盐胁迫对薄壳山核桃幼苗光合作用及叶绿体超微结构的影响
2022-11-07章建红沈登锋洪春桃焦云魏斌潘存德
章建红, 沈登锋, 洪春桃, 焦云, 魏斌, 潘存德
(1.宁波市农业科学研究院 林业研究所,浙江 宁波 315040; 2.新疆农业大学 林学与园艺学院,新疆 乌鲁木齐 830052)
光合作用是所有植物生存生长的基础。高盐环境会影响植物光合作用的正常进行,主要表现为气孔抑制与非气孔因素抑制,一般表现为植物在盐胁迫环境中的叶片胞间二氧化碳浓度升高,并通过调节气孔开度以降低其CO2浓度。槲栎在盐胁迫环境中光合作用受限,主要与气孔抑制有关[1];枸杞受到盐胁迫后,短期内光合作用受限是由于气孔因素导致,长期盐胁迫致使光合作用能力下降则为非气孔因素所导致[2];这一结果与对海滨锦葵的研究结果一致[3];张会慧等[4]发现,桑树在盐胁迫下其叶黄素循环受到抑制,PSⅡ反应中心正常功能受影响导致光合作用的下降。叶绿素荧光动力学研究表明,盐环境下植物的电子传递效率[5]和PSⅡ活性[6]同样受到影响。然而,不同植物或相同植物在不同生理时期和试验环境下的光合作用受影响的程度有一定差异。薄壳山核桃原产于北美,目前在我国广泛种植[7],但薄壳山核桃Pawnee在盐胁迫下的抗性表现还不是很清楚。本研究以一年生薄壳山核桃Pawnee幼苗为试验材料,采用水培法比较不同浓度的NaCl对其叶绿素含量、光合特性及叶绿体超微结构的影响,探讨薄壳山核桃Pawnee耐盐性的生理机制,以期为健全薄壳山核桃滨海地区盐碱地栽培技术提供理论支持。
1 材料与方法
1.1 供试材料
以一年生薄壳山核桃品种Pawnee容器苗为供试材料,试验于2019年4—11月在宁波市农业科学研究院农业高新技术园区温室大棚内进行。
1.2 处理设计
配置不同浓度水平的NaCl溶液(0、0.3%、0.6%、0.9%)对一年生薄壳山核桃Pawnee的容器苗进行盐胁迫处理,视干湿情况每隔2~5 d浇1次NaCl溶液,每次浇NaCl溶液定量为1.5倍容器的容量,重复3次,每处理含30株幼苗,按0、1、10、30、45、75、105 d处理时间点采集叶片样品用于后续检测分析。
1.3 观察和测定
1.3.1 叶绿素含量
每处理组分别从5个以上植株随机剪取1张叶片(顶部开始往下第三轮全功能叶)作为1个样品,重复3次。将采回的新鲜叶片剔除中脉剪碎混匀,称取0.2 g样品浸泡于5 mL 95%乙醇中,室温暗环境放置48 h后(至叶片完全发白),对样品进行短暂离心,弃去管底部的叶片残渣,保留上清液。用紫外可见分光光度计(UV-1800PC,Mapada,Shanghai)并参照相关文献中的方法[8],测定叶片中叶绿素和类胡萝卜素含量。
1.3.2 光合作用与叶绿素荧光参数
每处理固定2个植株,不作其他采样处理,对其相同部位的全功能叶片,使用便携式光合测定系统(LI 6400XT,USA),CO2浓度为400 μmol·mol-1,光照强度设置为1 000 μmol·m-2·s-1,参考Wang等[9]方法测定光合作用参数,测定在上午8:30—11:30进行,每处理重复测6次。同时,叶绿素荧光参数采用PAM-2500便携式调制叶绿素荧光仪(Walz,Germany),测定前用夹子夹住叶片,暗适应处理30 min,参照高丽丽等[10]方法测定有关参数,每处理重复测定6次。
1.3.3 叶绿体超微结构
取不同处理组植株叶片的相同部位(避开叶脉位置),用去离子水冲洗干净后,将叶片切成2 mm见方的小块,置于磷酸缓冲液(0.1 mmol·L-1,pH 7.0)配制的2.5%戊二醛离心管中,抽真空使叶片沉于戊二醛底部,放在4 ℃下固定过夜。小心吸出戊二醛,用1.0 mL磷酸缓冲液(0.1 mmol·L-1,pH 7.0)对样品进行漂洗,每次15 min,重复3次。弃去漂洗液,在通风橱环境中加入1.0 mL由磷酸缓冲液(0.1 mmol·L-1,pH 7.0)配制的1%锇酸,对样品固定2 h;弃去锇酸溶液,加入1 mL磷酸缓冲液(0.1 mmol·L-1,pH 7.0)对样品漂洗,每次30 min,重复4次。再参照Guo等[11]的方法,用透射电镜(Hitachi H-7650,Japan)观察叶绿体的超微结构。
1.4 数据处理
所有试验数据均汇总至Excel 2016,并利用DPS v 14.10软件[12]对相关数据进行统计分析,采用邓肯新复极差检验法进行多重比较,在α=0.05水平进行显著性检验,使用OriginPro软件2021版绘图。
2 结果与分析
2.1 叶绿素含量
如图1所示,与CK相比,0.3%NaCl处理的总叶绿素含量呈先下降后上升再下降的趋势,在处理30 d时总叶绿素含量开始高于CK,且在45 d时达到最大值,随后又开始逐渐降低并显著的低于CK;而对于0.6%NaCl处理,总叶绿素含量趋势上先上升而后下降,在处理1 d后总含量迅速上升并超过CK,约为CK的1.2倍,随后开始随着处理时间的延长而逐渐慢慢降低,并显著低于CK。对于0.9%NaCl处理,在处理1 d后总叶绿素含量便显著低于CK,并随着处理时间的延长逐渐降低,并始终低于CK。
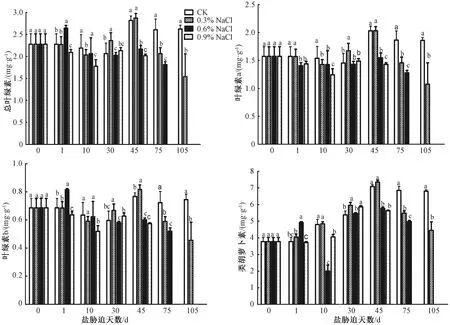
同天数比较柱上无相同小写字母者表示组间差异显著(P<0.05)。图2~3同。
此外,不同处理组中叶绿素a、b含量的变化与总叶绿素含量的变化趋势是类似的,同时,与CK相比,0.3%和0.6%NaCl处理的叶绿素a/b比值在处理前30 d差异不显著,而之后显著低于CK,而0.9%NaCl处理在10 d差异不显著,之后显著低于CK(表1)。NaCl处理下的胡萝卜素含量随处理时间的增加而逐渐的低于CK。由此可见,短期内低浓度水平(0.3%NaCl)处理可能使植株叶绿素的含量有所增加,但长期处理下3种浓度水平的NaCl处理均会导致叶绿素含量的降低。
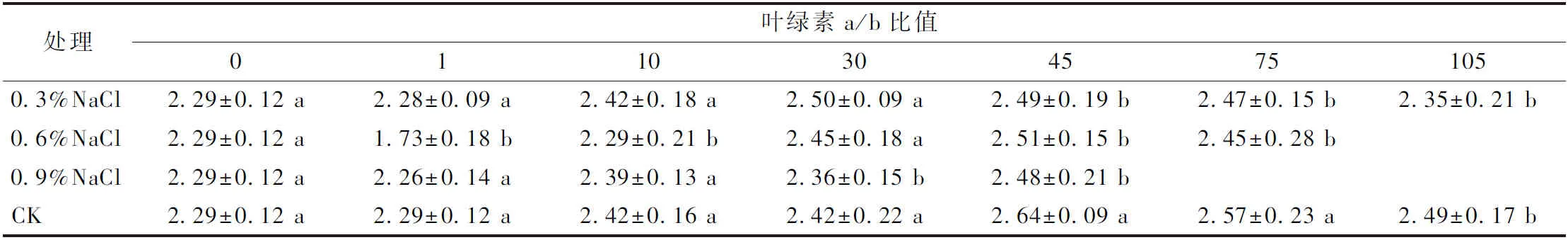
表1 不同浓度NaCl处理下叶片中叶绿素a/b比值
2.2 光合参数
如图2所示,与CK相比,0.3%NaCl处理的净光合速率(Pn)呈先下降后上升再下降的趋势,在处理前30 d,Pn值持续下降,而后Pn值开始回升,至处理45 d时达到峰值,并达到近似CK的水平,且一直持续到75 d,其Pn值一直与CK无显著差异。对于0.9%NaCl处理,其Pn值随着处理时间的延长呈下降趋势,但前期降幅较为缓和,在处理30 d时降为CK的83%,之后降幅迅速增加,到45 d时仅有1.49 μmol·m-2·s-1。由此表明,薄壳山核桃幼苗在0.3%NaCl短期处理下,光合作用会呈现一定的适应性,但3种浓度的NaCl长期处理均会使得Pn值降低,且降幅随着处理浓度和时间的延长而增加。此外,0.6%NaCl处理的细胞间隙CO2浓度(Ci)在处理1 d 后,增长为CK的1.61倍,并在后期处理中一直显著高于CK。而0.9%NaCl处理在1 d后增长为CK的1.35倍,并随着处理时间的延长而一直增长,在处理45 d时增长为CK的1.77倍,这意味着高水平盐浓度(0.6%和0.9%)处理下会使薄壳山核桃叶片的Ci值显著增加。同此,0.9%NaCl处理的蒸腾速率(Tr)值在处理1 d后便增长为CK的1.11倍,在处理10 d后达到峰值,为CK的1.43倍,直到盐处理后期(植株发生萎蔫时)才低于CK。这表明盐胁迫会显著影响薄壳山核桃的蒸腾速率,尤其是高盐浓度显著加剧了蒸腾速率。值得注意的是,0.9%NaCl处理的气孔导度(Gs)在处理1 d后就上升为CK的1.24倍,并一直保持高于CK,而该组中各数据的标准差较大,意味着高盐浓度下可能会由于离子平衡的紊乱使得薄壳山核桃气孔导度增加,且气孔开张的波动性较大。
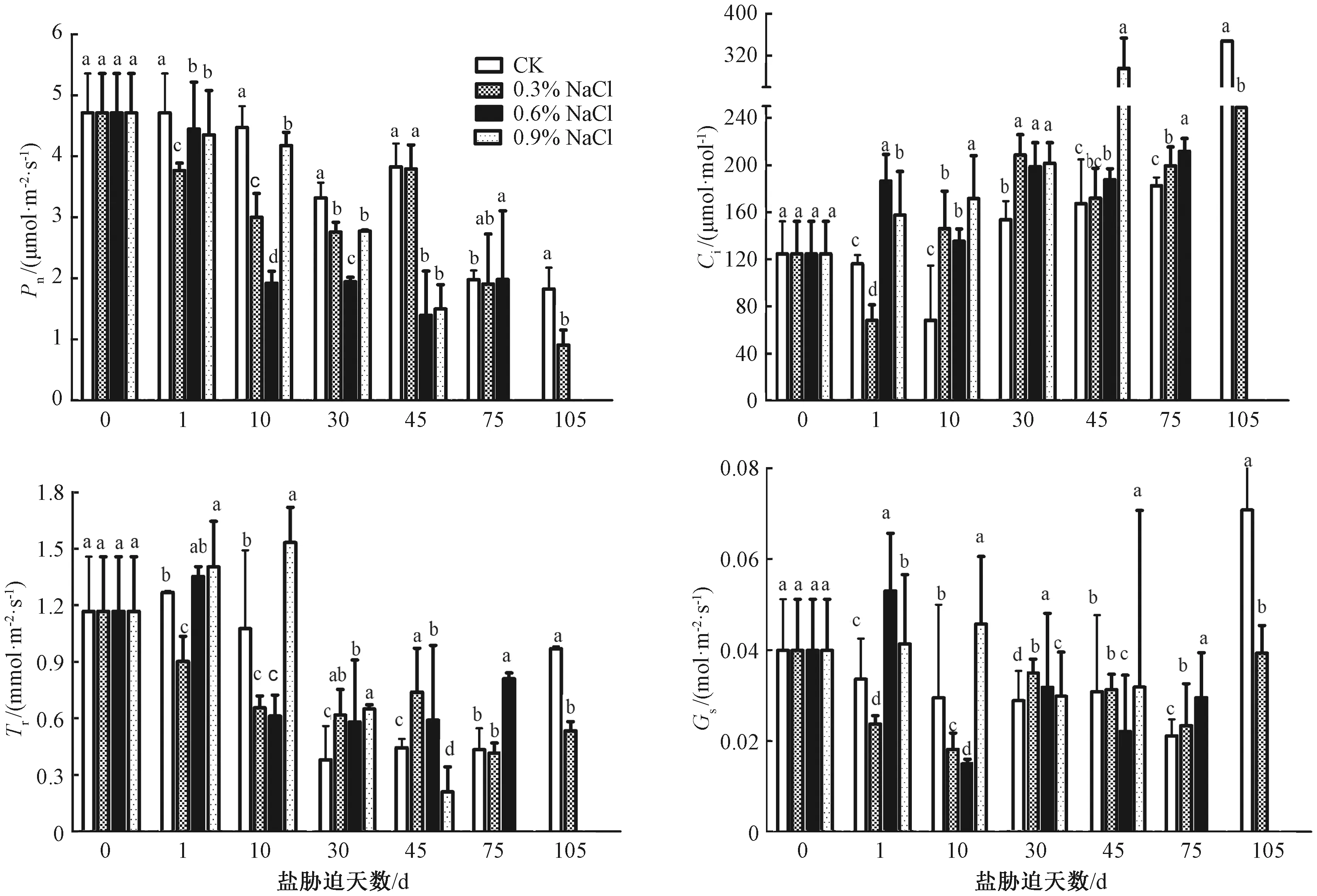
图2 不同浓度NaCl处理对叶片光合作用的影响
2.3 叶绿素荧光参数
由图3可知,0.3%NaCl处理在前45 d,其PSⅡ最大光化学量子产量(Fv/Fm)的值与CK无明显差异,在处理75 d时略微超过CK,但在处理105 d时降为CK的94%,这意味着该浓度水平盐胁迫对薄壳山核桃PSⅡ的活性影响较小。而高盐浓度水平(0.6%、0.9%)处理会迅速减弱PSⅡ的活性,但在处理后期,Fv/Fm又可恢复到正常水平,暗示着PSⅡ在该浓度下(0.6%、0.9%)受到的损伤是可逆的。类似的,0.6%和0.9%NaCl处理的非光化学淬灭系数(NPQ值)均呈先下降后上升的趋势,在处理前10 d均低于CK,而处理30 d后NPQ值迅速上升,并显著高于CK,并一直持续到盐处理后期,这意味着植株以热耗散形式散失的能量增多,减轻了PSⅡ光抑制程度,同时减缓了光合系统的损伤,可能是薄壳山核桃应对盐胁迫的一种保护机制。此外,在0.3%和0.6%NaCl处理的前期,通过降低光化学猝灭系数(qP值),减小PSⅡ的开放程度,降低光合速率,而处理后期通过qP值的升高增加PSⅡ的开放程度,进而减缓了光合速率的下降。
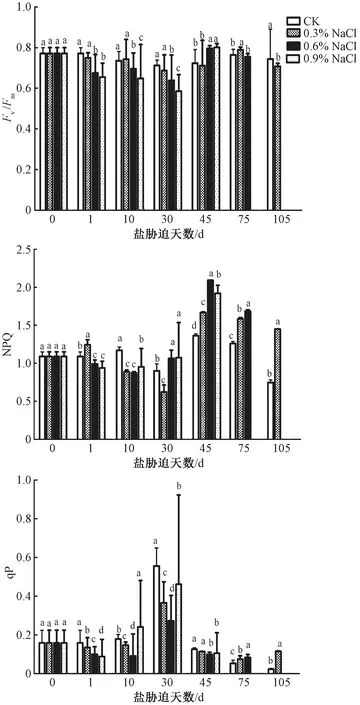
图3 不同浓度NaCl处理对叶片叶绿素荧光参数的影响
2.4 叶绿体超微结构
利用透射电镜对不同浓度NaCl处理10 d的叶片进行切片观察。图4显示,CK的叶绿体呈长椭圆形或橄榄球形,球体饱满,类囊体片层整齐,层数多,排列紧密,淀粉粒饱满积累在叶绿体中,叶绿体被膜清晰;0.3%NaCl处理下类囊体片层排列虽整齐紧密,但叶绿体膜已开始出现与细胞壁分离,叶绿体膜与细胞壁之间出现空隙;0.6%NaCl处理下叶绿体萎缩与细胞壁分离,类囊体片层排列不整齐、松散;0.9%NaCl处理下叶绿体扭曲呈扁圆形或球形,类囊体片层排列不整齐、稀疏。由此可见,不同浓度的NaCl处理均对叶绿体的结构造成一定程度的损坏,且受损程度随着NaCl浓度水平的增加而变得严重。
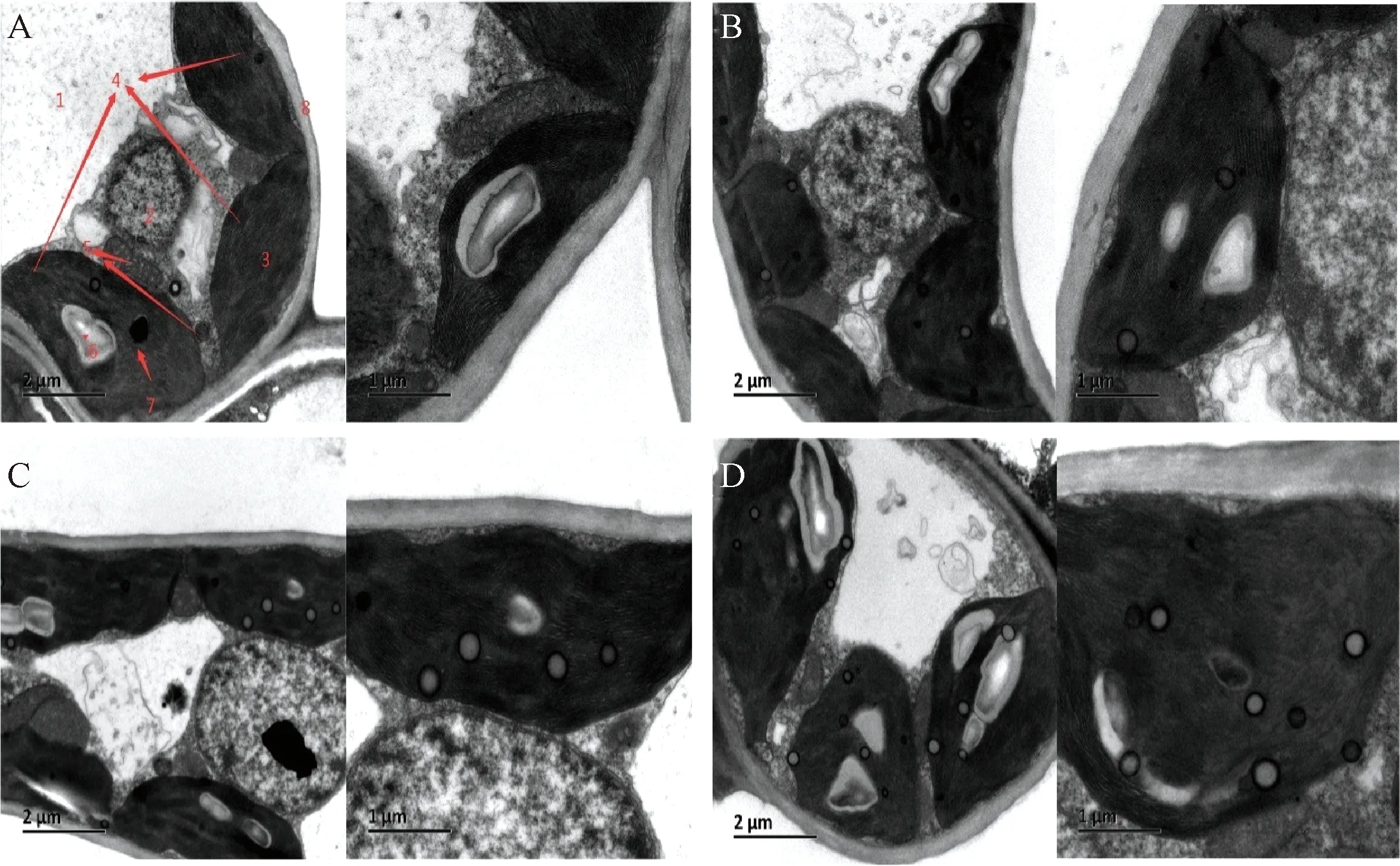
A、B、C、D分别为CK、0.3%NaCl、0.6% NaCl、0.9%NaCl处理,各处理中的左右图比例尺分别为2μm和1μm。1~8分别为中央大叶泡、细胞核、叶绿体、类囊体片层、线粒体、淀粉粒、噬锇颗粒和细胞壁。
3 小结与讨论
盐胁迫会对植物的光合作用造成一定的影响[13-14]。本研究发现,在0.6%和0.9%浓度处理下,薄壳山核桃Pawnee幼苗的Pn值整体上随着处理时间的延长而逐渐降低;而在0.3%NaCl短期时间处理下Pn值呈现一定的上升趋势并恢复到CK水平,光合作用会表现出一定的适应性,但在处理105 d时又低于CK,说明三种浓度的NaCl长期处理均会使得Pn值降低,且降低幅度随着处理浓度与处理时间的延长而增加。此外,NaCl胁迫条件下,光合作用的下降主要由气孔因素与非气孔因素两个原因所引起[15]。气孔因素主要是通过Gs的降低或关闭而引起CO2的供应不足,使光合速率下降,如低盐浓度下拟南芥、小麦光合速率的下降可归结于此类原因[16-17]。而非气孔因素则是通过改变光系统及碳同化过程关键酶活性等影响光合作用,如一定盐浓度下黄瓜、野生大豆光合作用的降低属于此类原因[18-19]。盐胁迫对光合作用的限制因素会随着NaCl浓度的不同而改变。在0.3%、0.6%NaCl处理下,酸柚苗Pn下降的主要原因是气孔限制,在0.9%NaCl处理下的Pn下降的主要原因是非气孔因素[20]。与酸柚苗相似的是,薄壳山核桃Pawnee在0.3%NaCl处理下,其Pn、Ci和Gs的变化趋势基本相同,说明在该浓度下,气孔因素可能是限制其光合作用的主要因素。对于0.6%NaCl处理组,其Gs随处理时间而变化的规律性不强,波动性较大;而0.9%NaCl处理组的Ci和Gs整体上随着处理时间的增加而增大,但Pn值却呈下降趋势,表明在高盐浓度(0.6%和0.9%)下,可能通过非气孔因素抑制其光合作用。
另外,叶绿素是植物进行光合作用的物质基础,在光能吸收过程中发挥着重要作用。同时,叶片叶绿素含量也是反映植物生理状态的重要指标之一[21-22]。盐胁迫条件下会使叶绿素发生降解,并降低植物光系统的活性[23-24]。本研究结果发现,短期内低浓度水平NaCl处理会使植株的叶绿素含量有所增加,这可能是对NaCl胁迫的一种适应性的表现,但长期处理下,3种浓度的NaCl处理依然会导致叶绿素含量发生不同程度的降低。因此,NaCl胁迫处理对薄壳山核桃PSⅡ的活性也造成一定影响。叶绿素荧光qP值的变化表明,在0.3%和0.6%NaCl处理的前期,可能通过减小PSⅡ的开放程度,降低光合速率;在盐胁迫处理后期,通过增加PSⅡ开放程度,可减缓光合速率的降低;0.9%NaCl则主要通过抑制qP值而减小PSⅡ的开放程度,降低光合速率。值得注意的是,在3种浓度NaCl处理的后期,Fv/Fm值又可以恢复到正常水平,表明PSⅡ的最大光化学效率是可以恢复的,PSⅡ并未受到非可逆性的损伤。此外,前期研究结果表明,盐胁迫会破坏叶绿体的结构,损坏光系统活性中心,使电子运输速率和碳同化速率显著降低,最终使植物的光合作用受到抑制[25-27]。本研究中透射电镜结果表明,3种浓度水平的NaCl处理均损坏了叶绿体的结构,且受损程度随着NaCl浓度的增加而更加严重,从而进一步解释了盐胁迫造成薄壳山核桃叶片光合作用降低的原因。
