桃红岭杜衡性状变异性及其对珍稀中华虎凤蝶保护的启示
2022-11-04杨文静吴问国刘小洪曾菊平
陈 亮,王 渌,杨文静,吴问国,刘小洪,张 毅,曾菊平*
(1.江西农业大学 林学院/鄱阳湖流域森林生态系统保护与修复国家林业和草原局重点实验室,江西 南昌 330045;2.江西桃红岭梅花鹿国家级自然保护区管理局,江西 九江 332700;3.江西庐山森林生态系统定位观测研究站,江西 九江 332900;4.江西庐山国家级自然保护区管理局,江西 九江 332900)
【研究意义】植物性状(Plant trait)是植物对外界环境长期响应与适应后所呈现的可度量特征[1-2],如来自形态、生理、生化等多方面特征。监测研究这些特征的变异性、稳定性,成为当前洞悉植物个体、环境和生态系统结构、过程及功能多层面联系与变化的重要途径,故也称植物功能性状(Plant functional trait)研究[2]。其中,诸如叶片形态特征(如叶片大小)、根属性、种子大小等体现植物对环境适应并(或)影响植物生产力等性状、指标最易被研究者选择[3-4]。植物形态性状变异反应了植物对环境的适应性变化[5],即植物个体或种群因生长在不同环境,而在形态、性状上出现差异或变异性[6-7],这种变异性可反映植物与环境的协同进化关系,以及其适应机制[8-10]。通常,植物营养器官(尤其叶片等地上部分)功能性状变异更普遍[11],繁殖器官性状则变异小,相对更稳定。事实上,叶片等营养器官的形态(如叶面长毛、叶片增厚等)、生理、生化(如次生物质产生或增加)性状变异能直接影响植食性昆虫适口性,影响其选择习性或取食行为,形成针对植食性昆虫的抗性机制。受此影响,昆虫不得不调整生存策略,种群发生、空间分布也因此发生变化。喙凤蝶属Teinopalpusspp.、虎凤蝶属Luehdorfiaspp.等珍稀蝶种的寄主植物选择性明显,为狭食性或专食性种类[12-20]。它们野外少见,发生点零散,分布区狭窄,易受寄主资源限制,包括数量(如株数、密度)与质量限制,如由植物性状变异产生的质量变化(如适口性下降)。尤其,这些珍稀蝴蝶野外种群小,当前如何促进其种群扩增、恢复是保护管理面临的关键问题,解决该问题不仅需围绕蝴蝶自身(如种群内禀增长率)展开研究,也需对其专性寄主植物开展调查,以弄清资源限制作用,包括与寄主植物性状变异有关的质量限制作用。当前珍稀蝶类保护研究很少考虑野外寄主植物性状变异的影响,因而可参考的案例少。【前人研究进展】植物功能性状及其变异性可一定程度反馈环境变化(如气候变暖)重要信号,反应植物应对环境变化的适应能力与机制[21],例如,植物叶片功能性状及其变异当前备受植物学、生态学研究关注[2,22],因为叶片作为光合作用重要场所,功能性状变异能表达环境胁迫下植物的生态适应与进化,这主要基于其在地理、气候、生境等梯度上留下的适应痕迹[23-24],性状变异常在种间与种内两个层面发生,而前者更常见[25],成为自然界物种共存的基础,所以颇受研究关注;而有关种内变异则仍关注不够[25-27],其生态功能仍不明确。其实,种内性状变异也是植物应对环境变化的重要响应方面,也是预测植物动态变化的重要内容[28-30]。研究种内性状变异有助于理解物种更替、种内基因型差异及环境条件导致的形态可塑性[28-30]。事实上,多数物种主要通过功能性状的可塑性与遗传多样性来适应环境或生境变化,因而洞晓种内性状变异能提高对群落物种多样性及其维持机制的理解,并预知不同环境的群落组成[26,32-33]。
【本研究切入点】基于珍稀中华虎凤蝶桃红岭种群分布狭窄、种群孤立的现状[34],本研究选择当地专性寄主植物杜衡Asarum forbesii为研究对象,探究寄主植物功能性状的种内变异(个体、种群水平),以及其对当地中华虎凤蝶种群发生、分布的潜在影响。杜衡为马兜铃科Aristolochiaceae 多年生草本[35],常生长在林下沟边阴湿地,而桃红岭杜衡包括在低海拔林下沟边的发生种群,与也有在高海拔山顶灌草地的发生种群,两地环境差异大,杜衡种内潜在变异性高。尤其,比较两个不同杜衡种群的性状与变异性,或有助于提高对桃红岭中华虎凤蝶当前发生、分布格局的理解。当前,桃红岭中华虎凤蝶仅在山顶灌草地发生,而在杜衡低海拔种群无访问记录,这是否与两个杜衡种群的功能性状变异有关,尤其是叶片功能性状。例如,有学者对朝鲜半岛中部的乌苏里虎凤蝶野外调查,发现雌蝶可能通过叶片大小识别来决定每片叶子的产卵量[36]。为此,本研究拟分别从中华虎凤蝶高海拔占据区与低海拔非占据区采样,通过测量各区杜衡植株根茎叶形态指标,分析两个区杜衡形态性状变异性,探究当地杜衡的适应性特征,并揭示可能参与影响中华虎凤蝶发生的关键性状。【拟解决的关键问题】杜衡是桃红岭中华虎凤蝶唯一寄主植物,其资源的数量或质量特征可能都参与影响该珍稀蝶种的发生、分布。本次在不同区域进行单株取样,一方面拟通过测量营养器官根茎叶与繁殖器官花、种子形态性状、指标(如叶长、叶宽等),揭示杜衡种内性状变异特征,分析性状稳定性或变异性,以及各部位功能性状间的关联性;另一方面,通过比较中华虎凤蝶占据区与非占据区杜衡种群间性状,探讨寄主植物功能性状变异及其与中华虎凤蝶发生、分布的耦合关系、保护意义,为后续基于寄主资源途径的珍稀蝶类生境保护,以及可能的人工建群、野外放归试验提供科学依据。
1 材料与方法
1.1 研究区域
选择赣北桃红岭梅花鹿国家级自然保护区为研究区域,位于江西省九江市彭泽县境内(29°42′~29°53′N,116°32′~116°43′E),见图1。当地中华虎凤蝶为江西省当前唯一残存种群,其幼虫已知专性取食当地多年生草本植物杜衡[34]。前期调查发现,当地杜衡的发生点明显多于中华虎凤蝶,从低海拔分布到高海拔,涉及林下、沟边、竹林、灌丛等多类生境。而当地中华虎凤蝶目前仅在山顶灌丛区有发生,且只在猫鹰窝、桃红山两个点有记录。为掌握当地杜衡种群的形态、性状变异与适应性、探究与性状变异相关的寄主植物质量对中华虎凤蝶发生的潜在影响,杜衡植株调查、取样覆盖全区进行。同时,基于中华虎凤蝶发生状况,将杜衡发生区划分成蝴蝶占据区(如高海拔猫鹰窝)与非占据区(如低海拔竹林),分别进行取样。
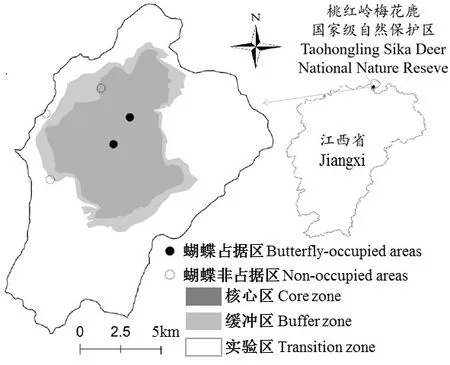
图1 研究区桃红岭国家级自然保护区位置与杜衡调查取样区(中华虎凤蝶占据区与非占据区)Fig.1 Study area location of Taohongling National Nature Reserve and Asarum forbesii sampling areas occupied or non-occupied by the butterfly of Luehdorfia chinensis
桃红岭保护区总面积12 500 hm2,地貌为平缓起伏的低山丘陵,海拔多在100~500 m,其中最高峰为猫鹰窝(海拔536.6 m)。区内气候属亚热带季风气候类型,全年季节性变化明显,日照充足,雨量充沛,主要植被包括草丛、灌丛、针叶林、阔叶林、竹林等[37]。
1.2 野外调查与形态测量
前期调查获知研究区杜衡多在3 月上旬进入抽叶期,为对完整植株取样,调查选在4—6 月份进行。采用样线法,沿非硬化道路、小路、林间通道、山脊等行进,用GPS仪记录线上杜衡植株的单株位置信息、编号,计数单株叶片数,并用米尺(精确度1.0 mm)测量地上部分(叶)形态指标,包括株高、每片叶叶长、叶宽(取最宽处测量值)、叶柄基部直径(基径)、叶柄顶端直径(端径)等。之后,选择发育完整植株(带有花器官),适量带土,单株采集放入透明自封袋,用防水记号笔写下编号,带回室内计数,测量其他地上或地下部分形态指标。每个点采集5株或以上个体。
将采回的杜衡植株用清水轻轻冲洗,去除泥土后,用网格法测量叶面积[13,38]。同时,计数侧根数量,用米尺或游标卡尺测量地下根、茎的形态指标,包括茎总长、第1至6茎节间长、根直径等。另外,为分析繁殖器官的形态变异性,选择花型饱满植株,测量花部形态性状,包括花柄长、花柄直径、花被长、花被直径、花被裂片长、花被裂片直径(取最宽处测量值)、喉孔直径。并用解剖刀将花部实体纵向切开,测量雄蕊长、雌蕊长;同时,取出内部饱满的种子,测量种子长、种子直径。
1.3 形态性状指标与计算
对杜衡植株地上部分(叶)、地下部分(根茎)与繁殖器官(花部、种子)的形态、性状指标进行测量、计算,性状、指标定义、描述与缩写符号等见表1。

表1 杜衡地上、地下的营养器官与繁殖器官形态性状、指标的定义、描述Tab.1 Definitions and descriptions of the up-ground,underground nutritive and propagative organs’morphology traits and indicators in Asarum forbesii
基于部分测量植株(n=40),以叶面积为因变量,叶长、叶宽、长宽比、长宽乘积为自变量进行多元线性回归分析,采用逐步回归法,获得最优模型y=0.007x+7.056(R2=0.771,P<0.001),其中,y为叶面积,x为长宽乘积。结果与其他植物一样[23,39-40],杜衡叶片长宽乘积的变化与叶面积变化较为一致。而考虑用网格法无法测量野外植株叶面积,本次选择对长宽乘积进行变异性分析,代表叶面积的变异性。
将性状测量数据输入Excel,用“STDEV()”与“AVERAGE()”函数计算平均值Mean、标准差SD,再用CV=SD/Mean 公式计算变异系数(Coefficient Variation,CV),据CV 大小确定各性状变异程度,包括:(1)CV<0.2,弱变异;(2)0.2≤CV<0.5,中等变异;(3)CV≥0.5,高度变异[41]。
1.4 数据分析
用Wilcoxon 秩检验比较蝴蝶占据区与非占据区杜衡地上部分性状差异。用Wilcoxon(配对)秩检验比较不同区域性状变异系数的总体差异。在R4.0.3 中用corrplot 数据包的“cor()”函数计算Spearman 秩相关系数,分析各性状的关联性,并基于显著性分析与相关系数r判定性状间的关联,包括:(1)r<0.3 为无关联;(2)0.3≤r<0.8 为弱关联;(3)r≥0.8 为强关联。显著水平设为P<0.05,全部分析在SPSS Statistics 22.0或R4.0.3[42]完成。
2 结果与分析
2.1 杜衡形态性状变异性
2.1.1 地上部分变异性从表2 可知,比较来自中华虎凤蝶占据区、非占据区或总体区域的杜衡地上叶部各性状指标间均无显著差异,变异系数CV也差异不显著。在所选性状、指标中,叶部长宽乘积变异系数最大,各区均为高度变异(CV>0.5)。而蝴蝶占据区叶长也为高度变异,且较其他区(中等变异)程度更高。相对地,叶部的叶柄基径变异程度最低。

表2 桃红岭杜衡的中华虎凤蝶占据与非占据区叶部性状比较Tab.2 Leaf traits comparison of A.forbesii between areas occupied or non-occupied by the butterfly of L.chinensis in Taohongling
2.1.2 地下部分变异性表3 显示地下部分根状茎与根部变异较高,总体CV 值逼近0.5,接近高度变异水平。根状茎各节间长的变异性差异大,CV 值从0.17(弱变异)至0.96(高度变异)不等(表3),无确定性。

表3 桃红岭杜衡的地下根状茎、根部性状变异系数Tab.3 Coefficient variation of underground rhizome and root traits of A.forbesii in Taohongling
2.1.3 繁殖器官变异性表4显示杜衡繁殖器官变异小,总体CV 为0.18,为弱变异。事实上,所选11个性状中仅花柄长、花被裂片长、雄蕊长为中等变异(CV>0.2),而雄蕊长变异程度相对更高(CV=0.27)。

表4 桃红岭杜衡的繁殖器官性状变异系数Tab.4 Coefficient variation of the reproductive organ traits of A.forbesii in Taohongling
2.1.4 各部分变异性比较杜衡地上与地下部分营养器官性状变异系数总体差异不显著(图2),但营养器官整体或地上、地下部分的变异性均显著高于繁殖器官(图2)。
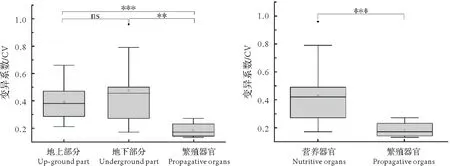
图2 桃红岭杜衡营养器官(含地上与地下部分)与繁殖器官性状变异性比较Fig.2 Comparison of trait coefficient variation between nutritive(up-ground and underground parts)and propagative organs of A.forbesii in Taohongling
2.2 杜衡形态性状的关联性
图3显示杜衡性状关联性在中华虎凤蝶占据区与非占据区并不一致,如在非占据区,株高与4个性状显著关联,但在占据区未发现显著关联;在占据区,叶柄基径与叶长关联性显著,与叶宽无显著关联,而非占据区情况相反,叶柄基径与叶宽显著关联,却与叶长无显著关联(图3)。另外,非占据区的单株叶数与叶柄基径显著负关联,占据区则无显著关联。然而,叶长宽乘积在两个区均与基径、端径显著关联(图3)。

图3 桃红岭杜衡中华虎凤蝶占据与非占据区地上部分性状相关矩阵图Fig.3 Correlation matrix among traits of upground parts of A.forbesii in butterfly occupied and non-occupied areas in Taohongling
而从图4 可知,桃红岭杜衡营养器官地上部分株高与地下部分性状均明显关联,但其他性状几乎与地下部分、繁殖器官性状无显著关联。相反地,地下的根状茎、根部性状则与繁殖器官花部性状均显著相关,且关联度逼近或达到强关联水平(r≥0.8,图4)。而与其他器官部位相比,花部性状内部间均具强关联性。然而,繁殖器官种子性状与其他各部性状关联度低,仅种子直径与花部花柄长、花柄直径等呈弱关联性(图4)。

图4 桃红岭杜衡地上、地下营养器官与繁殖器官形态性状相关矩阵图Fig.4 Correlation matrix among up-ground,under-ground nutritive and propagative organs traits of A.forbesii in Taohongling
3 结论与讨论
调查野生植物功能性状,分析其变异性或稳定性,能提高人们对植物个体、环境及其所在生态系统结构、功能等多层面关系的理解,提高对物种共存与物种分布格局的认识[26]。本研究对桃红岭野生杜衡的根、茎、叶、花、种子形态性状进行测量、比较,发现种群内部营养器官性状的变异性高,接近高变异水平,而繁殖器官性状变异弱,显著低于营养器官(包括地上、地下部分)的变异水平,这种情况较为常见,如黄花苜蓿[43]、曼陀罗[44]、华中五味子[45]等植物。事实上,繁殖器官作为植物进行种群扩散和后代繁殖的载体,其性状的稳定性是植物物种延续与后代持续的重要保证,因而它们总体倾向保守,性状变异程度低[46-47]。不仅如此,繁殖器官的性状稳定性或保守性,也体现在其内部性状的强关联性上。例如,杜衡花部大部分性状间关联性均达到强关联水平。然而,杜衡营养器官内部性状间的关联性则整体偏弱,且不同发生区关联性不同。例如,在中华虎凤蝶非占据区,杜衡株高与4个性状显著关联,而在占据区则未发现类似关联性。
植物生长、存活依赖营养器官根、茎、叶的相关功能(如支撑功能)与作用(如光合作用),有赖于根、茎、叶对环境的适应力,并常体现在其性状的可塑性与变异性上。本研究用变异系数(CV)大小表示各性状的可塑性强弱或变异性高低[48-49],发现高度变异性状既出现在杜衡的地上部分营养器官,如叶部的长宽乘积(相当于叶面积),也出现在地下部分营养器官,如根状茎长、根部的侧根数等。而类似地,马蹄参[50]、独花兰[51]等植物的营养器官性状也具有高变异性。叶片功能性状直接影响植物获取资源(如光照)、利用资源(如光合作用)的能力,尤其叶片大小、面积等性状,可集中反映植物在长期适应环境变化过程形成的生存策略[24]。例如,阔叶树种被认为是对温湿环境长期适应进化的结果,但常绿阔叶树种叶面积常显著小于落叶树种[26],反映其生存策略不同。
叶面积性状变异不仅在种间发生,也可能种内发生,如在不同的种群间。通常,位于阴蔽、湿润、资源丰富区域的植物种群可能具有更大的叶片[52-54],以适应植物在不同环境下气体、能量交换与调节平衡的需要。本研究比较杜衡低海拔林区与高海拔灌草丛种群,也发现前者叶片长宽乘积或叶面积较后者更大些,但两者差异仍未达显著水平(P>0.05),这可能与受两个种群内部叶面积性状的高CV 值影响,即两个种群叶面积性状均达高变异水平,说明当地杜衡叶片在叶面积上表现出宽幅适应性。这种情况似乎与当地中华虎凤蝶的发生分布格局不一致,因为蝴蝶发生仅占据了寄主植物高海拔种群。而这种不一致性似乎也暗示当地中华虎凤蝶在发生、分布上可能与寄主植物叶片性状无关。因为,两个杜衡种群的其他叶部性状间也都未检测到显著差异,包括变异系数CV值。因此,叶部性状的高变异性,反映的是杜衡在当地环境变化(如海拔梯度)下的生存策略,即宽幅适应策略或宽生态位,而专食性昆虫中华虎凤蝶的取食压力可能在其生存策略形成过程中影响甚微。也就是说,若忽略其他气候、地形等条件,中华虎凤蝶也可能在低海拔寄主植物种群区发生,预示着人工寄主植物资源干预途径(如人工增加野外杜衡资源)的可行性,以帮助蝴蝶种群在更短时间内恢复、扩大,从而提高野外种群应对气候变暖、干旱化趋势的减缓能力[20,55],降低小种群效应与局域灭绝风险[20,56-57]。
受物理、生理等因素综合影响,植物功能性状间常呈现出一定的协变和权衡关系[58-59]。本研究分析杜衡不同部位的性状关联性,发现地上部分株高与地下部分根状茎、根的几个性状明显关联,而与繁殖器官性状无显著关联,但地下部分与繁殖器官则具有强关联(r≥0.8)。这表明杜衡地下部分无论对地上部分还是繁殖器官,都能产生影响,即后两者生长发育对地下部分存在依赖性,部位间的性状存在协变关系。例如,地上的株高性状可能受地下根状茎长、根直径与侧根数性状影响,存在正向协变关系。同样地,杜衡花部性状(如花柄长、花被长等)也与地下部分的一些性状存在正向协变关系。这种情况主要与杜衡的多年生草本习性有关。在桃红岭,调查获知杜衡10月份左右落叶后,完全依靠地下部分生活,应对不良环境(如越冬),而来年3月份的抽叶质量,及后续的开花结籽情况等,可能都与地下部分前一年的资源储备有关。但是,值得提及的是,地上-地下性状协变关系仅在少数性状间出现,且不同区域种群间情况并不相同。例如,中华虎凤蝶占据区的杜衡种群,其株高与地下部分性状的关联并不显著,这可能与高海拔区杜衡地上部分所受的更大环境压力(如低温、大风等)有关,也就是说,在环境胁迫下,性状间的协变关系也会发生变化,以满足植物在当地所需的生存策略调整要求。因而,在中华虎凤蝶保护管理中,通过同时为地上部分与地下部分提供良好的资源条件(如在杜衡人工培育条件下),发挥出植物各部位性状间的协变机制,或能显著提高杜衡植株生长质量与繁育能力,进而研究如何以人为干预方式,提高桃红岭的野外寄主资源供给数量与质量,促使中华虎凤蝶种群恢复、扩大(如人工干预下的小种群建立等),达到降低地方或局域灭绝风险的保护目标[20,56-57]。
致谢:江西桃红岭梅花鹿国家级自然保护区管理局与鄱阳湖流域森林生态系统保护与修复国家林业和草原重点实验室开放基金课题(PYHKF-2020-03)对本次野外调查提供了帮助与支持,谨致谢意!
