长江禁捕后长江口刀鲚资源特征
2022-10-25马凤娇杨彦平方弟安应聪萍殷国俊
马凤娇 杨彦平 方弟安, 应聪萍 徐 跑, 刘 凯, * 殷国俊, *
(1.南京农业大学无锡渔业学院, 无锡 214081; 2.中国水产科学研究院淡水渔业研究中心,农业农村部淡水渔业和种质资源利用重点实验室, 无锡 214081)
河口是淡水和海洋生态系统的过渡区域, 其特殊生境对河口定居性鱼类和洄游性鱼类有着重要的生态意义[1—3]。长江口是我国最大的河口生态系统和典型的生态群落交错区, 具有独特的地理条件和生态环境, 孕育了丰富的渔业资源, 是刀鲚(Coilia nasus)、鳗鲡(Anguilla japonica)等重要鱼类溯河和降海洄游的必经通道, 也是中华绒螯蟹(Eriocheir sinensis)等半咸水水生生物重要的栖息和繁殖场所,对淡水和海水渔业资源有着重要的支撑作用。
刀鲚又名长颌鲚, 俗称刀鱼, 隶属于鲱形目(Clupeiformes)、鳀科(Engraulidae)、鲚属(Coilia),是一种中小型经济洄游型鱼类[4—9]。刀鲚主要分布于太平洋的西北部和西部地区, 包括中国的黄渤海、东海近海沿岸水域及通海的江河如长江、黄河、淮河及钱塘江等[10], 其中以长江流域的产量最高, 禁捕之前曾是长江下游及长江口水域重要的捕捞消费对象[11]。刀鲚生殖洄游群体于每年春季聚集于长江口, 集群溯江而上进入长江中下游及湖泊中繁殖[12,13]。历史上, 刀鲚曾为长江口水域最重要的捕捞对象之一, 1973年最高捕捞量达392×103kg。但在生态环境持续恶化及高强度长期捕捞等多重因素影响下, 资源量急剧衰退并且逐年下降, 捕捞量锐减至2016年的3.7×103kg, 与最高捕捞量相比,降幅达99.06%, 渔汛几近消失[14,15]。
为了保护和恢复长江刀鲚资源, 农业农村部于2018年12月29日发布《农业农村部关于调整长江流域专项捕捞管理制度的通告》, 规定自2019年2月起, 全面禁止刀鲚天然资源生产性捕捞, 这对遏制刀鲚资源快速衰退、从根本上保护和恢复刀鲚资源至关重要。为巩固和扩大长江禁捕退捕成效,加强长江口水域禁捕管理, 农业农村部于2020年11月20日发布《农业农村部关于设立长江口禁捕管理区的通告》, 决定扩延长江口禁捕范围(122°15′E、31°41′36″N和30°54′N), 设立长江口禁捕管理区。规定自2021年1月1日0时长江口禁捕管理区以内水域, 实行长江流域禁捕管理制度。为跟踪评估刀鲚生产性捕捞退出后其资源恢复效果, 本研究自2019年以来持续开展长江口刀鲚资源调查, 并结合刀鲚生产性捕捞退出前(2017—2018年)的调查成果, 以期掌握长江口刀鲚生物学特征、资源密度及其时空特征等资源参数, 探索生产性捕捞退出及长江流域重点水域全面禁捕后长江口刀鲚种群结构及资源现状, 为刀鲚资源保护及禁捕效果评估积累基础数据。
1 材料与方法
1.1 调查水域与调查时间
长江刀鲚的生产性捕捞已于2019年正式退出,生产性捕捞退出前(2017—2018年)每年在长江口刀鲚捕捞汛期内, 随机从持有刀鲚捕捞许可证的生产船上采集刀鲚样本。生产性捕捞退出后(2019—2021年)每年于刀鲚洄游汛期内(3—6月), 利用中国水产科学研究院淡水渔业研究中心自有科研调查船, 并经上海市农业农村委员会批准, 在长江口崇明水域设置3个调查样区(A水域, 中心经纬度为E121°29′50″, N31°31′08″; B水域, 中心经纬度为E121°35′56″, N31°30′18″; C水域, 中心经纬度为E121°39′49″, N31°30′31″; 图1)。2017—2021年调查网具均为抛定刺网, 网目尺寸4 cm, 网长150 m,网高12 m, 调查期间每天实际捕捞约2.5h (低潮位时放网, 涨至平潮时收网), 同步采集调查样区水温、水深等环境信息(表1)。调查时段内, 除台风等恶劣天气外, 每天开展刀鲚调查。

表1 2019—2021年长江口不同调查时段平均水温变化Tab.1 The mean water temperature at different sampling periods in the Yangtze River estuary from 2019 to 2021
图1 长江口刀鲚调查样区示意图Fig.1 Sampling regions for investigation of Coilia nasus in the Yangtze River estuary
1.2 样品采集
采集的刀鲚样本全部进行全长、体长和体质量等表观生物学测量, 精确到1 mm和0.1 g, 并统计刀鲚的调查总重量和尾数。同时随机抽取部分样本进行解剖, 目测法鉴定其性别、性腺发育情况,按照Ⅰ—Ⅵ期进行划分[16—18]。
1.3 数据处理和分析
由于体长的全距(最大体长与最小体长的差值)较分散, 为便于分析, 对所测定的刀鲚以20 mm为组距进行分组, 分析其长度组成频率。
刀鲚按照体质量定义体型规格, 体质量≥100 g为大规格刀鲚, 75—99 g为中等规格刀鲚, 体质量≤75 g为小规格刀鲚。
采用SPSS 24.0软件拟合体长与体质量之间的幂函数关系:W=aLb,W为体质量(g),L为体长(cm),a、b为参数, 其中a值表示单位体长时鱼的体质量,b值可反映鱼体的生长特征, 同时, 配对T-检验确定参数b与3的显著性差异; 根据K=(W/L3)×100计算丰满度(Condition factor),W为体质量(g),L为鱼的体长(cm),K为丰满度(%)。
单位捕捞努力量(Catch Per Unit Effort, CUPE)是反映资源的丰度变化的重要指标, 单船各时段、各水域刀鲚样本重量以CPUE/kg来表征, 将采集的刀鲚尾数NB和重量WB标准化为日均调查尾数(尾/d)和重量(kg/d), d为汛期实际作业天数。用Nt、Wt分别表示单网调查尾数(尾/网)和重量(kg/网)。
采用Excel和SPSS Statistics 24软件处理数据,计算体长、体质量、丰满度和资源密度参数的均值(mean±SD), 并分析其时间和空间差异, 用t检验和单因素方差分析进行差异显著性分析, 组间差异显著性使用LSD检验, 显著水平P<0.05, 极显著水平P<0.01。其中统计刀鲚体长和体质量、丰满度及资源密度的时空特征用2019—2021年的数据; 统计刀鲚体长和体质量、丰满度变化和繁殖生物学特征的年际特征用2017—2021年的数据。
2 结果
2.1 体长、体质量的变化
时间特征在专项捕捞退出后, 2019—2021年共采集6105尾刀鲚样本, 体长变幅为103—388 mm,均值为(262±40) mm, 优势体长组为(260±60) mm,占群体总数的79.57%—92.40%; 体质量变幅为3.2—256.7 g, 均值为(78.7±36.5) g (图2A)。0—75 g的小规格刀鲚比例由2019年的63.92%减少至2020年的57.64%和2021年的28.98%, 而大规格刀鲚在2021年大幅增加, 相比2019年和2020年分别上升18.24%和18.52% (图2B)。专项捕捞退出前, 2017—2018年共采集刀鲚342尾, 体长变幅为119—381 mm,均值为(257±51) mm, 体质量变幅为4.9—206.8 g,均值为(74.4±42.2) g (表2)。由ANOVA分析可得, 生产性捕捞退出前(2017—2018年)后(2019—2021年)平均体长和平均体质量差异显著(P<0.05), 且2019—2021年显著大于2017—2018年(P<0.05)。
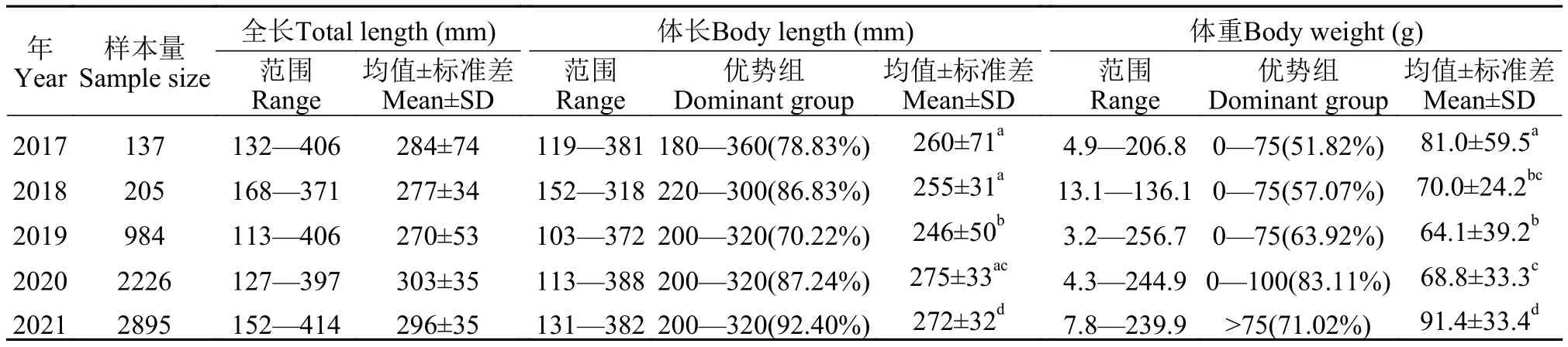
表2 2017—2021年长江口刀鲚生物学参数年间比较Tab.2 The comparison of biological parameters of Coilia nasus in the Yangtze River estuary from 2017 to 2021

图2 2017—2021年长江口刀鲚体长与体质量分布比例Fig.2 Percentages of (A) body length, and (B) body weight for Coilia nasus in the Yangtze River estuary from 2017 to 2021
2019年和2020年刀鲚样本平均体长和平均体质量随着调查时间的后延, 总体呈显著上升的趋势(P<0.01)。与之相反, 2021年两项生物学指标随调查时间后延呈逐渐下降的趋势, 且显著大于2019—2020年(图3)。

图3 2019—2021年长江口刀鲚体长和体质量均值时间特征Fig.3 The temporal characteristics of mean body length and mean body weight of Coilia nasus in the Yangtze River estuary from 2019 to 2021
空间特征2019—2021年在刀鲚洄游汛期内3个调查样区水深信息见表3。同年不同水域平均体长和平均体质量均存在显著差异(P<0.05), 以2020年A水域的平均体长为最大, 平均为(275±36) mm, 2021年B和C水域次之, 2019年C水域的最小, 平均仅为(233±52) mm。2019—2020年不同水域体质量波动十分明显, C水域显著小于A和B水域(P<0.01), 而2021年C水域显著大于A和B水域(P<0.01; 图4)。
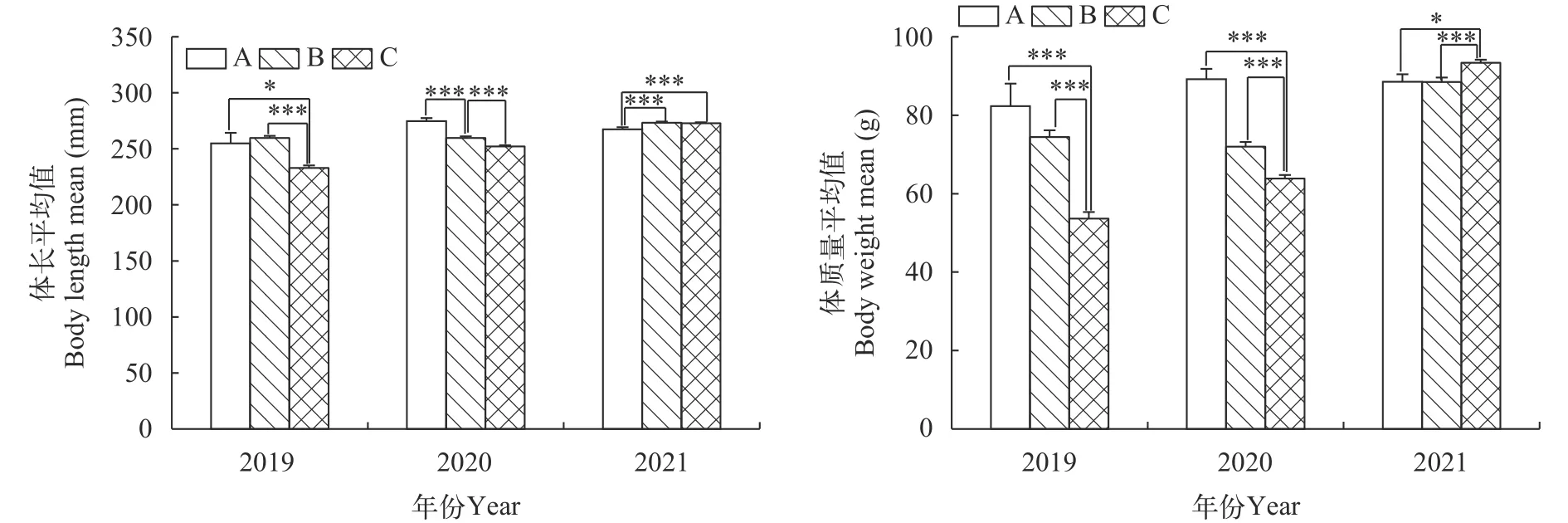
图4 2019—2021年长江口刀鲚体长和体质量均值空间特征Fig.4 The spatial characteristics of mean body length and mean body weight of Coilia nasus in the Yangtze River estuary from 2019 to 2021
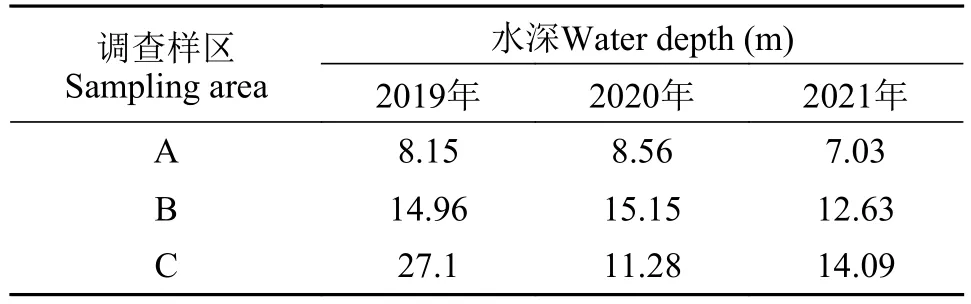
表3 2019—2021年长江口刀鲚调查水域水深信息Tab.3 Water depth of sampling area of Coilia nasus in the Yangtze River estuary from 2019 to 2021
体长—体质量关系根据2019—2021年长江口汛期刀鲚体长L(cm)与体质量W(g)数据, 在不考虑性别的情况下, 分别拟合出不同年份体长—体质量的幂函数关系式(图5), 其关系式为:
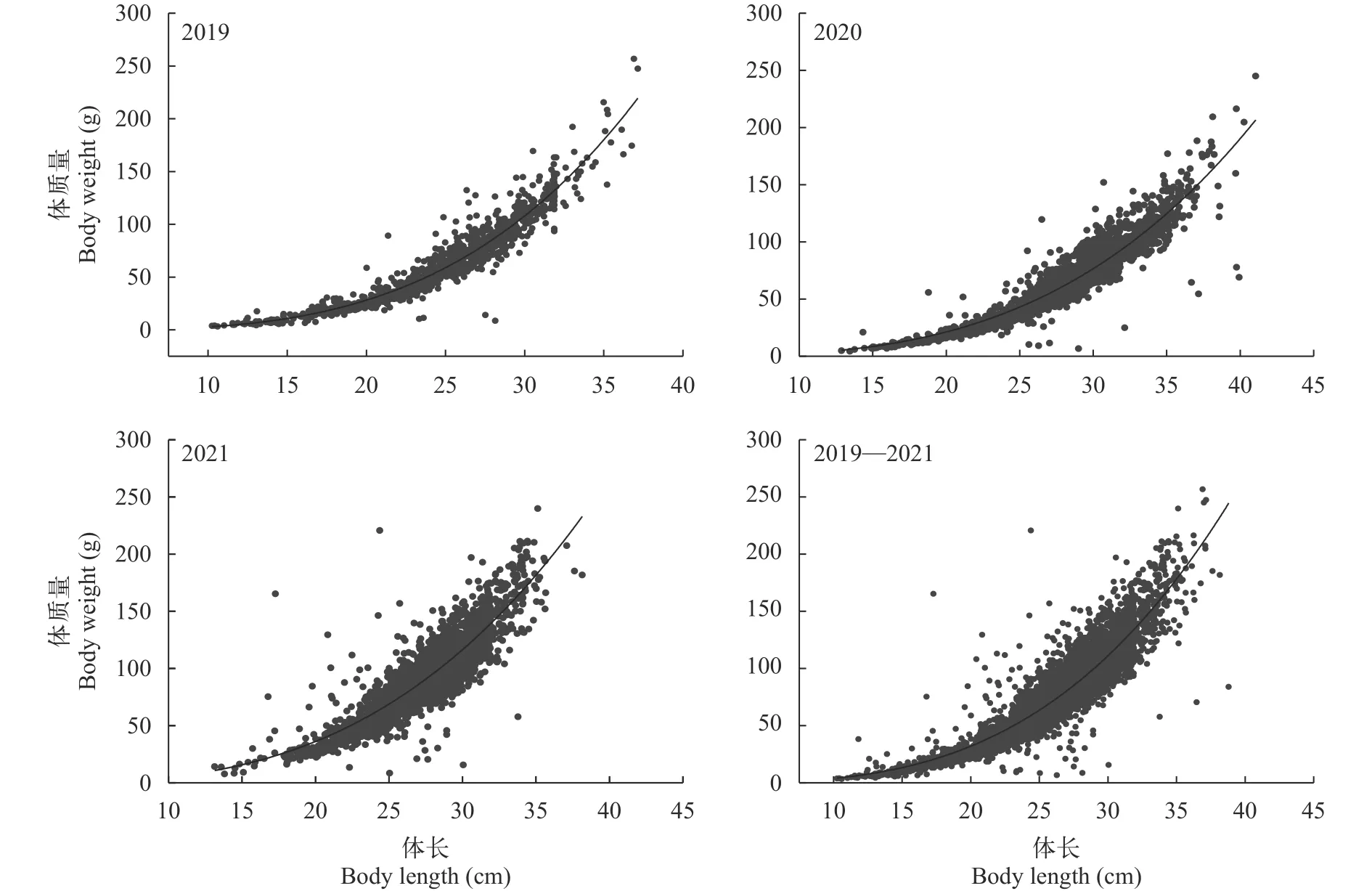
图5 2019—2021年长江口刀鲚体长与体质量关系Fig.5 The relationship between body length and body weight of Coilia nasus in the Yangtze River estuary from 2019 to 2021
2019年:W=0.001×L3.319(R2=0.910,n=984)
2020年:W=0.001×L3.164(R2=0.882,n=2228)
2021年:W=0.006×L2.884(R2=0.781,n=2895)
2019—2021年:W=0.003×L3.065(R2=0.840,n=6105), 经配对T检验, 参数b与3存在显著差异(P<0.01), 且b>3, 刀鲚的生长呈正异速生长。
2.2 丰满度的变化
年际分析显示, 长江口刀鲚繁殖群体的平均丰满度系数在各年间差异显著(ANOVA,P<0.05),2019—2021年平均丰满度系数为0.40±0.09, 较2017—2018年(0.39±0.06)略有增长, 但未达到显著水平(ANOVA,P>0.05)。时间特征显示, 丰满度系数在相同年份不同时段之间均存在显著性差异(ANOVA,P<0.05)。空间特征显示, 同年各水域间丰满度存在显著性差异(ANOVA,P<0.05; 图6)。

图6 2019—2021年长江口刀鲚繁殖群体的丰满度变化Fig.6 The condition factor changes of breeding population of Coilia nasus in the Yangtze River estuary from 2019 to 2021
2.3 繁殖生物学特征
性比和性腺发育2019—2021年随机抽样解剖4626尾刀鲚样本, 其中雌鱼3262尾, 雄鱼1364尾, 雌雄比例为2.39∶1, 与理论值1∶1存在极显著差异(P<0.01)。2017—2021年刀鲚的雌雄比分别为2.17∶1、1.97∶1、2.17∶1、3.25∶1和1.87∶1。雌雄个体性腺发育I—Ⅵ期的样本均有发现, 卵巢和精巢的发育期以Ⅰ—Ⅲ期为主(表4)。
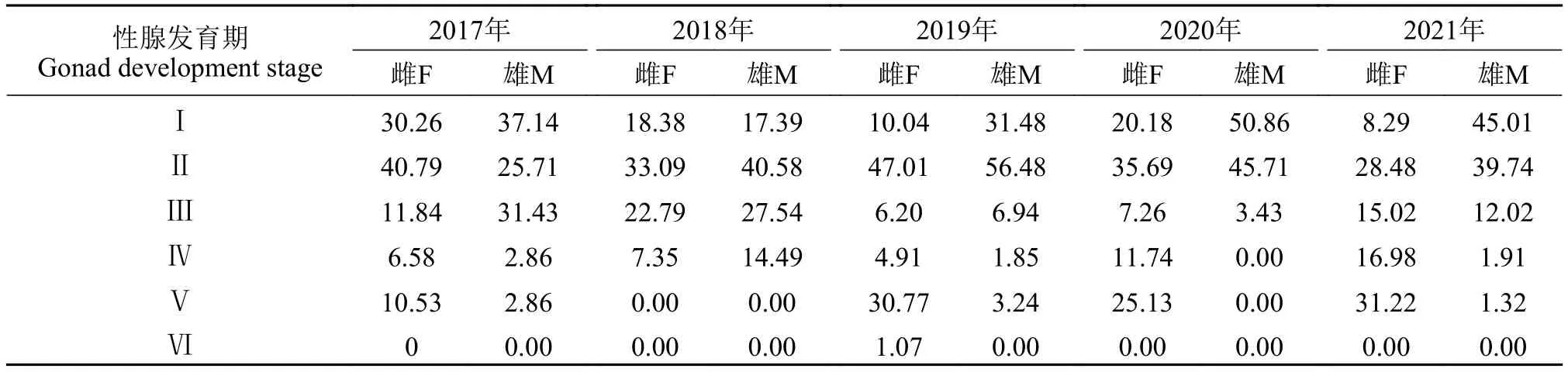
表4 2017—2021年长江口刀鲚繁殖群体性腺发育期比例Tab.4 The percentage of gonad development stages of breeding population of Coilia nasus in the Yangtze River estuary from 2017 to 2021
时空特征在洄游的早期阶段(3—4月),2019—2021年长江口刀鲚雌雄性腺发育期为Ⅰ—Ⅲ期的比例均大于97.34%; 随洄游时间的后延, 性腺发育成熟个体比例增加, 第3、4时段(5—6月)卵巢发育期在Ⅳ以上的雌性个体占比在66.67%—81.63%, 而发育至Ⅴ期的个体比例高至43.42%以上;而雄鱼精巢发育期在Ⅳ—Ⅴ期的比例也呈增加的趋势, 第4时段比例最高。从不同采样水域来看, 雌雄鱼的性腺发育没有规律性变化, 除2020年所有水域雄鱼个体的精巢发育期为Ⅰ—Ⅲ, 其他水域基本都有性腺发育到Ⅰ—Ⅴ期的个体(图7)。

图7 2019—2021年长江口刀鲚繁殖群体性腺发育期时空特征Fig.7 The temporal and spatial characteristics of gonad development stages of breeding population of Coilia nasus in the Yangtze River estuary from 2019 to 2021
2.4 相对资源密度及其时空特征
时间特征2019—2021年调查时间为3月中旬至6月底, 作业天数变幅为17—22d, 平均20d; 获得的刀鲚样本总重量变幅为63.07—264.51 kg, 平均为160.24 kg; 日均重量变幅为2.87—15.56 kg/d, 平均为8.70 kg/d; 2020年采集到的刀鲚样本重量较2019年增加了2.43倍,NB、WB、Nt和Wt较2019年分别增加了148.84%、166.90%、146.46%和164.86%;2021年刀鲚样本重量较2019年和2020年分别增加了4.19倍和1.73倍,NB、WB、Nt和Wt较2020年分别增加了53.00%、103.13%、74.88%和132.65%。长江口汛期刀鲚资源密度各参数变化显示, 各年份间刀鲚样本重量、NB、WB、Nt和Wt具有一致的变化趋势, 呈逐年上升的趋势(ANOVA,P<0.05; 图8)。
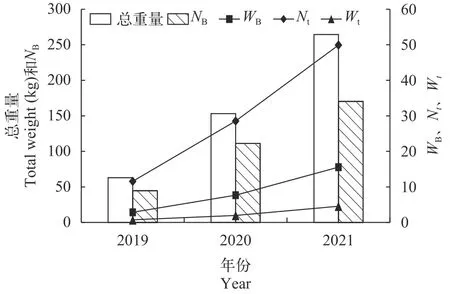
图8 2019—2021年长江口刀鲚资源密度变化Fig.8 The resource density change of Coilia nasus in the Yangtze River estuary from 2019 to 2021
2021年刀鲚洄游的第1时段水温较低[(13.59±0.48)℃], CPUE (104.97 kg)分别是第2、第3和第4时段的1.9倍、1.19和6.65倍, 较2019年(第3时段)最高值增加5.05倍, 但各时段NB、WB、Nt和Wt差异不显著(ANOVA,P>0.05)。2020年CPUE的变化与2021年相似, 第1时段CPUE最高, 较2019年各时段增加了2—5倍。2019年各时段WB、Nt和Wt存在显著性差异(表5)。从图9可以看出刀鲚各时段单日调查量随水温变化未呈现出一定的变化规律, 2020年和2021年刀鲚主要集中在3—4月洄游,而2019年则集中在5月份, 洄游高峰期显著提前。
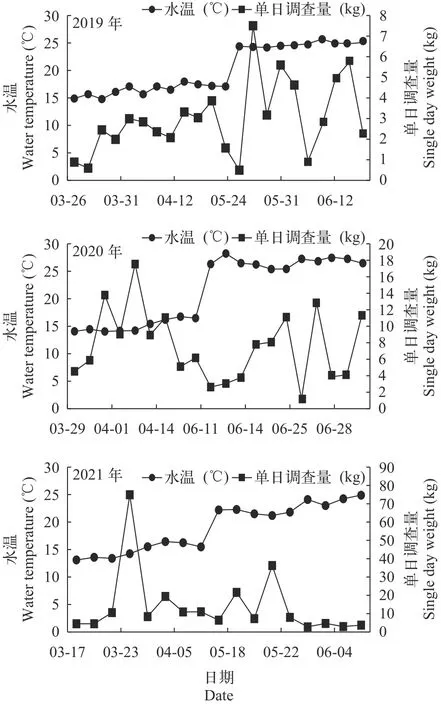
图9 2019—2021年长江口刀鲚汛期单日调查量与水温的关系Fig.9 Relationships between single day weight of Coilia nasus and water temperature in the Yangtze River estuary from 2019 to 2021

表5 2019—2021年长江口刀鲚资源密度的时间特征Tab.5 Temporal characteristics of resource density of Coilia nasus in the Yangtze River estuary from 2019 to 2021
空间特征从不同调查水域来看, 2021年C水域刀鲚样本的CPUE分别是A水域和B水域的5.42倍和2.14倍, 各水域间NB和WB不存在显著差异,而C水域的Nt和Wt均显著大于A水域(ANOVA,P>0.05)。2020年C水域的NB、WB、Nt和Wt均显著大于其他两个水域, 2019年C水域的NB和WB显著高于A水域(表6)。

表6 2019—2021年长江口刀鲚资源密度的空间特征Tab.6 Spatial characteristics of resource density of Coilia nasus in the Yangtze River estuary from 2019 to 2021
3 讨论
3.1 长江口刀鲚洄游群体的生物学特征
刀鲚曾是长江下游传统的重要捕捞对象, 20世纪70年代刀鲚捕捞量达到峰值, 其中1973年捕捞量为有记录的最高值, 约为4142×103kg。但自20世纪90年代以来, 受拦河筑坝、水域污染、岸坡硬化、采砂挖沙及过度捕捞等多重因素的影响, 刀鲚捕捞量急剧下滑, 2001—2005年均值降至673×103kg,2011—2013年均值降至116.6×103kg, 此后刀鲚捕捞量维持在极低的水平且波动下降, 长江刀鲚资源在40年来几乎走向了枯竭的边缘[19]。在此期间, 与长江刀鲚资源整体衰退趋势基本一致, 长江口刀鲚资源急剧下降, 最大降幅达99.06%[5]。本研究结果显示, 随着鱼汛后延, 2019—2020年长江口刀鲚样本平均体长和平均体质量均显著上升(P<0.001), 表明鱼汛早期阶段洄游的个体规格较小, 后期规格增大。长江口水域尚未全面禁捕之前, 在调查期间样本规格呈现出逐渐增大的趋势, 与历史研究报告的大规格刀鲚通常先行入江洄游的趋势并不一致[20],这应有两个方面的原因, 一是较早洄游的大规格刀鲚会率先被捕捞, 加之其在种群中占比较低, 因此越向长江口内, 捕获到大规格刀鲚的概率越低; 二是早期刀鲚经济价值较高, 随着时间后延, 特别是4月中旬以后, 刀鲚价格大幅下降, 捕捞意愿下降,捕捞强度减弱[21—23]。2021年调查结果则与之不同,3月和4月刀鲚群体平均体长和平均体质量显著大于5和6月, 表明大规格个体较早洄游进入长江下游产卵。这应是大规格刀鲚积累了更多的能量储备,在没有生产性捕捞扰动的情况下, 可以顺利进入长江口进行生殖洄游所致。
长江口是刀鲚进入长江进行上溯洄游的起点,该水域的水温、水深和盐度等环境因子均可能影响其洄游过程。在生殖洄游期间, 刀鲚对能量的需求较高, 而水温的高低将直接影响其生理和代谢活动, 所以刀鲚的洄游受温度的影响较大[24]。就性腺发育来看, 2019—2021年第1和第2时段水温在13.59—16.7℃, 刀鲚性腺发育期以Ⅰ—Ⅲ期为主,占比大于97.34%; 随水温的上升(21.8—27.18℃),雌鱼卵巢发育期在Ⅳ—Ⅴ期的个体比例显著增加,可见水温的升高加速了性腺发育[25,26]。在洄游早期, 性腺未成熟个体可能上溯至长江安庆段或鄱阳湖产卵, 而后期性腺发育成熟的刀鲚会沿江寻找适宜水域进行繁殖, 可能在长江河口段或偏上游的下游河段产卵[27,28]。上述结果表明, 长江刀鲚在较长时段内梯次经长江口洄游入江, 且性腺发育随着时间后延渐趋成熟, 长江刀鲚产卵场并不集中, 应主要分散分布于长江下游至鄱阳湖水域, 当性腺发育成熟, 在水环境条件适合的水域都可以完成繁殖活动[29]。
3.2 长江口刀鲚洄游群体的种群结构
渔获规格通常是生物学最小型的直接表现, 在一定程度上反映了种群结构变化。从整体上来看,2019—2021年长江口刀鲚体长均值较10年前增加了12.62%, 体质量均值增加了63.39%, 体长240—300 mm的比例由2017—2018年的52.05%上升至59.85%, 小规格刀鲚的比例下降, 中、大规格刀鲚的比例增加[30—32]; 平均丰满度较2017—2018年增加。本研究结果表明自2019年以来, 长江口大规格刀鲚比例增加, 且体长和体质量显著大于2017—2018年, 资源恢复效果良好。近3年刀鲚体重均值逐年增大, 且显著大于生产性捕捞退出前(2017—2018年)。尽管2019年刀鲚生产捕捞即已退出, 但由于受前期过度捕捞的影响, 加之长江口其他渔业生产尚未全部退出, 因此2019年刀鲚规格并未立即出现恢复趋势。经过3年的资源养护, 加之自2021年1月1日起, 长江流域重点水域禁捕及长江口水域全面禁捕, 极大减轻了捕捞压力, 长江口刀鲚规格即出现极显著增大。
3.3 长江口刀鲚洄游群体的资源密度
与之前的研究结果有所差异, 长江口水温变化和资源密度不存在显著的相关关系, 资源密度并未随水温的回升而明显增加[4,7]。与刀鲚样本规格的变化趋势类似, 刀鲚资源密度同样表现出时间特征。2019年整体上随洄游时间的后移呈上升趋势,而2020年和2021年均表现为前高后低的趋势, 这同样反映了自2019年刀鲚生产性捕捞退出后, 其捕捞压力逐渐消除及资源逐渐恢复的过程。刀鲚资源密度在空间上也呈现一定规律, 表现为在深水区域采集的样本尾数、重量、NB、WB、Nt和Wt显著大于浅水区域, 表明刀鲚在水深大于10 m的水域中分布相对集中, 与之前的研究结果一致[33,34]。
3.4 禁捕管理对长江口刀鲚资源的影响
长江口是洄游性物种江海洄游的重要通道, 因此长江口对洄游性物种生活史的整体保护至关重要[35]。2019年汛期内刀鲚调查总重量和资源密度均低于生产性捕捞退出前, 推测其原因可能有三个方面, 一是尽管刀鲚专项捕捞退出, 但长江口其他渔业生产依然存在, 捕捞的影响并未消除; 二是在长江流域重点水域全面禁捕之前, 长江口渔政监管难度极大, 非法捕捞现象并未杜绝; 三是资源恢复需要一定的时间过程, 存在一定的滞后性。自2021年1月1日起, 长江流域重点水域全面禁捕的实施和长江口禁捕管理区的成立, 从根本上消除了禁捕范围内的捕捞压力, 打通了刀鲚的洄游通道, 长江口刀鲚的资源量将会大幅上升。本研究的调查结果证实了这一资源的恢复, 2021年长江口刀鲚调查期内资源密度显著大于2019年和2020年, 逐年成倍增加, 这与鄱阳湖水域刀鲚数量和资源量出现极显著增加的趋势基本一致[36,37]。随着长江十年禁渔的全面实施, 可以从根本上缓解渔业资源的捕捞压力, 长江流域包括刀鲚在内的鱼类资源将逐步得以恢复。但考虑到其他人类活动的干扰仍将长期存在, 应针对刀鲚等洄游性物种资源开展长期跟踪监测, 以系统评估资源恢复效果, 支撑渔业主管部门有关行政决策。
