施硅可缓解夜间增温对稻田CH4氧化过程的抑制作用,并可提高CH4氧化过程的固碳潜力。施加氮肥可大幅提高稻田CH4好氧氧化速率,而其驱动的13C-SOC固碳量无明显变化。
