75 份箭筈豌豆种子形态特征遗传多样性评价
2022-10-21陆忠杰杨正禹
陆忠杰,杨正禹,罗 奔,李 舟,董 瑞
(1. 凉州区农业技术推广中心,甘肃 武威 733000;2. 贵州大学动物科学学院, 贵州 贵阳 550025)
箭筈豌豆(Vicia sativa)是野豌豆属中重要的一年生、二倍体、自花授粉豆科植物[1],具有生育周期短、适应性强、兼固氮及改善土壤结构的能力,是一种重要的饲草和绿肥兼用类作物。在中国、土耳其、新西兰等国广泛种植[2-4],在我国普遍种植于华北、西北以及长江中下游等高海拔地区[5]。
野豌豆属植物全球约有190 多个物种[6],其中在欧洲、高加索地区以及中国分布的种类最多,是该属植物的3 个现代分布中心[7]。Ball[8]首次将野豌豆属植物划分为Vicia、Cracca、Ervum和Faba4 个亚属。Maxted[9]在表型分类的基础上又将亚属细分为9 大系列,38 个种,14 个亚种。我国有野豌豆属植物43 种,其中栽培种类5 种,野生种类36 种(其中两个被合并)[10]。
种质资源是作物育种的重要物质基础,在开发新的优良作物栽培品种方面发挥着重要的作用[11]。箭筈豌豆遗传多样性可为该种的改良以及选育提供遗传基础。对大量种质多种性状进行比较筛选,发现特异性高且可用性强的遗传变异性状,是种质改良的常用手段。王鹏等[12]对30 份不同来源的大豆(Glycine max)种质的17 个主要农艺性状进行分析,筛选出了‘晋豆39’和‘中作966’两种农艺性状表现突出的种质。梁国玲等[13]对590 份燕麦(Avena sativa)种质的穗部进行遗传多样性分析,筛选出了117 份种质作为粮饲兼用型育种目标亲本。陈柳宏等[14]对东北地区的205 份甜菜(Beta vulgaris)的5 个品质性状、18 个农艺性状以及3 个抗性性状分别进行遗传多样性分析,筛选出 5 份高抗褐斑病兼抗根腐病种质,可作为抗病育种的优质亲本进行改良选育。
种子的形态多样性变异是植物遗传变异的重要特征之一,在遗传分类学上具有重要研究价值[15],可作为种内及种间的分类依据,以及用来鉴定某一特异性品种[16]。种子形态性状还可作为作物改良计划中的选择标准[17-18],还是识别种子的重要依据,其与种子的繁衍能力、种子的萌发、幼苗建植和存活,甚至种群的分布格局密切相关[19]。且牧草种子的形态特征是对其进行分级和检验的重要手段,是研究各种牧草种子真实性以及纯度鉴定的主要依据[20]。种子的形态特征与其他植物器官相比,以种子作为试验材料来研究物种的遗传多样性具有操作方便、易收集且易储存等众多优点[21]。周生坛等[22]对251 份鹰嘴豆(Cicer arietinum)的11 个种子表型性状数据进行综合分析,为鹰嘴豆优异基因发掘、育种改良以及种质创新提供了理论依据。郭宇琦等[23]对104 份北方春大豆地方品种的10 个种子活力相关的性状进行测定,筛选出了‘保险豆’、‘汤原秃荚子’等8 个高活力地方品种,可作为培育高活力品种的亲本材料。
然而,我国对箭筈豌豆种子形态性状的遗传变异研究较少。这对于选育具有较强环境适应性、较高抗逆性且种植范围较广的箭筈豌豆品种造成了极大的不便。因此本研究选用广泛分布于世界各地的12 份V.sativa和63 份V.sativasubsp.nigra亚种,共75 份箭筈豌豆种质作为试验材料。通过对各个种质的9 个种子性状进行分析,并估计关键性状的基因型变异,旨在筛选出具有优良的种子形态特征的种质,为箭筈豌豆的新品种的选育以及种质鉴定提供有价值的数据和材料基础。
1 材料与方法
1.1 种质来源
75 份箭筈豌豆种质分别为12 份箭筈豌豆和63份箭筈豌豆亚种。75 份种质种子分别由美国种质资源库(National Plant Germplasm System of the United States, NPGS)和兰州大学草地农业科技学院提供,将兰州大学选育的‘兰箭1 号’、‘兰箭2 号’、‘兰箭3 号’箭筈豌豆本地栽培品种作为对照。种质详细信息如表1 所列。

表1 75 份箭筈豌豆种质信息Table 1 Information of 75 common vetch accessions
1.2 试验地概况
试验地点位于贵州省贵阳市花溪区贵州大学实验田(106°07′ E,26°11′ N,海拔1 100 m),地处贵州高原中部。该区具有明显的高原气候特点,属于亚热带湿润温和型气候。年平均气温14.8 ℃,年均降水量1 347.3 mm。
1.3 试验设计
试验共进行8 个月,从2020 年10 月到2021 年5 月。每个种质每个重复种植10 个单株,3 个重复,每个种质的一个重复种植在同一个花盆内。用贵州本地普通黄壤土过筛(2 mm)后作为种植土壤,每盆装土约5 kg,花盆大小为35 cm (高) × 22 cm (底径)。
1.4 测定项目及其方法
在2021 年3 月至5 月种植期间内,根据不同种
质种子的成熟时间,对种子进行分批次收集。将收集到的种子进行烘干,使其含水量保持在9%~10%,然后在4 ℃ 条件下进行储存,于每份种质每个重复中随机挑选50 粒种子作为待测样品。
本研究中共测定了9 个种子性状,分别为种子长(SL)、宽(SW)、周长(Pe)、厚度(ST)、弯曲长度(CL)、弯曲宽度(CW)、种脐长(HL)、百粒重(WS)和形状。种子形状示意图如图1 A 所示。
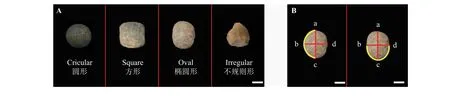
图1 种子形状和种子长、宽等性状测量位置示意图Figure 1 Schematic diagram of the measurement position of seed shape, seed length, and width
对种子长、宽、周长、厚度、弯曲长度、弯曲宽度5 项指标进行测量时,首次使用平板扫描仪(Epson Perfection V800 Photo)对每份种质每个重复的50 粒待测种子样品进行扫描,扫描图片的规格为350 dpi的图像分辨率以及2 040 × 2 040 Px 的像素。在扫描期间为防止种子的大小、形状以及重量等性状发生变化,实验室的温度和相对湿度应保持在20 ℃和15%。最后通过WinRHIZO 图像分析系统(加拿大)对扫描图像进行处理,并得到相应数据。种子厚和种脐长度用游标卡尺进行测量,百粒重则使用万分之一天平进行称量,详细测量方法见文献[24]。
1.5 数据处理
数据分析基于:1)进行方差分析以评估各种质间基因型变异的显著性和大小。2)模式分析,包括聚类分析和主成分分析(principal component analysis,PCA)[25-27],以图形方式对按性状收集的数据矩阵进行了总结,其中聚类分析通过K-means 聚类法(stats 包)完成。
使用GenStat 7.1 (2003)中的“残差最大似然”(residual maximum likelihood,REML)选项进行方差分量分析。随机线性模型用于分析亚种中不同种质的数据。详细数据分析方法见Dong 等[28]的文章。
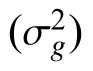
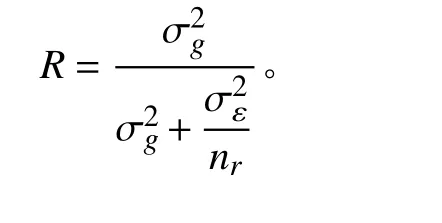
2 结果与分析
12 份箭筈豌豆和63 份箭筈豌豆亚种种质之间除形状外的8 个种子性状均存在显著的基因型变异(P< 0.05) (表2)。根据平均重复值(R)可判断各表型性状的遗传稳定性。12 份箭筈豌豆和63 份箭筈豌豆亚种的种子长、宽、周长、弯曲长度以及弯曲宽度具有较为相似的R估计值,均大于0.964。且性状弯曲宽度在箭筈豌豆和箭筈豌豆亚种中具有最高的R估计值,分别为0.981 和0.965,而性状种脐长则在箭筈豌豆和箭筈豌豆亚种中具有最低的R估计值,分别为0.972 和0.798 (表2)。对8 个种子性状均值分析发现,箭筈豌豆和箭筈豌豆亚种之间,8 个性状均存在显著差异(P< 0.05) (表2)。
表2 箭筈豌豆种质内估计基因型( )、试验误差()方差分量及相关标准误差( ± SE)评估Table 2 Estimated genotype ( ) , experimental error (), variance component, and related standard error ( ± SE)evaluation from 12 Vicia sativa and 63 V. sativa subsp. nigra accessions
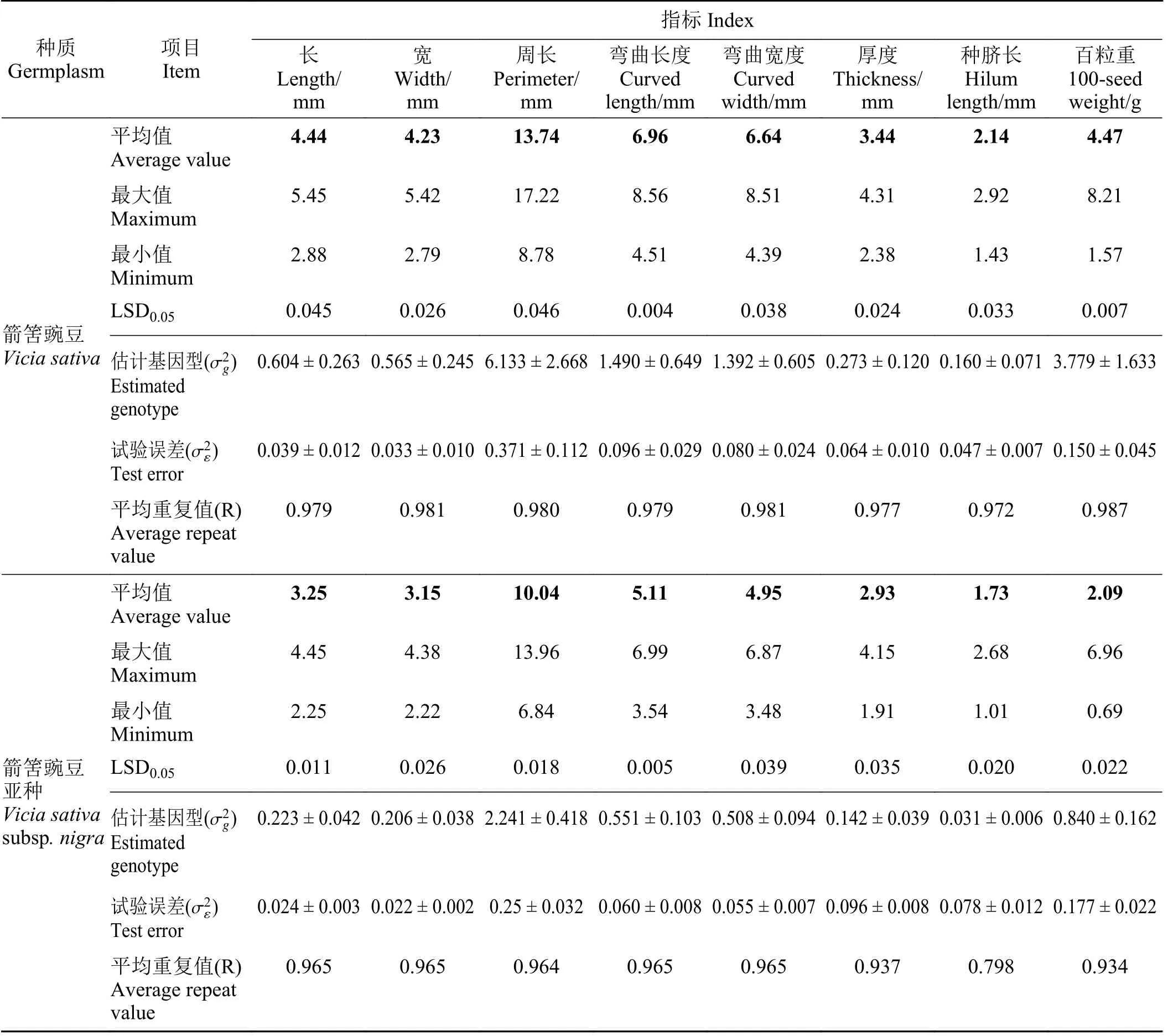
表2 箭筈豌豆种质内估计基因型( )、试验误差()方差分量及相关标准误差( ± SE)评估Table 2 Estimated genotype ( ) , experimental error (), variance component, and related standard error ( ± SE)evaluation from 12 Vicia sativa and 63 V. sativa subsp. nigra accessions
加粗的平均值代表箭筈豌豆和箭筈豌豆亚种间存在显著差异(P < 0.05)。Means in bold represent significant differences between the common vetch and common vetch subspecies (P < 0.05).
指标 Index种质Germplasm项目Item 长Length/mm宽Width/mm周长Perimeter/mm弯曲长度Curved length/mm弯曲宽度Curved width/mm厚度Thickness/mm种脐长Hilum length/mm百粒重100-seed weight/g平均值Average value最大值Maximum 4.44 4.23 13.74 6.96 6.64 3.44 2.14 4.47 5.45 5.42 17.22 8.56 8.51 4.31 2.92 8.21最小值Minimum 2.88 2.79 8.78 4.51 4.39 2.38 1.43 1.57 LSD0.05 0.045 0.026 0.046 0.004 0.038 0.024 0.033 0.007箭筈豌豆Vicia sativa σ2g估计基因型( )Estimated genotype 0.604 ± 0.263 0.565 ± 0.245 6.133 ± 2.668 1.490 ± 0.649 1.392 ± 0.605 0.273 ± 0.120 0.160 ± 0.071 3.779 ± 1.633 σ2ε试验误差( )Test error 0.039 ± 0.012 0.033 ± 0.010 0.371 ± 0.112 0.096 ± 0.029 0.080 ± 0.024 0.064 ± 0.010 0.047 ± 0.007 0.150 ± 0.045平均重复值(R)Average repeat value 0.979 0.981 0.980 0.979 0.981 0.977 0.972 0.987平均值Average value最大值Maximum 3.25 3.15 10.04 5.11 4.95 2.93 1.73 2.09 4.45 4.38 13.96 6.99 6.87 4.15 2.68 6.96最小值Minimum箭筈豌豆亚种Vicia sativa subsp. nigra 2.25 2.22 6.84 3.54 3.48 1.91 1.01 0.69 LSD0.05 0.011 0.026 0.018 0.005 0.039 0.035 0.020 0.022 σ2g估计基因型( )Estimated genotype 0.223 ± 0.042 0.206 ± 0.038 2.241 ± 0.418 0.551 ± 0.103 0.508 ± 0.094 0.142 ± 0.039 0.031 ± 0.006 0.840 ± 0.162 σ2ε试验误差( )Test error平均重复值(R)Average repeat value 0.024 ± 0.003 0.022 ± 0.002 0.25 ± 0.032 0.060 ± 0.008 0.055 ± 0.007 0.096 ± 0.008 0.078 ± 0.012 0.177 ± 0.022 0.965 0.965 0.964 0.965 0.965 0.937 0.798 0.934
通过对种子形状的分析发现,种子形状在12 份箭筈豌豆和63 份箭筈豌豆亚种中的变异相对较低。除了在63 份箭筈豌豆亚种中有20 份椭圆形种子的种质以及方形、不规则形种子种质各1 份外,其余种质种子均为圆形种子。
2.1 12 份箭筈豌豆种质种子形态特征模式分析及表型相关性
主成分分析和聚类分析结合的模式分析主要通过图形化汇总形式分析和比较75 份箭筈豌豆各种质种子性状之间的差异,同时还能直观地表现出性状之间的关联信息(正相关性或负相关性)[30]。其中,各性状之间的相关性由方向向量表示,不同符号则表示聚类分析产生的不同分组。对12 份箭筈豌豆种质进行模式分析,第1 主成分解释了总体性状变异的94.82%,第2 主成分解释了2.8%(图2)。在12 份箭筈豌豆亚种种质中,8 个种子性状两两性状之间呈现极显著(P< 0.01)正相关关系(方向向量之间的角度 < 90°) (图2)。
在模式分析中,根据8 个种子性状将12 份箭筈豌豆种质进行聚类分析,可将其分为两个群组(图2),其中第Ⅰ组群种质的种子长、宽、周长、弯曲长度、弯曲宽度、厚度、种脐长以及百粒重8 个性状的平均值均高于第Ⅱ组群种质的平均值(表3)。其中,兰箭1 号、2 号、3 号和67 号4 个种质为第Ⅰ组群种质,8 个种质为第Ⅱ组群。
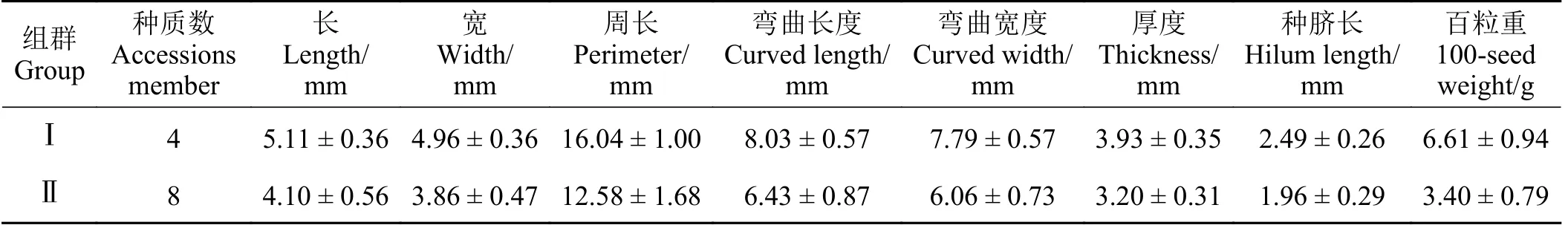
表3 12 份箭筈豌豆种质聚类分析产生的两个种群各定量性状均值Table 3 The mean values of each quantitative trait of 2 groups from the cluster analysis of 12 Vicia sativa accessions
表4 中所示的表型相关系数(rp)进一步验证了图2 中所示的性状种子长、宽、周长、弯曲长度、弯曲宽度、厚度、种脐长以及百粒重8 个性状之间具有极显著正相关性。其中,最小值为厚度和种子长以及厚度和弯曲长度之间的0.854,最高值则为种子长和弯曲长度之间的0.999。
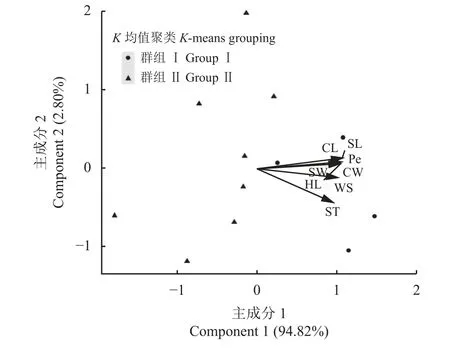
图2 12 份箭筈豌豆种质8 个种子性状的模式分析Figure 2 Model analysis of 8 seed traits of 12 Vicia sativa accessions
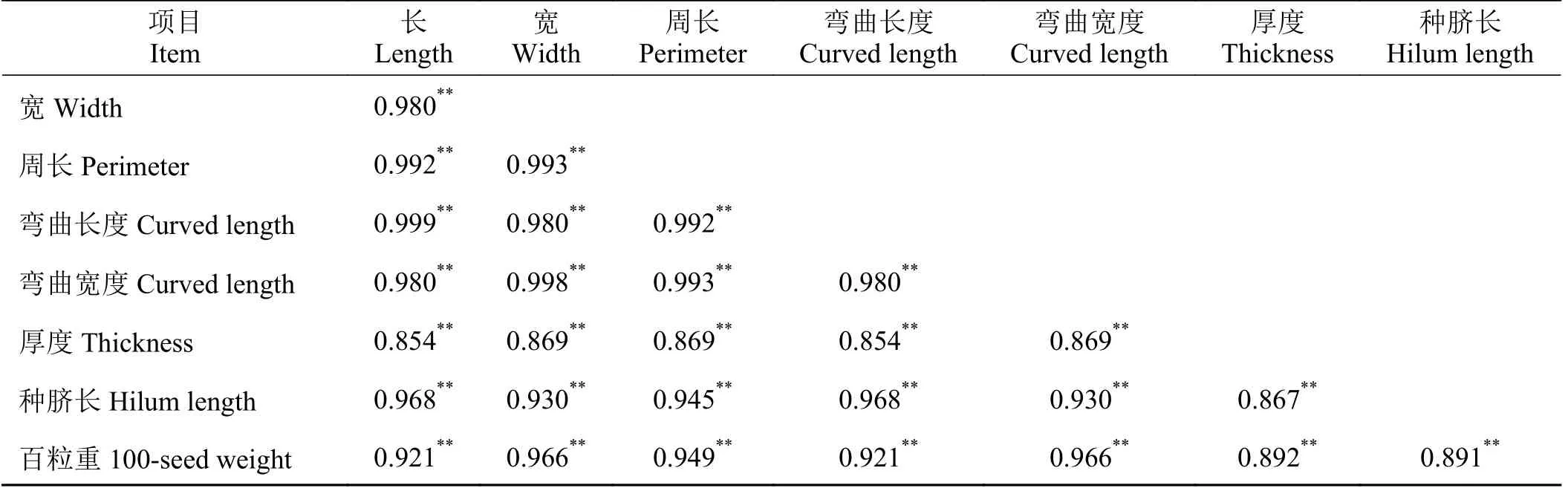
表4 12 份箭筈豌豆种质各性状之间的表型(rp)相关系数Table 4 Phenotypic (rp) correlation coefficients between the traits of 12 Vicia sativa accessions
2.2 63 份箭筈豌豆亚种种子形态特征模式分析及表型相关性
63 份箭筈豌豆亚种种质基于种子长、宽、周长、弯曲长度、弯曲宽度、厚度、种脐长以及百粒重8 个性状的模式分析结果表明,第1 主成分解释了总体性状变异的84.0%,第2 主成分解释了9.1%(图3)。在63 份箭筈豌豆亚种种质中,8 个种子性状两两之间呈现极显著(P< 0.01)正相关关系(图3)。
同时,基于种子长、宽、周长、弯曲长度、弯曲宽度、厚度、种脐长以及百粒重 8 个性状对63 份箭筈豌豆亚种种质进行聚类分析,可将其分为3 类(表5)。其中第Ⅰ组群包含的种质最多,达33 份种质;第Ⅱ组群次之,有16 份种质;第Ⅲ组群包含的种质最少,只有14 份种质(表5)。同时,第Ⅱ组群中8 个性状的平均值均高于其余组群的平均值。而在第Ⅲ组群中,8 个性状的平均值均低于其余群组的平均值(表5)。
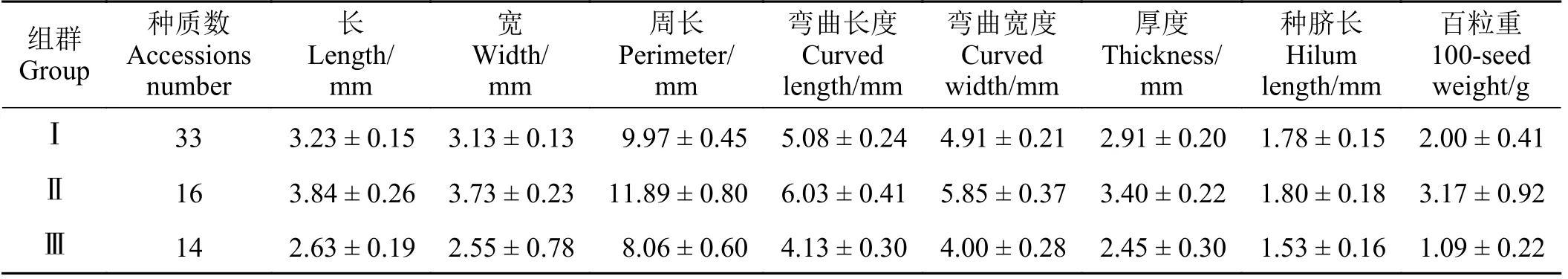
表5 63 份箭筈豌豆亚种种质聚类分析产生的3 个种群各定量性状均值Table 5 The mean values of quantitative traits of 3 groups generated by cluster analysis of 63 Vicia sativa subsp. nigra accessions
表型相关系数(rp)进一步验证了图3 中所示的性状种子长、宽、周长、弯曲长度、弯曲宽度、厚度、种脐长以及百粒重之间的相关性(表6)。性状种子长、宽、周长、弯曲长度、弯曲宽度、厚度以及百粒重的所有成对组合的rp值均极显著(P< 0.01)正相关,其中,最小值为种子长和百粒重之间的相关系数0.759,最高值则为种子宽和弯曲宽度之间的相关系数为0.999。种脐长和其余7 种性状之间的rp值则略低,但也存在极显著的正相关关系(表6)。
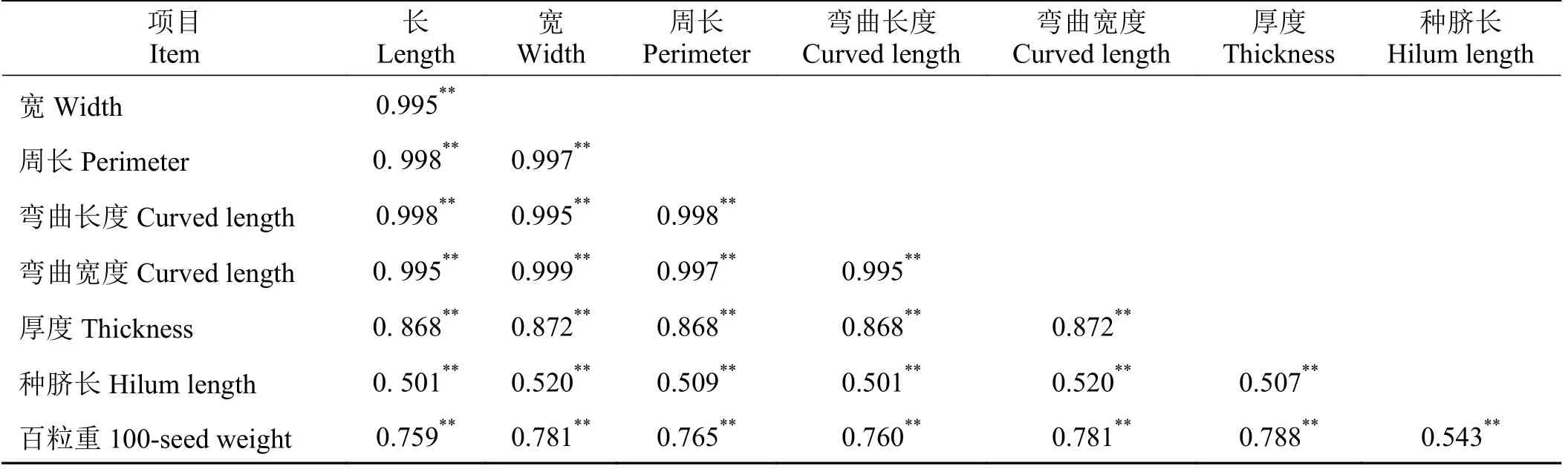
表6 63 份箭筈豌豆亚种种质各性状之间的表型(rp)相关系数Table 6 Phenotypic (rp) correlation coefficients among the traits of 63 Vicia sativa subsp. nigra accessions
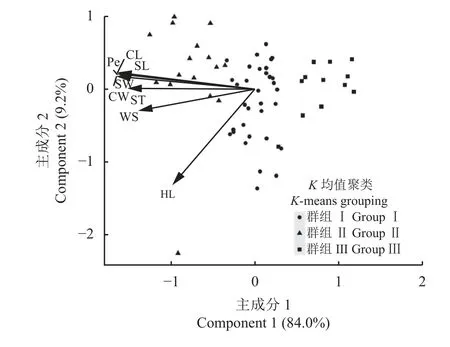
图3 63 份箭筈豌豆亚种种质8 个种子性状的模式分析Figure 3 Model analysis of 8 seed traits of 63 Vicia sativa subsp. nigra accessions
3 讨论
在本研究中,12 份箭筈豌豆以及63 份箭筈豌豆亚种之间,8 个种子性状种子长、宽、周长、弯曲长度、弯曲宽度、厚度、种脐长以及百粒重均存在显著(P< 0.05)的基因型变异。这些种子性状的基因型变异和平均重复性(R)估计值表明,在所试的75 份箭筈豌豆种质内均有潜在可用的遗传变异。种子长、种子宽以及百粒重的变化范围在12 份箭筈豌豆以及63 份箭筈豌豆亚种中分别为2.88~5.45 和2.25~4.45 mm、2.79~5.42 和2.22~4.38 mm、1.57~8.21 和0.69~6.96 g。同时本研究发现,所试的12 份箭筈豌豆各种质8 个性状的平均值都大于63 份箭筈豌豆亚种的(表2)。且对12 个箭筈豌豆种质聚类发现,67 号这一种质与兰箭系列的3 个种质被分在8 个性状均较高的一组(图2)。因此,本研究认为67 号这一种质的种子形态多样性对于种质的遗传改良具有重要意义。
这些与种子大小相关的性状可作为种质种植前期的筛选标准,这对于幼苗的建植和存活是十分重要的。Moles[31]的研究发现,种子的大小在种质的后期生长发育过程中各有特点。与大种子相比,小种子种质在受到同样的外界能量下可以产生更多的种子,可用于种子生产。而大种子种质应对环境变化而产生的胁迫能力更强。因此,研究认为在生长环境一定时,不同大小的种子对其种质幼苗的建植、存活以及生长发育具有显著影响。李振声[32]研究发现种子粒重是决定作物产量的重要因素,其受环境条件的影响相对较小,对种子产量的直接贡献最大,通过对种子百粒重的选择可以提高产量。因此本研究对种子种质资源进行评价,得到关键种子遗传性状的数据是选育新品种的重要依据,尤其是对种子生产以及幼苗的建植、存活以及生长发育的改良具有十分重要的意义。在本研究中,12 份箭筈豌豆以及63 份箭筈豌豆亚种各种质间的种子长、宽以及百粒重都存在着显著的差异(P< 0.05),其中在12 份箭筈豌豆内种子长和百粒重最大的种质都是兰箭1 号(5.45 mm, 8.21 g),种子宽最大的种质则是兰箭2 号(5.42 mm),而种子长、宽以及百粒重3个性状最小的种质则都是64 号(2.88 mm, 2.79 mm,1.57 g)。在63 份箭筈豌豆亚种内种子长、宽以及百粒重最大的种质分别为14 号(4.45 mm)、12 号(4.38 mm)和20 号(6.96 g)种质,而种子长、宽以及百粒重3 个性状最小的种质则都是57 号(2.25 mm, 2.22 mm,0.69 g) (表2)。因此,本研究中,64 号、57 号、20 号等种质可为箭筈豌豆种子大小改良提供育种材料和一定数据基础。目前,箭筈豌豆主要种植于我国青藏高原等高海拔地区,是该地区的主要牧草和绿肥作物。但是,这些地区往往环境恶劣,生长季较短。因此,该地区种植大种子箭筈豌豆种质有利于其幼苗的建植和存活。
根据植物的性状指标对植物进行聚类分析,在种质鉴定、筛选以及探究其亲缘关系等方面是常用且有效的方法。韩梅等[33]根据9 个产量性状对124份箭筈豌豆进行聚类分析,可将其聚为5 类。白金顺等[34]根据11 项抗旱指标对14 份箭筈豌豆进行聚类分析,将其聚为4 类。在该研究中,12 份箭筈豌豆以及63 份箭筈豌豆亚种的8 个种子性状均具有显著(P< 0.05)的基因型变异估计值,而两个亚种之间单个性状平均值的比较也呈现显著(P< 0.05)的差异(表2)。因此,本研究根据上述8 个种子性状分别将12 份箭筈豌豆种质和63 份箭筈豌豆亚种种质分为2 组和3 组(图2、图3),分类清晰且明确。综上所述,本研究认为在箭筈豌豆和箭筈豌豆之间具有显著(P< 0.05)差异的性状可用于箭筈豌豆分类学关系的研究。
目前,已有许多学者利用模式分析来识别育种研究中的重要植物材料。例如Jahufer 等[35]、Zhang 等[36]和Harch 等[37]分别在白三叶草(Trifolium repens)、 春小麦(Triticum aestivum)和花生(Arachis hypogaea)中均利用模式分析对育种材料进行了分析和研究。本研究对12 份箭筈豌豆种质以及63 份箭筈豌豆亚种种质进行模式分析,探究了其中在种子性状方面有价值和潜力的种质群体,而这些种质群体内的种质资源所蕴含的遗传多样性,对种子形态特征相关的分类学和育种研究具有十分重要的意义。
4 结论
本研究对75 份箭筈豌豆种质的种子性状进行以主成分和聚类分析相结合的模式分析、基因型变异分析和相关性分析,结果发现,在75 份种质除形状外的其他8 个种子性状均表现出两两呈极显著正相关关系(P< 0.01),且8 个种子性状均呈显著的(P< 0.05)基因型变异估计值。同时,通过对亚种内各种质单个性状平均值的比较也发现其具有显著差异(P< 0.05)。且根据8 个种子性状能清晰且明确的分别将12 份箭筈豌豆以及63 份箭筈豌豆亚种种质分别分为2 组和3 组。因此,本研究认为以上8 个种子性状均可以作为V. sativa和V. sativasubsp.nigra亚种种质之间进行分类的指标,且具有优良性状的种质也可以作为育种和品种改良的基础材料。
