不同分离方法对红托竹荪菌种的复壮效果
2022-11-08黄晓润黄万兵刘倾城卢颖颖朱国胜
黄晓润,黄万兵*,刘倾城,卢颖颖,朱国胜,桂 阳
(1.贵州省农作物品种资源研究所,贵州 贵阳 550006;2.贵州省食用菌育种重点实验室,贵州 贵阳 550006;3.贵州省食用菌现代农业产业技术体系资源育种功能实验室,贵州 贵阳 550006;4.贵州省食用菌工程技术研究中心资源育种实验室,贵州 贵阳 550006)
红托竹荪(Dictyophora rubrovolvata)目前主要分布于我国贵州、云南、四川等西南地区,其营养丰富,味道鲜美,是贵州省的特色珍稀食用菌,具较高的营养和经济价值[1-4]。菌种退化是食用菌生产中存在的普遍问题[5-6],菌种退化势必会影响菌种质量与产量。菌种退化分为可逆和不可逆两类,其中,可逆的退化可通过复壮恢复优良特性,通常采用的复壮方法有菌丝尖端分离法、子实体组织分离法和原生质体再生法等[7-8]。红托竹荪在栽培过程中常出现菌丝退化、菌种老化,出菇延迟,导致产量严重下降。为避免红托竹荪优良性状的退化,以保存1年的红托竹荪菌株为主要试验材料,利用菌丝尖端分离法、竹蛋组织分离法和子实体组织分离法进行复壮,以期筛选出红托竹荪复壮的最佳方法。
1 材料与方法
1.1 试验材料
1.1.1 供试菌株 保存1年的红托竹荪菌株,编号分别为2182、yzs020、1907,由贵州省农作物品种资源研究所真菌研究室提供。
1.1.2 培养基 木屑培养基配方为葡萄糖5 g、果糖25 g、蛋白胨2.5 g、硫酸铵2.5 g、维生素B60.16 g、琼脂8 g、木屑70 g(60目筛)、土豆100 g、玉米粉20 g、蒸馏水1 000 mL,pH自然。
1.2 试验方法
1.2.1 尖端分离法 采用菌丝尖端分离法,将红托竹荪2182、yzs020、1907 接种于木屑培养基平板,长到2 cm 后挑取尖端部分1 cm的菌丝再次接种于木屑培养基平板;对照(CK)为保藏种直接接种于木屑培养基平板。
1.2.2 竹蛋组织分离法 将种植的红托竹荪竹蛋用75%酒精擦洗表面,在无菌室超净工作台紫外灯照射10 min 后进行组织分离,接种于木屑培养基平板,挑取菌盖、菌裙、菌柄、胶质层、菌托膜5个部位的组织,接入木屑培养基平板培养观察[9]。
1.2.3 子实体组织分离法 将种植的红托竹荪子实体用75%酒精擦洗表面,在无菌室超净工作台紫外灯照射10 min 后进行组织分离,接种于木屑培养基平板,挑取菌盖、菌裙、菌柄、胶质层、菌托膜5个部位的组织,接入木屑培养基平板培养观察[9]。
1.3 指标测定
挑取约0.5 mm×0.5 mm 的组织块,置于培养基上22℃培养。每组分离方法及CK均设5 个平板重复,每天定时观测,记录萌发时间、菌丝生长速度、菌丝长势、萌发率及污染率。
2 结果与分析
2.1 红托竹荪尖端分离菌丝体的生长情况
由表1可知,CK 菌丝长势稀疏;尖端分离后2182、yzs020、1907 菌丝体的长势强于CK,均为生长浓密。生长速度CK 最低,为0.96 mm/d;尖端分离后的菌株均高于CK,其中,yzs020 生长最快,为1.19 mm/d。尖端分离后菌丝体萌发率、萌发时间、污染率一致,分别为100%、24 h、0%。

表1 红托竹荪尖端分离菌丝体的生长情况
2.2 红托竹荪竹蛋组织分离菌株的生长情况
由表2 可知,接种菌盖组织除2182 菌株全部污染(污染率达100%)外,1907 菌株、yzs020菌株的菌丝长势、生长速度相同,均为菌丝生长密、0.21 mm/d。1907 菌株污染率为60%,yzs020 菌株全萌发。菌株萌发时间yzs020为48 h,1907菌株为24 h。
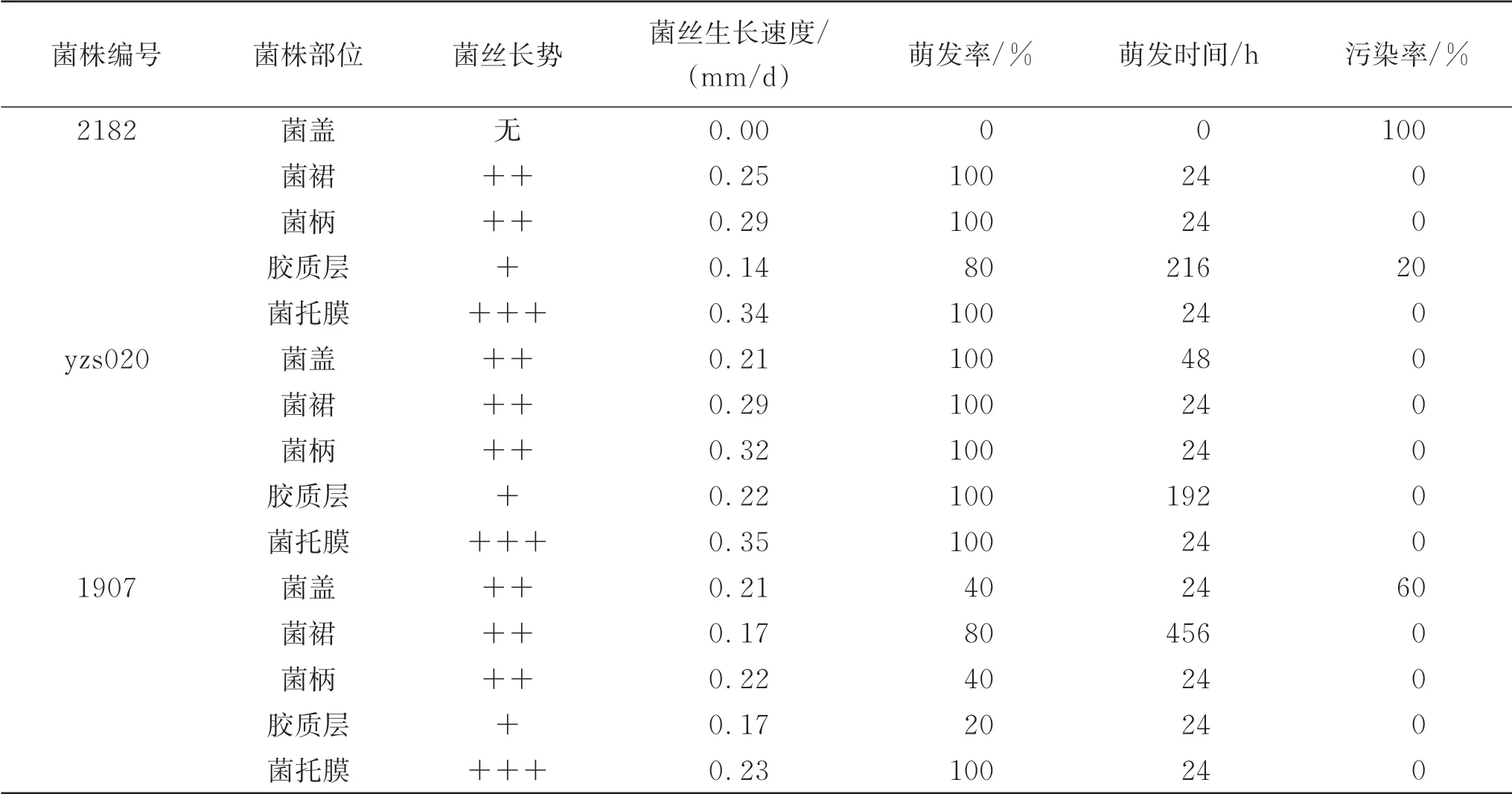
表2 红托竹荪竹蛋组织分离菌株的生长情况
菌裙组织2182 菌株、yzs020 菌株、1907菌株菌丝长势均为生长密,菌丝生长速度分别为0.25 mm/d、0.29 mm/d、0.17 mm/d。2182 菌株、yzs020 菌株全萌发,1907 菌株萌发80%。2182 菌株、yzs020 菌株萌发时间均为24 h,1907菌株萌发时间较慢,为456 h。
菌柄组织2182菌株、yzs020菌株、1907菌株菌丝长势均为生长密,菌丝生长速度分分别为0.29 mm/d、0.32 mm/d、0.22 mm/d。2182菌株、yzs020菌株全萌发,1907菌株萌发40%。萌发时间均为24 h。
胶质层组织2182 菌株、yzs020 菌株、1907 菌株菌丝长势均为生长稀疏,菌丝生长速度分别为0.14 mm/d、0.22 mm/d、0.17 mm/d。2182菌株萌发80%,yzs020菌株全萌发,1907 菌株仅萌发20%。萌发较慢,2182 菌株、yzs020 菌株萌发时间分别为216 h、192 h,1907菌株为24 h。2182菌株污染率为20%。
菌托膜组织2182 菌株、yzs020 菌株、1907 菌株菌丝长势均为生长浓密,菌丝生长速度分别为0.34 mm/d、0.35 mm/d、0.23 mm/d。3个菌株全萌发,萌发时间均为24 h。
2.3 红托竹荪子实体分离菌株的生长情况
由表3 可知,菌盖组织2182 菌株、yzs020菌株、1907菌株全部污染(污染率达100%)。

表3 红托竹荪子实体分离菌株的生长情况
菌裙组织除1907 菌株未萌发外,2182 菌株、yzs020 菌株的菌丝长势均为生长密,生长速度分别为0.22 mm/d、0.20 mm/d,萌发率、萌发时间均分别为20%、24 h。2182菌株污染率为80%。
菌柄组织除1907菌株全部污染(污染率达100%)外,2182菌株、yzs020菌株的菌丝长势均为生长密,生长速度分别为0.24 mm/d、0.22 mm/d。2182菌株、yzs020菌株萌发率分别为40%、100%,萌发时间均为24 h。2182菌株污染率为60%。
胶质层组织2182 菌株全部污染(污染率达100%)、yzs020 菌株未萌发。1907 菌株菌丝长势为生长稀疏,生长速度0.21 mm/d,萌发率为20%,萌发时间72 h,污染率为80%。
菌托膜组织2182 菌株、yzs020 菌株、1907 菌株菌丝长势均为生长浓密,菌丝生长速度分别为0.34 mm/d、0.35 mm/d、0.35 mm/d。萌发率yzs020 菌株为100%,2182 菌株、1907 菌株为80%。萌发时间均为24 h。2182 菌株、1907 菌株污染率均为20%。
3 结论与讨论
食用菌菌种的衰老主要与自身遗传变异和环境因素相关。自身遗传主要包括变异和基因突变等;环境因素主要包括培养条件不适宜、菌种保藏环境不适合、菌种保藏不当和菌株被病毒感染等[10-11]。针对红托竹荪及其他食用菌菌种老化、衰老最主要的预防措施,其基本原则就是“预防为主”,减少环境因素带来的影响,保证菌种的质量,因此不能考虑单一因素,需要从多方面进行预防。对退化菌种,可采取定期查看剔除退化菌种、定期分离纯化、创造适合的培养基质及环境条件、控制菌种的传代次数等措施入手[12]。
采用尖端分离技术分离时,3 株复壮菌株长势为生长浓密;生长速度为1.09~1.15 mm/d,复壮菌株生长速度明显高于保存1年的对照菌株(0.96 mm/d);萌发率、萌发时间、污染率一致,分别为100%、24 h、0%。尖端分离的菌丝体进行复壮效果最好,这可能与尖端分离后菌丝体内潜伏或感染的杂菌显著下降,菌丝体对培养料的分解力、适应性增强有关[13-15]。但由于多次尖端分离之后,可能会造成菌丝体发生变异等情况,因此该种方法可用于短期内的菌株复壮,食用菌的传代次数一般不得超过5次[16]。
红托竹荪竹蛋组织分离复壮时,菌托膜部位分离的萌发率最高,3 个菌株全萌发,萌发时间较快,均为24 h;菌丝生长速率较快,2182 菌 株、yzs020 菌株和1907 菌株 分别为0.34 mm/d、0.35 mm/d 和0.23 mm/d,且没有污染,但由于竹蛋未形成完整的子实体,无法知晓其农艺性状的优势,所以通过竹蛋的菌托膜复壮可能不是较佳的菌株。
红托竹荪子实体进行组织分离复壮时,其菌盖、菌裙、菌柄因暴露在外,导致其被污染的概率大大增加,通过菌落形态特征初步判断常被霉菌、枯草芽孢杆菌等菌污染,菌托膜和胶质层因有部分包裹在内,所以污染的概率相对较小,菌托膜部位分离的萌发率相对较高,yzs020 菌株全萌发,2182 菌株和1907 菌株萌发率均为80%;污染率也较低,仅20%;萌发时间24 h;菌丝生长速度较快,为0.34~0.35 mm/d。
在红托竹荪菌株复壮中,目前采用的方法主要有菌丝尖端分离法和组织分离法,研究只对复壮菌株菌丝的一些生长指标进行测定,对于红托竹荪的尖端分离最佳次数,尖端分离和组织分离复壮之后的农艺性状特征还有待进一步深入研究。
