封育措施对呼伦贝尔草甸草原区草地生产力的影响*
2022-10-28聂莹莹辛晓平徐丽君杨桂霞
聂莹莹,辛晓平,徐丽君※,杨桂霞
(1.中国农业科学院农业资源与农业区划研究所,北京 100081;2.呼伦贝尔草原生态系统国家野外科学观测研究站,内蒙古呼伦贝尔 021011)
0 引言
草原是陆地生态系统的主体和重要资源,不仅是人类生存发展的重要生态屏障,更是优质畜产品的生产基地与生物遗传资源基因库[1]。近些年,随着生产发展和人口增加,人们对草地的掠夺逐渐加剧,导致草地发生不同程度的退化[2]。我国是草原资源大国,拥有各类草原总面积3.928亿hm2,其中内蒙古草原面积约8 670万hm2,是我国北方重要的绿色生态屏障和畜牧业生产基地[3]。呼伦贝尔是我国温带草甸草原分布最集中、最具代表性的地区,植被类型复杂多样,草原植物资源丰富,是欧亚草原的重要组成部分[4]。近年来,由于极端气候以及人为干扰的影响,草原已经发生了大面积的退化,覆盖度降低,生物多样性下降,生态功能减弱的现象,草原退化已经从20世纪的点状退化发展到21世纪的区域性的整体退化[5]。草原退化制约了畜牧业的发展,引起的土地荒漠化、水土流失和沙尘暴迭等生态环境问题,更是直接影响了社会经济的可持续发展。
目前,亟待解决的问题是抑制草原退化,恢复退化草原生态系统。为解决草原退化问题,国内外众多学者进行了多方面的研究工作,主要集中在合理化放牧[6]、围封[7]、刈割[8]、快速建植[9]等方面都取得了一定的进展。其中,围栏封育是一种有效而且简便易行的方法。围栏封育可以快速恢复植被,提高植被盖度[10]、增加物种丰富度[11],显著提升退化草地生产力[12,13],而且在一定程度上也可起到恢复土壤养分和水分[14]的作用。
围封年限是影响草地群结构征与物种多样性的重要因子[15]。Alice[16]等研究结果表明,围栏封育显著提高了生产力,并且使群落结构趋于稳定。崔雨萱等[17]认为短期封育利于退化蒿类荒漠草地生物量的积累,而对植物群落多样性的影响因地区不同而不同。李媛媛等[18]研究结果显示围栏封育有利于改善青藏高原退化高寒草地的植物群落结构,提高草地植物群落的盖度和生物量,促进其恢复演替。郑翠玲[19]等对呼伦贝尔沙化草地封育效果的研究中指出,封育年限的增加可使其群落趋于稳定,物种丰富度、均匀度和多样性都呈现增加趋势[20]。但也有研究显示,随着封育年限的延长,物种多样性呈现出降低的趋势[21]。通过以上分析得知,封育措施对群落结构以及植物多样性的影响较为复杂,不仅是不同类型草地间的差异,即使是同一类型的草地也会有不同的结论[22]。围栏封育是实现受损草地自我修复,维护生态系统平衡最经济、有效的方法之一。国内外诸多学者的研究结果也验证封育措施可以促进退化草原原有植被恢复及草地生态系统良性循环[23-28]。
文章通过对不同封育年限下草甸草原群落特征变化,深入研究草地生产力和物种多样性在封育过程中的变化规律,以探讨围栏封育对草甸草原群落特征的影响,进而为草甸草原生态系统的优化管理提供一定的理论依据。
1 研究区与数据
1.1 研究区
研究区位于内蒙古谢尔塔拉镇呼伦贝尔草原生态系统观测研究站(49°19′N,119°56′E),海拔666~680 m,属温带半干旱大陆性气候,年平均降水量350~400 mm,降水期多集中在6—9月。年平均气温1.5℃,最高和最低气温分别为36.2℃和-48.5℃;≥10℃年积温1 700~2 300℃,无霜期110d左右。草甸草原主要植物有羊草(Leymus chinensis)、麻花头(Serratula komarovii)、贝加尔针茅(Stipa baicalensis)和糙隐子草(Cleistogenes squarrosa)等,并伴以其他杂类草。土壤类型主要为淡栗钙土。
1.2 样地选择
研究区域内的植被类型为温性羊草草甸草原,于2006年开始进行了围栏封育处理。每年于7月份生长旺盛时节进行植被调查工作。该研究选择2008年、2009年、2010年、2012年、2014年、2015年和2018年的封育样地为研究对象(2015年该区遭遇了草原火灾,但被及时扑灭),同时在封育区相邻外围选取未封育区作为对照样地(该区自由放牧)。试验采用样方法,分别在围栏内外进行取样,测定植物地上生物量、盖度、密度以及地下生物量(其中,地下生物量的监测年份为2008年、2010年、2012年和2014年)指标,样方面积设置为1 m×1 m,围栏内外各取10个样方,共计样方20个。
1.3 测定指标及技术
1.3.1 测定指标
盖度:采用针刺法测定盖度。
密度:同一物种出现的株丛数。
地上生物量:将1 m×1 m样方内的植物分种齐地面剪下,装入信封袋中,并做好编号,带回实验室在(85℃)恒温下烘干12 h,称其干重。
地下生物量:在每个样方旁用直径10 cm的根钻,按照0~10 cm、10~20 cm、20~30 cm的深度钻取土柱,每个样方重复取3钻,用0~30 cm的根量表示该样方的地下生物量。将土柱放在0.5 mm网眼的纱网袋里,用水冲洗,将采集到的根系置于85℃的烘箱中烘干称重,计算地下生物量。
1.3.2 重要值计算


1.3.3 物种多样性计算
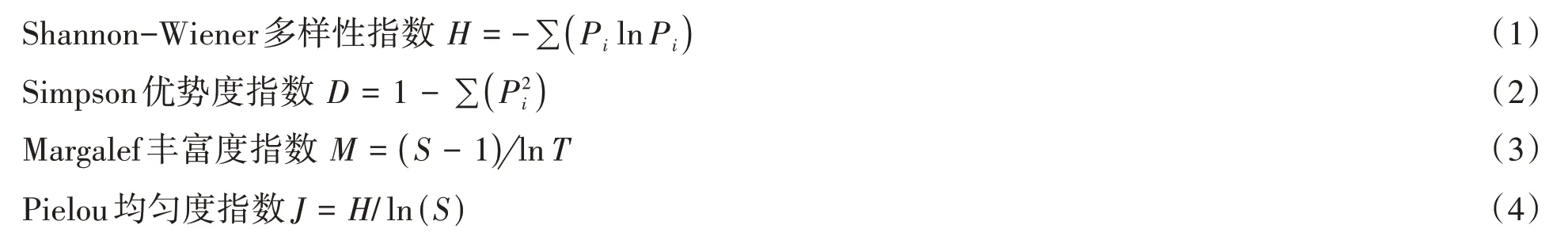
式(1)至(4)中,T为样方中的个体总数,S为样方内的物种数,Pi=Ni/N,Ni为物种i的相对重要值,N为植物种i所在样方的各个植物种的相对重要值总和。
1.4 数据分析
采用Excel 2013软件进行数据整理与作图,运用SPSS 20.0软件进行数据分析,物种多样性差异显著性采用单因素分析(One-Way ANOVA),并用Duncan检验进行平均值之间的多重比较,显著性水平设为P<0.05。
2 结果与分析
2.1 封育对群落地上植被数量特征和植物多样性的影响
表1方差分析结果显示,群落盖度、群落地上生物量和杂类草生物量对两个因素及其交互作用的响应均达到极显著水平(P<0.001);群落密度、多样性指数、Pielou均匀度指数和多年生禾草生物量对年际变化以及封育处理与年际变化的交互作用的响应均达到极显著水平(P<0.001),对封育处理的响应达到显著水平(P<0.05);Margalef丰富度指数对封育处理的响应未达到显著水平,但对年际变化及二者的交互作用的响应均达到极显著水平(P<0.001)。

表1 方差分析评估封育和年际变化及其相互作用对群落特征与植物多样性的影响
2.2 封育对群落结构的影响
植被高度、密度和盖度用来反应植物群落结构,该研究选取了盖度和密度来描述群落结构特征。从图1可知,封育显著提高了群落总盖度(P<0.05);各年际间变化,围栏内分别高出围栏外114.75%、62.81%、25.00%、50.49%、13.79%、98.19%和31.09%。从年际变化来看,群落盖度随着封育年限的增加表现出显著增加—降低—显著增加的变化趋势,2015年时群落总盖度最大,达到87.8%。围栏外自由放牧措施下的群落总盖度随着时间的增加呈现出先增加后降低的变化趋势。
封育措施对群落密度没有起到显著性作用,且在2012年和2018年围栏外的植被密度显著高于围栏内(P<0.05),分别比围栏内高出520.83%和474.60%。随着时间的增加,封育措施下群落密度整体表现出先增加后降低的变化趋势。

图1 封育对草甸草原群落盖度和密度的影响
2.3 封育对群落生物量的影响
封育显著增加了群落地上生物量,各年际间围栏内较围栏外分别增加224.16%、132.90%、66.00%、176.31%、309.19%、887.30%和182.72%;从年际间变化来看,封育措施下的群落植被地上生物量是随着时间的增加呈现字母“M”型的变化,即先显著增加后显著降低再显著增加降低的波动性变化(P<0.05),2015年达到了峰值,为310.44g/m2;围栏外的自由放牧群落植被地上生物量随着时间的增加呈现出先显著增加后降低的变化趋势,且在2015年值最小,仅为封育的1/10,峰值出现在2019年,但也只有88.97g/m2(图2)。封育措施显著提高了地下生物量,各封育年限地下生物量分别比同年度的围栏外的地下生物量高出了25.24%,143.98%,89.02%和20.47%,2010年和2012年围栏内外地下生物量差异显著(P<0.05);地下生物量随着封育年限的延长表现出先增加后减小的变化趋势,2010年群落地下生物量值最大,比封育2008年、2012年和2014年地下生物量分别高出42.44%,17.56%和126.67%。群落中的枯落物重量随着封育时间的增加呈现出逐渐增加,但由于2015年草地经历了火灾,累积的枯落物被烧掉,值为0,在火灾后3年的2018年枯落物又有所增加(图2)。封育也显著提高了多年生禾草生物量。其中封育措施下的群落多年生禾草生物量随着时间的变化趋势与地上生物量的变化趋势一致,也是“M”型的变化,即先增加后降低再增加降低,2015年出现了峰值,禾草生物量达到184.88g/m2;围栏外的禾草生物量随着时间的增加整体呈现出逐渐降低的变化趋势,2015年出现了最小值,仅为4.06g/m2(图2)。
2.4 封育对群落多样性的影响
该研究选取了Shannon-Wiener多样性指数、Simpson优势度指数、Margalef丰富度指数和Pielou均匀度指数来描述群落多样性的变化。从表2可知,2012年和2018年围栏内的Shannon-Wiener多样性指数分别比围栏外显著提高了45.24%和44.63%(P<0.05),但比2009年围栏外低了8.57%,比2014年低了18.46%(P<0.05),其余年际,封育虽然提高了多样性指数,但效果不显著(P>0.05)。2012年和2018年围栏内的Simpson优势度指数比围栏外显著提高了27.99%和30.87%(P<0.05),但在2009年和2014年却比围栏外降低5.29%和8.94%。Margalef丰富度指数在2012年、2015年和2018年围栏内比围栏外分别提高了90.24%、24.22%和50.24%,差异达到显著水平(P<0.05),2008—2014年分别比围栏外低了3.15%、18.75%、2.10%和30.79%。围栏内的Pielou均匀度指数在2010年、2012年、2015年和2018年分别比围栏外提升了0.48%、23.51%、3.07%和36.62%(P<0.05),2008—2012年分别比围栏外降低了1.37%、1.86%和9.90%(P<0.05)。从年际变化来看,多样性指数、丰富度指数和均匀度指数均表现出相同的变化趋势,即围栏内的指数随着监测年份的增加表现出在封育初期(2008—2010年)时表现出逐渐增加,封育后期则表现出显著降低的变化趋势(P<0.05);而围栏外表现出增加—降低—增加—降低的波动变化。
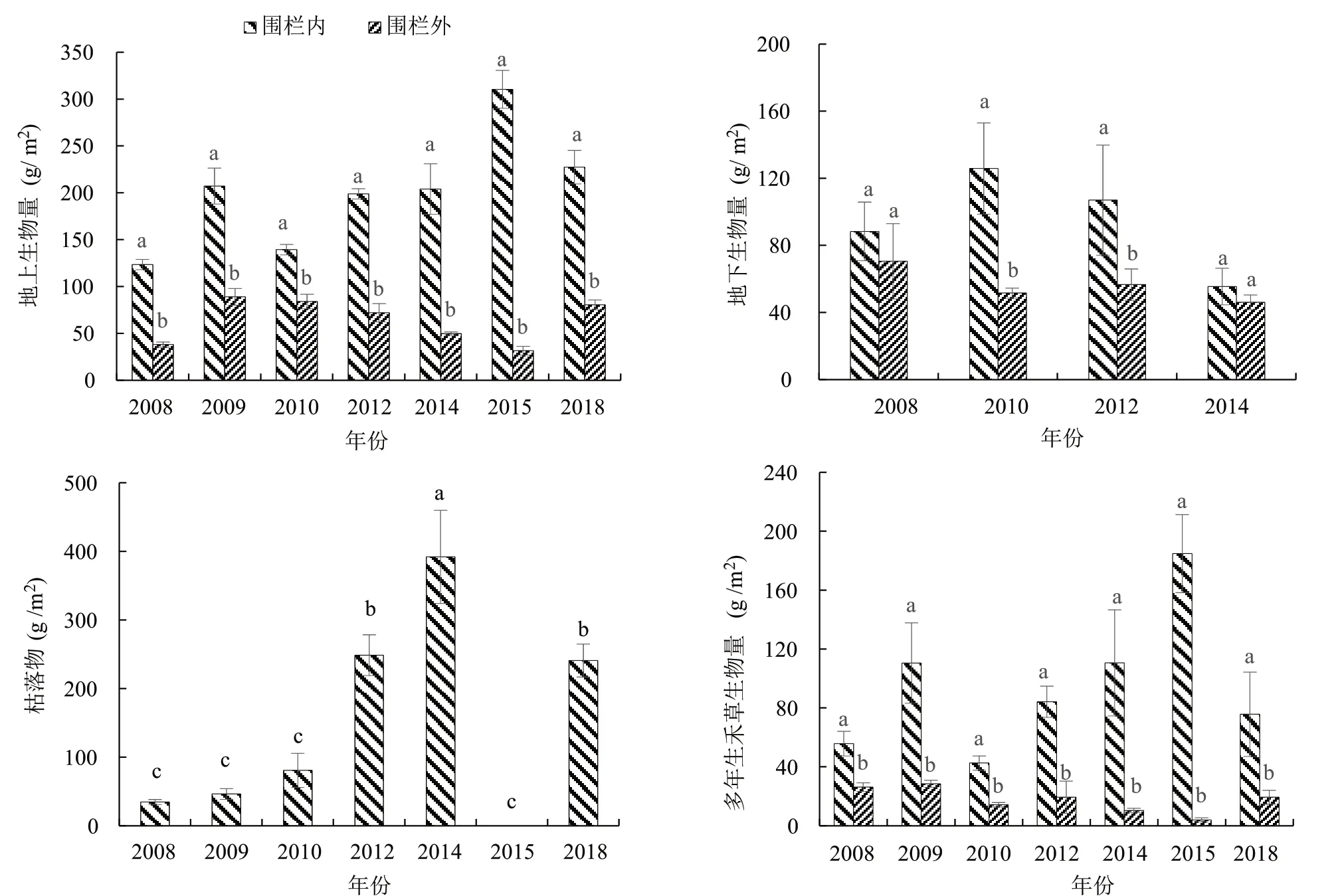
图2 封育对群落植被生物量的影响
3 结论与讨论
3.1 围封年限对群落数量特征的影响
盖度和密度从侧面反映了绿色植物光合能力的强弱、群落覆盖的繁茂程度以及经济性状的大小[29],是草地植物群落的重要的数量特征[30];其中,植被盖度的增加是植被恢复的重要指标,能够显著减少水土流失和改善地表小气候。围栏封育作为修复退化草地的有效措施被广泛采用,形成了良好的生态效益。主要表现在封育可以显著增加植被的盖度与高度。实验结果表明,封育显著增加了植物盖度;封育后的草地,由于没有受到牲畜啃食,人文活动等,植被能够正常发育—开花—结实,完成了整个生育周期,且植被根茎新生枝条增多,促使围栏内的群落盖度相较于围栏外的自由放牧地显著提高。围栏外的自由放牧地由于牲畜的选择性采食,一些适口性好的高大型禾草如羊草等是牲畜最为喜欢的植物,使高大型禾草在围栏外的竞争减弱,使群落中的一些低矮型植物优势地位上升,具有一定的生长空间,使群落密度增加。这与前人的研究结果一致[31]。

表2 围栏封育对植物多样性的影响
3.2 围封年限对植物多样性的影响
植物多样性是维持草地生态系统发展和生产力的基础[32],关于植物多样性对封育年限的响应,在前人的研究中的结论不尽一致。比如,宋明华[33]等研究表明围栏封育增加了物种多样性,赵菲[34]的研究表明围栏封育使物种多样性降低,而Meissner[21]等的研究结果显示封育对物种多样性没有显著性影响。究其原因主要是草地类型或生境、封育时间以及放牧强度等差异所造成的。因此,封育与物种多样性的关系很难一般化[35]。就本试验而言,封育措施可使草甸草原群落物种多样性有一定程度的提高,但并不是所有的监测年份下都有所提升;比如2009年和2014年的Shannon-Wiener和Simpson指数就表现出围栏外高于围栏内。随着封育年限的增加,群落中的禾草显著增加,禾草在水和养分竞争中占有较大的优势,抑制了其他植物的生长,进而使植物多样性降低。Margalef丰富度指数在2008—2010年以及2014年时表现出围栏外好于围栏内,而在2012年、2015年和2018年又显著低于围栏内的指数值。从物种丰富度指数公式中可以看出,群落密度越大,其丰富度指数值就越小,封育前期,由于封育避免了不利因素的干扰,草甸草原植物得以较好的恢复以及繁殖更新,密度较围栏外有所增加(图1);随着封育年限的延长,优势种补偿性生长,种间竞争力增强,导致其他低矮型植物种逐渐减少甚至消失,所以围栏内的密度较围栏外显著减少,从而导致了Margalef丰富度表现出封育初期围栏内高于围栏外的指数值,到了封育后期表现为围栏外高于围栏内的指数值。
3.3 围封年限对草地生产力的影响
生物量是指植物(地上现存量和地下根系)生长发育所产生的物质总量,这不仅是生态系统生产力的重要标志,也是整个生态系统运行的能量基础和营养来源[36-38],地上生物量是群落生产力水平的重要功能指标,反映了群落在特定时期内积累有机物的能力[39]。地下生物量是指存在于地下根和根茎生物量的总量,是草地土壤碳库最主要的输入源,在草地碳循环中起着关键作用[40]。该实验研究结果显示,围栏封育显著提高了地上生物量、地下生物量以及禾草生物量(P<0.05)。这一结果与毛绍娟[41]等对藏北高寒草原群落特征和贾志峰[42]等对退化高寒草甸植被特征对封育响应的研究一致。进一步说明封育是提高草地生产力、恢复退化草地的重要管理手段。
诸多研究证明了封育措施显著提升了草地生产力,但是根据草地生态系统的可持续原理,草地围封具有一定的阈值[36]。封育时间过长,不利于牧草的正常生长和发育,长期累积的枯落物抑制植物的再生和幼苗的形成,导致草地的繁殖更新缓慢。而且,有研究证实,适宜的刈割或者放牧干扰可以改良草地质量,刺激牧草分蘖,促进牧草再生[43]。地下生物量/地上生物量在研究陆地生态系统中碳、氮分配和储存时起着重要的指示作用[44]。围栏封育对草地的影响一是直接减少牲畜的采食和践踏,另一个是通过牲畜对土壤结构及土壤水分的干扰而间接影响着植物;围栏封育后的退化草地地上部分繁衍得生息得意恢复,可以为根系生长提供较多的营养物质、养分含量和浅层土壤水分状况明显改善,为根系的生长发育提供较为优越的环境,有利于地下部分的生长发育,所以围封初期增加了地下生物量。但由于草原植物生物学特性及环境资源有限的制约,围封后地下生物量累积也不是无限制的。试验结果显示,2010年时,地下生物量达到峰值,后随着封育时间的增加逐渐降低。其原因可能是地下根系的能量一部分维持自身的生长,另一部分通过微生物分解将能量转化在土壤中,封育5年后的草地,地上生物量获取除了光合作用的能量外,还获取了地下部分的能量,而且地下部分自身能量并未能够得到及时补充,从而导致封育年限增加,地下生物量反而降低。该实验的研究侧重点集中于讨论封育年限对草地生产力的影响,关于根部养分流失、植被、根系以及土壤中的碳、氮之间的转化机制对封育年限的响应,仍然需要更进一步的研究。
3.4 火烧干扰对封育草地生产力的影响
实验结果显示,随着封育时间的延长,地上生物量基本保持在200g/m2左右,增加率一度趋于平缓,直到2015年突增到310g/m2,这是因为2015年封育的草地遭遇了草原火灾,群落中累积的枯落物被消除,增加了群落内部的光资源,并且增加了土壤养分、土壤微生物活动,从而增加了草地生产力[45]。这一结果与前人研究相同[46]。但也有不同的结论,刘健[47]等对半干旱黄土区草地火烧干扰对草地生产力的研究结果显示,火烧显著降低了植物地上生物量。也有研究结果表明,火烧对地上生物量没有影响[48]。由此可见,火烧对草地的影响研究结果不尽一致,其内在机制仍然需要进一步研究。该研究中的退化草地在长期封育后,草地生产力增长滞缓,在经历过草原火灾干扰后,生产力得到了显著性增加,由此说明,退化草地长期封育后应该进行适度利用,可使草地生态系统的能量流动和物质循环保持良性状态,进而保持草地生态系统平衡。火烧干扰虽然提高了封育草地生产力,但却不提倡通过火烧干扰来改善草地质量,因为不能保证计划用火对生态安全不存在任何潜在的威胁,计划用火的生态风险依然存在。而且研究也表明,长期封育的草地应运用适度的放牧或者刈割来改良草地质量,刺激牧草分蘖,促进牧草再生草地生态系统平衡[36,49]。
