林窗改造对马尾松人工林土壤动物群落结构的影响
2022-11-02高雅荣罗熳丽鲁琴琴张庭昊肖玖金
高雅荣 ,蒋 罗 ,罗熳丽 ,5,鲁琴琴 ,张庭昊 ,肖玖金 **,张 健
(1.长江上游林业生态工程四川省重点实验室,成都 611130;2.长江上游森林资源保育与生态安全国家林业和草原局重点实验室,成都 611130;3.华西雨屏区人工林生态系统研究长期科研基地,成都 611130;4.彭州市规划和自然资源局,四川 彭州 611930;5.渠县自然资源局,四川 渠县 635200;6.成都市农林科学院,成都 611130)
马尾松人工林在川南地区广泛分布,良好的马尾松人工林生态系统是长江上游生态安全的重要保障。作为我国南方地区重要的用材树种和荒山绿化的先锋树种[1],在缓解木材供需矛盾的同时,也充分发挥着保持水土、净化空气的作用[2],然而长期过度的纯林化经营,使其林分结构逐渐趋于单一化致使病虫害频发[3],同时频繁的人类活动干扰,使马尾松人工林出现严重的水土流失[4],造成大面积马尾松人工林生态系统退化[5]。且我国人工林主要分布于人类生产活动较为活跃的区域,普遍存在生产与生态的重要矛盾。因此,对马尾松人工林进行近自然化改造亟待进行。
林窗作为森林生态系统中普遍存在的一种干扰形式和森林生态过程的重要环节[6-7],在人工林近自然化改造中得以广泛应用。在林窗中补植珍贵乡土树种兼顾了经济及生态效益,形成了乡土树种与人工林的斑块混交模式,这种模式能有效促进凋落物分解和养分循环,改善土壤肥力状况,提高人工林生态系统稳定性和维持较高的生产力,进而有利于生产与生态的协同发展。
土壤动物作为地下生态系统物质循环过程中的重要部分,对细微环境变化敏感[8-9],在分解生物残体、改变土壤理化性质以及促进土壤生态系统物质循环与能量转化过程中起着重要作用[10],可作为生态系统稳定性的生物学指标[11]。因此,可利用土壤动物的指示作用研究整个生态系统。目前对于林窗的研究主要集中在天然形成的林窗,相关学者对其林窗内的微生物多样性[12]、土壤理化性质[13]、植物多样性[8]、土壤碳库[14]以及凋落物分解[15]等方面,均认同了林窗对森林的生态恢复总体上有正向作用。然而,对通过人工间伐形成的林窗内的土壤动物群落特征仅有少量报道,这些报道发现,柳杉人工林中林窗的形成降低了土壤动物的密度,肖玖金等[16]的研究仅揭示了林窗土壤动物群落结构特征。赵波[17]对马尾松人工林的研究表明在西南地区的生产实践中,可采用中型(625~900 m2)林窗择伐。目前,林窗作为一种较强且有效的人工干扰措施[18],土壤动物群落对人工林窗大小的长期的响应还有待进一步研究。课题组对马尾松人工林进行林窗改造,并在林窗中补植1 a生珍贵乡土树种油樟,研究马尾松人工林开窗补植油樟8 a后油樟斑块的土壤动物群落特征。探讨不同面积林窗中中小型土壤动物群落特征,以期为马尾松人工林近自然改造效果评估提供依据,并加深对植被和土壤生物区系的相互作用的认知。
1 材料和方法
1.1 研究区概况
研究地位于长江上游四川省高县来复镇毛巅坳林区经营小班 (104°21′37′′~104°34′22′′E,28°35′45′′~28°36′19′′N),属低山丘陵地貌,海拔在400~450 m,属亚热带湿润季风气候,平均气温18.1℃,年均降水量1 021.8 mm,年日照时数1 148 h,土壤为黄壤土,土层厚度约50 cm。各样地林下植被主要灌木有柃木(Eurya japonica)、展毛野牡丹(Melastoma normale)、肖梵天花(Urena lobata)、铁仔(Myrsine africana)、枹栎(Quercus serrata)、梨叶悬钩子(Rubus pirifolius)、香花崖豆藤(Millettia dielsiana)和荚蒾(Viburnum dilatatum)等;主要草本为苦买菜(Ixeris polycephala)、戟叶堇菜(Corydalis bungea)、麦 冬(Ophiopogon japonicus)、芒(Miscanthus sinensis)、芒萁(Dicranopteris dichotoma)和爵床(Rostellularia procumbens)等。
1.2 样地设置
课题组于2011年对39 a生的马尾松人工林进行团状采伐,形成面积为小型林窗(100~225 m2)、中型林窗(625~900 m2)和大型林窗(1 225~1 600 m2),每个类型大小林窗9个,以未采伐的马尾松人工纯林作为对照(CK)。各林窗地形地貌、海拔、母岩、土壤类型、坡度、坡位等相同或相近,形状为正方形。为防止相互干扰,各林窗间间隔均大于10 m。2012年3月,在各林窗内按2.5 m×2.5 m的密度补植1a生珍贵乡土树种油樟。油樟生长状况良好,树高达7.5 m,冠幅达3 m×4 m,郁闭度在0.79~0.88之间。
1.3 研究方法
1.3.1 试验方法
于2018年9月在各样地内随机设置3个以“品”字形分布的样方,各样方面积为0.25 m2(50 cm×50 cm)。在各小样方土壤中,分0~5 cm层、5~10 cm层和10~15 cm层,用环刀(r=5 cm,v=100 cm3)自上往下依次取土样,用Tullgren干漏斗法和Baermann湿漏斗法分离土样中的土壤动物[19-21];同时在各样点收集10 cm×10 cm(0.01 m2)面积的枯落物进行分离。
土壤动物的分离在烘虫箱中进行,烘虫时控制铺土厚度约3 cm,土壤湿度约80%,并用温度控制器将土表温度控制在34~36℃[21-22],烘虫时间均为48 h。干生漏斗分离的土壤动物用盛有75%浓度的酒精培养皿收集,在解剖镜(SZ760)下观察计数;湿生土壤动物则用清水收集。干生漏斗分离每隔12 h观察一次,湿生每隔4 h观察一次,观察间隔时间逐步增长,对观察到的所有土壤动物进行分类并计数。
土壤动物的分类鉴定参照《幼虫分类学》[23]、《昆虫分类检索》[24]、《中国土壤动物检索图鉴》[25]和《中国亚热带土壤动物》[26]等资料,对所采集到的土壤动物进行分类鉴定,一般鉴定至目、科等较高的分类阶元。
1.3.2 数据分析与处理
土壤动物多样性分析采用Shannon-Wiener多样性指数、Margalef丰富度指数、Pielou均匀度指数和Simpson优势度指数对土壤动物群落多样性特征进行分析[27-29]。
多样性指数(Shannon-Wiener):H′=∑PilnPi
丰富度指数(Margalef index):D=(S-1)/lnN
均匀度指数(Pielou index):E=H′/lnD
优势度指数(Simpson index):C=∑(Ni/N)2
式中:ni为第i个类群的个体数;N为所有类群的个体数;S为类群数。
中小型土壤动物各类群等级划分:个体密度大于捕获总量密度10.0%及其以上的为优势类群(+++),占1.0%~10.0%的类群为常见类群(++),不足1.0%的类群为稀有类群(+)[30]。
土壤动物营养功能类群划分:参照张雪萍等人的研究[31],将收集的中小型土壤动物按照取食对象的差异分为植食性、杂食性、菌食性、腐食性、捕食性等5个营养功能类群。
数据的处理和分析采用Microsoft excel 2013和SPSS 25.0完成,采用Origin 8.1绘制图形。土壤动物群落特征差异采用单因素方差分析(One-way ANOVA)和最小显著差异法(LSD)检验,显著性水平设定为P=0.05。
2 结果分析
2.1 中小型土壤动物群落组成
本试验共采集中小型土壤动物2 518头,隶属3门12纲30目112科(表1),以线虫纲(Nematoda)为优势类群,占总个体数的50.37%;常见类群有等节䖴科(Isotomidae)、蚁科(Formicidae)、懒甲螨科(Nothridae)和跳虫科(Poduridae)等11种,共占个体数的27.59%;稀有类群有长须螨科(Stigmaeidae)、盾螨科(Scutacaridae)、奥甲螨科(Oppioid mites)、寄螨科(Parasitidae)和跗线螨科(Tarsonemidae)等100种,共占个体数的22.04%。各样地土壤动物平均密度排序:大型>中型>小型>纯林;土壤动物类群数依次为大型>中型>小型>纯林。

表1 不同林窗土壤动物群落组成统计Table 1 Statistics of soil animal community composition in different forest gaps

(续 表 1)

(续 表 1)
其中,马尾松纯林平均密度为1.03×104ind/m2,类群数为34类252只,小型林窗平均密度为3.18×104ind/m2,类群数为65类576只,中型林窗平均密度为5.30×104ind/m2,类群数为75类847只,大型林窗平均密度为 5.48×104ind/m2,类群数为 80类843只。
2.2 中小型土壤动物群落水平分布变化特征
土壤动物个体密度和类群数量随林窗面积的增大而发生变化(图1)。如图1a所示,土壤动物平均密度随林窗面积的增加而增加。大型和中型林窗均显著高于小型林窗和马尾松纯林(P<0.05)。土壤动物平均密度在大型林窗达到最高,达2.74×104ind/m2以上。纯林密度最低,为1.03×104ind/m2以上。如图1b所示,与纯林相比,各林窗内土壤动物的类群数均有所增加,林窗改造有利于土壤动物的类群数增加。中型和大型林窗样地土壤动物类群数均显著高于纯林(P<0.05),中型林窗土壤动物类群数最多。

图1 不同林窗大小中小型土壤动物群落平均密度及类群数(n=9,mean±SE)Figure 1 The meso and micro-soil faunal group number and density in different gap sizes
2.3 中小型土壤动物群落垂直分布特征
各处理下,土壤动物平均密度和类群数在土壤层中的分布随土层的加深而下降。如图2a,与纯林相比,除小型林窗的凋落物层以外,各层土壤动物平均密度均有所增加。凋落物层中大型林窗显著高于纯林和小型林窗;0~5 cm层中型和大型林窗显著高于纯林;10~15 cm层中型和大型林窗显著高于纯林,中型林窗显著高于小型林窗和纯林;10~15 cm层中大型林窗显著高于小型林窗和纯林。此外,中小型土壤动物主要生活在0~5 cm层土壤中,占总密度的48.81%,平均密度高于凋落物层、5~10 cm层和10~15 cm层3个层次,因此土层越深,平均密度越低。
如图2b,与纯林各层相比,其他林窗凋落物层、0~5 cm、5~10 cm层和10~15 cm层土壤动物类群数均有所增多,在凋落物层和0~5 cm层中各林窗均不显著;5~10 cm层中型林窗显著高于小型林窗和纯林;10~15 cm层中型和大型林窗显著高于纯林。

图2 不同林窗大小中小型土壤动物群落的垂直分布(n=9,mean±SE)Figure 2 The vertical distribution of meso and micro-soil faunal in different gap sizes
2.4 中小型土壤动物的群落多样性特征
由图3可以看出,土壤动物的群落多样性指数、均匀度指数、丰富度和优势度指数随林窗改造面积的改变。总体来看,林窗改造增加了土壤动物的优势度指数和丰富度指数,降低了多样性指数和均匀度指数。各林窗间土壤动物多样性指数差异不显著;中型和大型林窗均匀度指数显著低于纯林,大型林窗优势度指数显著高于纯林,中型林窗丰富度指数显著高于纯林。多样性指数和均匀度指数在纯林出现最大值,在丰富度指数出现最小值;中型林窗的丰富度最高;大型林窗的优势度最高,多样性指数和均匀度指数最低。
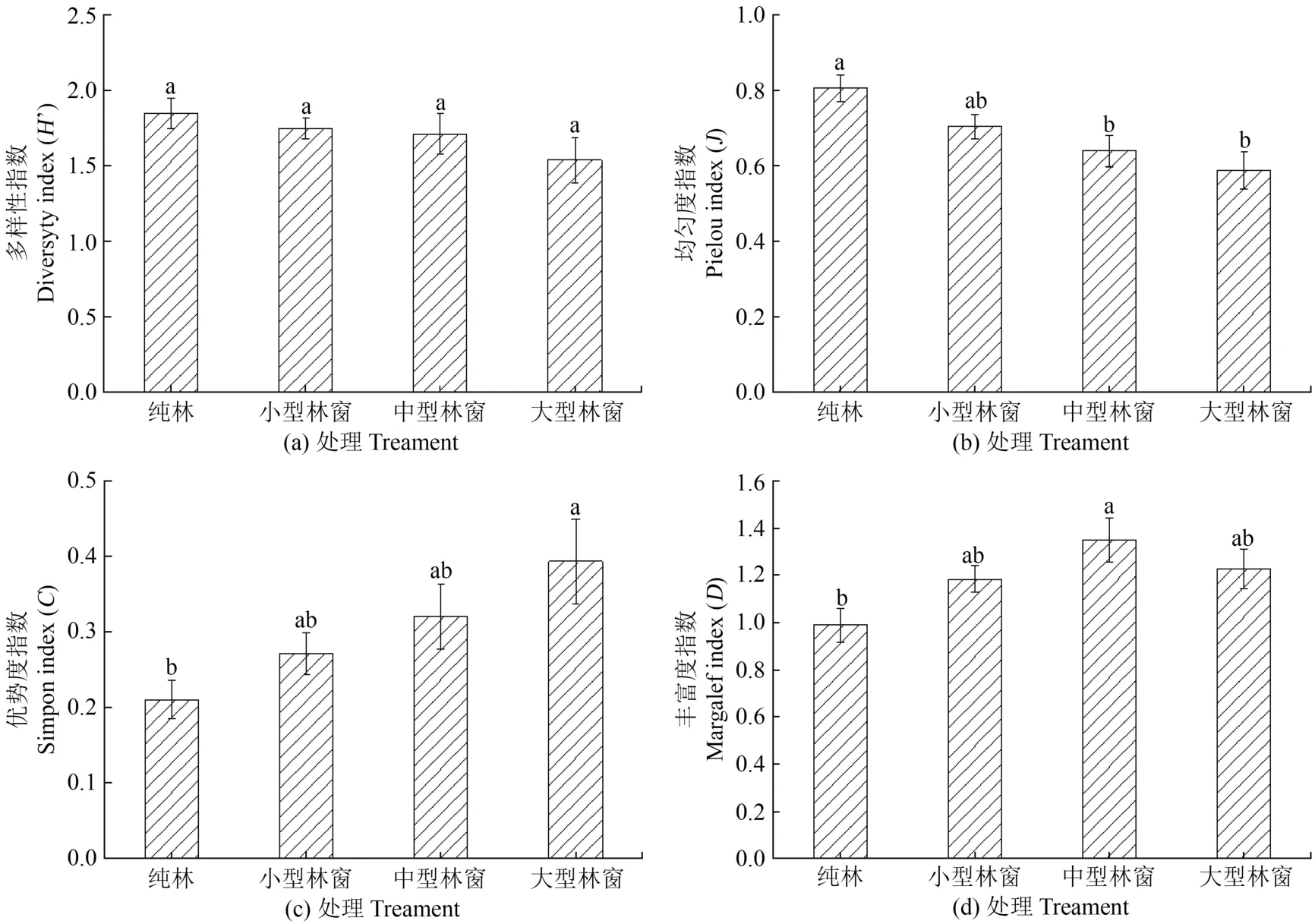
图3 中小型土壤动物群落特征指数(n=9,mean±SE)Figure 3 Micro-soil faunal characteristic index(n=9,mean±SE)
2.5 中小型土壤动物的功能类群
林窗改造下中小型土壤动物按照取食对象的差异性可分成植食性、杂食性、菌食性、腐食性和捕食性等5个营养功能类群。根据图4a可知,各林窗内以杂食性土壤动物为主,杂食性土壤动物对环境要求较低易生存,各样地数量占比在42.69%~65.72%之间,由此可见杂食性中小型土壤动物为川南马尾松人工林森林土壤动物的主体,随林窗改造面积的增大呈现出先减少后增加的趋势,在大型林窗达到最大值;其次为腐食性土壤动物,最高值出现在小型林窗,林窗面积越大,其所占百分比越小;由于中小型土壤动物食物以枯落叶为主,因此,腐食性土壤动物平均密度显著高于植食性土壤动物;植食性、菌食性和捕食性中小型土壤动物受林窗改造面积影响不明显。如图4b,林窗改造土壤动物功能群的类群数排序为腐食性>植食性>菌食性>捕食性>杂食性。其中,腐食性中小型土壤动物小型林窗达到最低值;纯林的菌食性、植食性中小型土壤动物类群数最低。
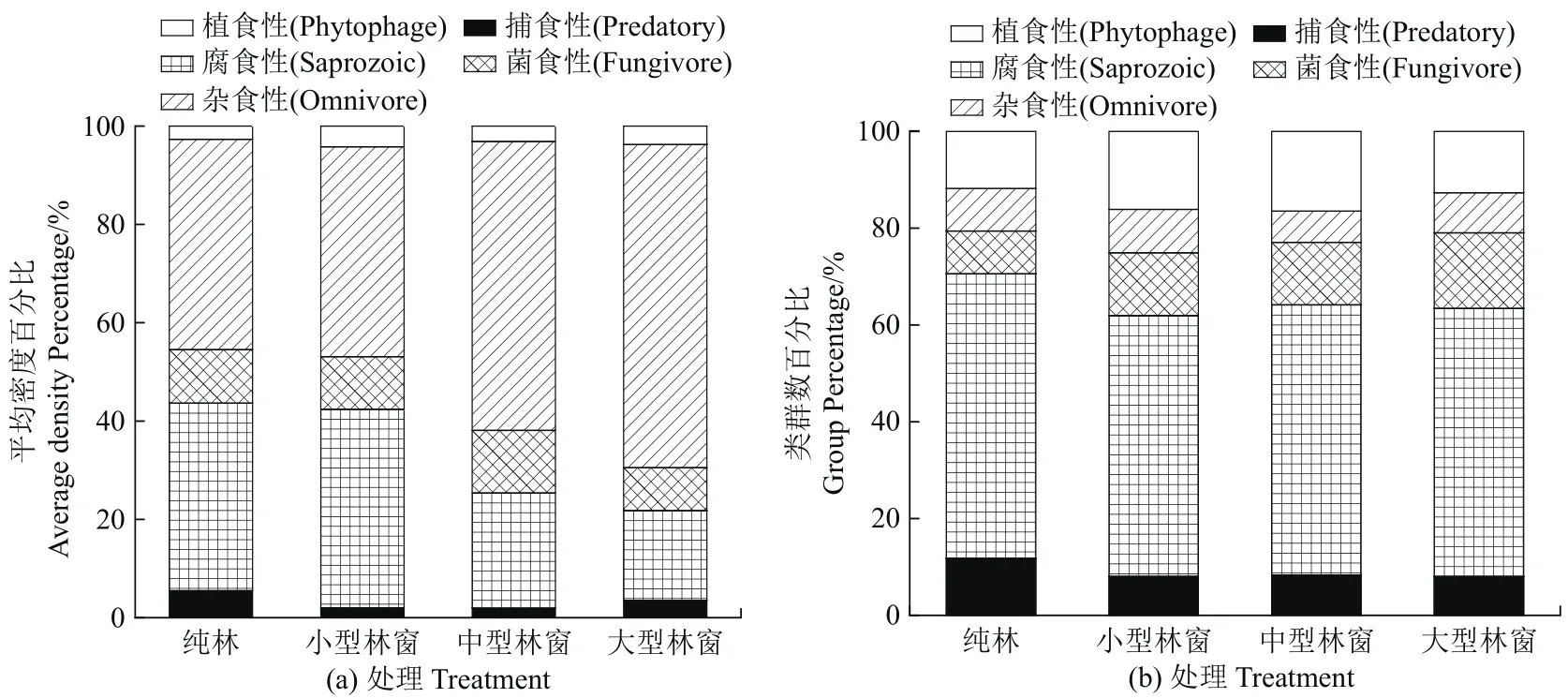
图4 不同林窗大小土壤动物功能类群百分比Figure 4 The percentage of functional group of soil fauna in different gap sizes
3 讨论
林窗作为森林生态系统常见的干扰形式之一,对森林生态过程有重要影响[16]。本研究表明:与纯林相比,改造后的林窗下土壤动物的平均密度和类群数均有所增加,这与潘业田[32]对柏木人工林林窗的研究结果一致。前期研究表明中等大小的林窗面积(625~900 m2)中马尾松人工林抚育初期物种丰富度最高,促进林下物种更新,改变林下光热条件,进而影响土壤理化性质,而土壤动物作为生态系统中的重要组成部分[33],对林窗所形成的环境变化尤其敏感,其群落结构和多样性的变化可反映环境中的细微变化[34]。线虫是土壤中最丰富的后生动物,占据土壤食物网的多个营养级。同时作为凋落物与土壤之间的桥梁,在调控生态系统的稳定发展、有机质分解以及植物矿化中发挥重要的作用[35-36]。各林窗内线虫纲土壤动物均为优势类群,共占总密度的50.37%,线虫纲土壤动物平均密度随林窗面积的增大呈现出递增的趋势,在大型林窗达到最大,林窗面积增大对线虫的数量产生一定的促进作用,中型和大型林窗内植被恢复较好,能为土壤线虫提供更多的食物来源以及土壤层的保湿作用,从而导致线虫的快速繁殖。在该样地中土壤动物的平均密度和类群数在不同面积的林窗中呈现出不一致的趋势,中型和大型林窗的平均密度显著高小型林窗和纯林,大型林窗平均密度最高,且大型和中型林窗的平均密度相差小;中型林窗类群数较高。林窗面积越大,林内受到的光照强度越大,影响林内的气温和相对湿度[37-38],从而对土壤动物产生影响,试验数据显示不同面积林窗对土壤动物平均密度的影响比土壤动物类群数的影响大。土壤动物平均密度随林窗改造面积的增大呈现出递增的趋势,与欧江[12]等研究的变化规律相似。土壤动物类群数在中型林窗数量最高,说明中等面积林窗(625~900 m2)更有利于土壤动物群落的发展。土壤动物的垂直分布存在显著表聚性[39],具体表现为随着土层的加深中小型土壤动物的密度降低,以0~5 cm土层的平均密度最大,这与苟丽琼等[40]研究结果类似。其中0~5 cm土层的土壤动物平均密度变化最明显,而凋落物层中小型土壤动物平均密度变动较小。林窗改造处理凋落物层土壤动物平均密度显著增长,这与林窗改造处理的林下植被有关,通过砍伐形成的林窗会使林下植被更为丰富,给予土壤动物丰足的食物来源,改善凋落物层土壤动物的生境环境,群落结构得到改善,明显增加中小型土壤动物数量。
土壤动物多样性能够反映出土壤动物群落组成的复杂程度及其群落结构的稳定性。土壤动物的群落多样性指数、均匀性指数、优势度和丰富度指数随林窗改造面积的改变,均呈现不同的变化趋势。在初期干扰过程中,土壤动物均匀度指数和优势度指数的变化呈相反趋势,物种均匀度来反映各物种种群数量的作用,优势度指数越大,表明生物群落内不同种类生物数量分布越不均匀,优势生物的生态功能越突出,物种数目越多,多样性越丰富,物种数目相同时,每个物种的个体数越平均,则多样性越丰富。而优势度指数的作用是反映各物种种群数量的变化情况。优势度指数越大,说明群落内物种数量分布越小均匀,优势种的地位越突出。实验结果表明,在大型林窗中土壤动物的均匀度指数最低,是由于林窗改造会改变群落或环境中的全部物种数目个体数目的分配状况。中度干扰假说提出,当群落在演替过程中受到中度水平干扰时,物种的多样性和丰富度最高[41]。土壤动物的多样性指数变化不明显、丰富度指数中型林窗最高。
土壤动物的生态作用是通过各功能类群间的食物网来实现的[42],自然界中的一种生物以多种生物为食来生存,占据多个营养级,由复杂的食物链形成食物网[43]。由于不同功能类群土壤动物对林窗干扰敏感程度不同,使得土壤动物的功能类群在分布上存在差异,导致群落结构发生变化。研究结果表明各样地土壤动物平均密度以杂食性土壤动物为主,因其拥有2~3种食性,占据绝对优势[44],除小型林窗以外其余样地杂食性平均密度占比均高于纯林样地。除杂食性土壤动物外,土壤动物最先的食物是以枯落叶为主的残体,经过植食性动物的分解,成为腐食性土壤动物的食物[45]。因此,腐食性土壤动物类群数显著高于植食性、菌食性和捕食性土壤动物。腐食性土壤动物受林窗改造的影响最明显,随面积增大类群数先减少后增加,说明林窗的形成对腐食性土壤动物的影响较大,对菌食性的影响较小。
4 结论
不同面积林窗改造使土壤动物平均密度和类群数均有所增加,在中等面积(625~900 m2)林窗中达到峰值,林窗内结构稳定,物种多样性高,更有利于土壤动物群落的发展。适度的干扰可以提高物种多样性及物种丰富度,而干扰强度超过一定限度时,就会破坏生态平衡,降低生物多样性。
