不同退化程度高寒草甸生态系统碳通量
2022-10-21徐干君柴永煜吴胜义康恩泽汪晓东康晓明张骁栋
徐干君,柴永煜,颜 亮,吴胜义,李 勇,康恩泽,汪晓东,杨 澳,康晓明,张骁栋
(1. 国家林业和草原局西北调查规划院, 陕西 西安 710048;2. 湿地生态功能与恢复北京市重点实验室 /中国林业科学研究院生态保护与修复研究所, 北京 100091;3. 四川若尔盖高寒湿地生态系统定位观测研究站, 四川 阿坝 624500)
联合国政府间气候变化专门委员会(Intergovernmental Panel on Climate Change, IPCC)第六次评估报告指出,人类活动造成温室气体排放急剧增加,全球地表温度持续上升,进一步改变了陆地生态系统的碳水循环格局[1]。青藏高原因其独特的海拔高度和地理环境,对全球气候变化的响应敏感而迅速[2]。高寒草甸作为青藏高原植被覆盖的主要类型,广泛分布于青藏高原腹地以及周围的山地区域,占青藏高原总面积的35%[3]。全球变暖和人类活动引起青藏高原高寒草甸退化面积增加,水源涵养功能衰退,生态系统生产力降低,生物多样性下降,削弱了生态系统多功能性[4]。
退化的高寒草甸生态系统植物群落组成、土壤理化特征、土壤微生物群落结构均发生改变。随着高寒草甸生态系统退化加剧,植物群落由禾本科和莎草科植物逐渐被杂草取代,地上和地下生物量均随着退化程度的加剧而显著下降[5-7]。同时,退化的高寒草甸土壤孔隙度降低[8],土壤有机碳、全碳、全氮和全磷降低[6-7],葡萄糖苷酶、脲酶、蔗糖酶等土壤酶活性显著降低[9-11]。不同地区的高寒草甸退化对土壤CO2排放的影响不同,例如海北地区的土壤CO2排放量随着退化加剧而减弱[12],而风火山地区的土壤CO2排放量随着退化加剧而升高[13]。地区间土壤CO2排放变化趋势不同是由于地域气候条件和生态系统各自特征差异引起的。目前普遍认为温度和土壤水分是驱动土壤碳排放的主要因素,而土壤质地、植被组成和物候差异,都会引起土壤呼吸的敏感性有较大差异[14]。青藏高原的气候为雨热同期,夏季植物生长旺盛,光合有效辐射和叶面积指数主导了生态系统碳交换,多数研究表明夏季高寒草甸为明显的碳汇[15-16]。随着高寒草甸退化,生态系统生产力可能因植被组成和盖度的变化而削弱,使夏季的碳汇减弱或转为碳源[14]。然而,有关不同退化程度高寒草甸生态系统碳通量组分的研究仍不充分,植被生产力和土壤呼吸如何影响高寒草甸的碳源/汇变化还不清楚。
基于此,本研究提出“在高寒草甸退化过程中,不同碳通量组分对环境变化的响应存在差异,共同影响生态系统碳源/汇特征”的假设,以青藏高原高寒草甸为研究对象,选取围封到重度退化4 个退化梯度,测定不同退化梯度下生态系统碳通量、植被和环境因子等指标,探讨不同退化梯度下高寒草甸生态系统碳通量变化规律,以及环境因子对不同退化梯度高寒草甸生态系统碳通量组分的影响。
1 材料与方法
1.1 研究地概况
本研究野外调查于2021 年7 月末在青海省果洛藏族自治州东北部的玛沁县(98°48′~100°55′ E,33°43′~35°16′ N)开展。玛沁县是典型的高原山地,平均海拔4 100 m 以上,属于高原寒冷气候。2021 年玛沁县年均气温1.52 ℃,最冷月1 月平均气温-9.3 ℃,最热月7 月平均气温11.6 ℃。年均降水量628 mm,全年75%降水集中于6 月-9 月(图1)。全县总面积134 万 hm2,其中草地面积117.57 万hm2,可利用草地面积108.53 万hm2,占草地面积的92.3%[17]。玛沁县的主要植被类型为以高山嵩草(Kobresia pygmaea)、矮生蒿草(Kobresia humilis)为优势种的高寒草甸和沼泽化草甸,以及以金露梅(Potentilla fruiticosa)为优势种的高寒灌丛[18]。主要土壤类型为高山草甸土、高山灌丛草甸土和沼泽土[19-20]。玛沁县属于典型牧业区,生态系统敏感和脆弱,也是黄河重要的流经区域与水源涵养地。

图1 2021 年玛沁县月降水量和月平均气温Figure 1 Monthly precipitation and mean temperature of Maqin County in 2021
1.2 样地选择与样品采集分析
在玛沁县选择3 个不同退化程度的高寒草甸样地和1 个围封3 年的样地作为对照(表1),轻度退化和中度退化样地为夏季牧场,重度退化样地于2020 年起实施退牧还草工程处于围封状态。高寒草甸的退化程度依据植被覆盖度(vegetation cover,VC)和优势植物划分[21]。轻度退化:VC≥ 70%,可食牧草相对盖度≥ 70%;中度退化: 50% ≤ VC <70%,30% ≤可食牧草相对盖度< 70%;重度退化:VC < 50%,可食牧草相对盖度< 30%。每个样地随机选择3 个0.5 m × 0.5 m 样方,调查样方内VC,收割植物地上部分,分层采集土壤样品,并原位测定CO2和CH4通量,表层0-10 cm 土壤温度用土壤水分仪(TDR 300, Spectrum,美国)测定。
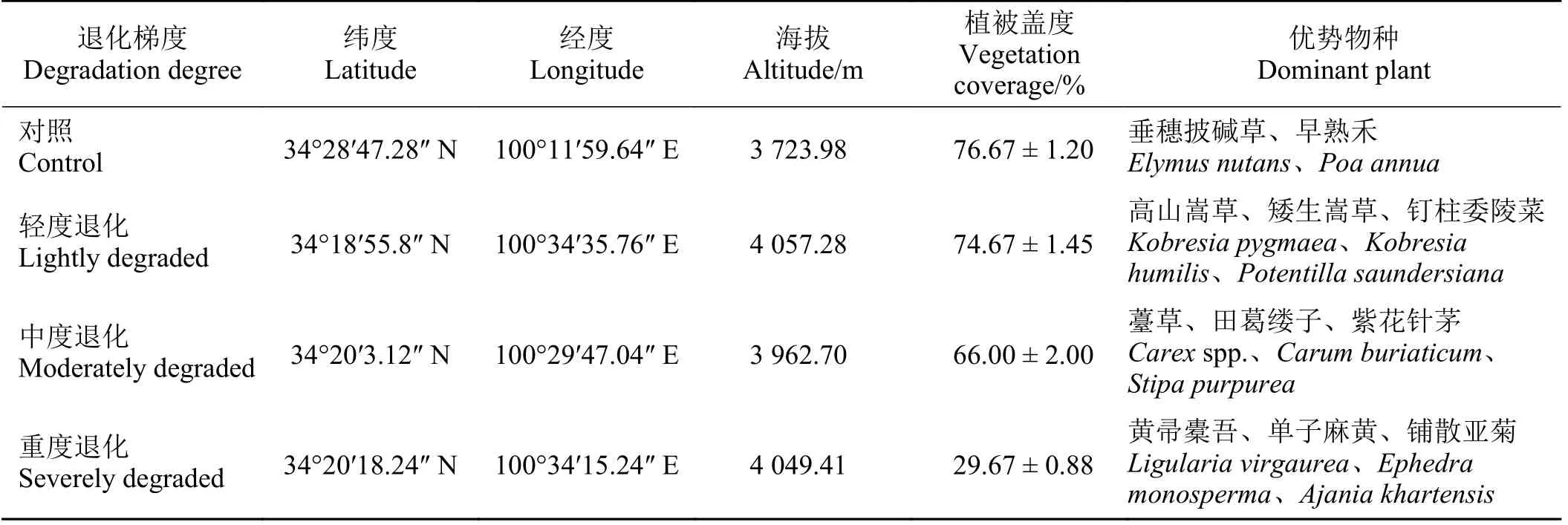
表1 样点地理位置与植被覆盖Table 1 Location and plant cover of sampling sites
植物地上部分全部收割后装入信封,在65 ℃烘箱中烘72 h 至恒质量后称重获得植物地上生物量(above ground biomass, AGB)。在 样 方 内 以 直 径3.8 cm 的土钻分层钻取0-10、10-30、30-50 cm土层的土壤样品。现场将土壤样品过2 mm 筛并去除细根,分成两份。一份土壤样品用烘干法测定土壤含水量(soil water content, SWC);另一份样品风干后用球磨仪研磨,采用元素分析仪(Vario EL III, EA GmbH, 德 国)测 定 土 壤 有 机 碳(soil organic carbon,SOC),用 1 ∶ 2.5 土水混合悬浊液测定土壤pH (PHS-3C,雷磁,中国)。
1.3 生态系统CO2 和CH4 通量测定
生态系统CO2通量组分包括生态系统净交换(net ecosystem exchange, NEE)、生态系统呼吸(ecosystem respiration, ER)和总生态系统生产力(gross ecosystem productivity, GEP)。生态系统CO2通量和CH4通量采用静态箱法测定[22],选择无云晴朗日09:00-12:00 进行。每个样地选择典型样方安装一个50 cm ×50 cm 铝合金框,方框的下侧边向下嵌入土壤3 cm。铝合金方框的上侧边有一个3 cm 宽的槽,槽中注满水后扣盖一个自制的透明同化箱(长50 cm × 宽50 cm × 高50 cm),使箱内处于封闭状态。同化箱内有两个小风扇用于混合箱内气体,箱体有一气管与温室气体分析仪(UGGA-30p, LGR, 美国)相连接。测定时同化箱扣在铝合金方框上,待系统稳定后开始计数,每10 s 记一次CO2和CH4浓度,连续记录9~12 次,根据测定时间内(90~120 s) CO2和CH4浓度的变化计算出CO2和CH4随时间的变化斜率,由此斜率结合测定期间的空气温度、水分压和气压等参数计算出单位时间内同化箱中CO2和CH4的通量。

式中:Fc为气体通量[mg·(m2·h)-1],对气体浓度进行线性回归分析求解直线斜率dc/dt,M和V0分别为气体摩尔质量(g·mol-1)和标准摩尔体积22.4 L·mol-1,P和P0分别为测量大气压与标准大气压(Pa),T与T0分别为箱内空气温度平均值和绝对温度,H为静态箱高度 (cm)。
CO2通量测定首先在光照条件下进行,此时测定的值为NEE (正值为碳排放,负值为碳吸收,以下ER 和CH4相同);然后将同化箱盖上不透光黑布测定,此时测定的值为ER;总生态系统生产力(GEP)为ER 与NEE 的差值(正值为碳吸收,负值为碳排放)。生态系统的CH4通量光照条件下与NEE同步测定。
1.4 数据分析
退化梯度对高寒草甸生态系统NEE、GEP、ER和CH4通量的效应采用单因素方差分析(one-way ANOVA)检验,然后用Duncan.test 对不同退化程度下高寒草甸NEE、GEP、ER 和CH4通量进行多重比较。NEE、GEP、ER 和CH4通量与VC、AGB、分层土壤性质的相关性采用Spearman 相关系数表示。本研究统计基于R 4.0 平台完成[23]。
2 结果
2.1 不同退化程度高寒草甸的碳通量特征
玛沁县高寒草甸7 月NEE 为-1 437.55~-98.45 mg·(m2·h)-1(CO2),表现为碳汇(图2a)。NEE 随着高寒草甸退化加剧而减弱,与对照样地相比,轻度、中度和重度退化高寒草甸生态系统NEE分别减弱了30%、70%和80%。GEP [778.8~2 481.4 mg·(m2·h)-1](CO2)和ER [342.6~1 238.3 mg·(m2·h)-1] (CO2)随着高寒草甸退化加剧呈先降低后增高的趋势。从轻度退化到中度退化样地GEP 和ER 分别降低了49%和44%,而中度和重度退化样地的GEP 和ER 均无显著差异(P> 0.05) (图2b,c)。GEP、ER 和NEE 三者间相关性显著(P< 0.05) (图3),其中GEP 与ER呈显著负相关关系,与NEE 呈显著正相关关系,NEE 与ER呈显著负相关关系。各个退化程度的高寒草甸样地均为弱小的CH4吸收汇,CH4平均吸收速率为0.029 mg·(m2·h)-1(CH4),不同退化程度样地间CH4的吸收速率无显著差异(图2d)。CH4与GEP、ER 和NEE均无显著相关性。

图2 不同退化程度高寒草甸生态系统净交换、生态系统呼吸、总生态系统生产力和甲烷通量Figure 2 Net ecosystem exchange, ecosystem respiration, gross ecosystem productivity, and methane flux in alpine meadows among different degradation degrees
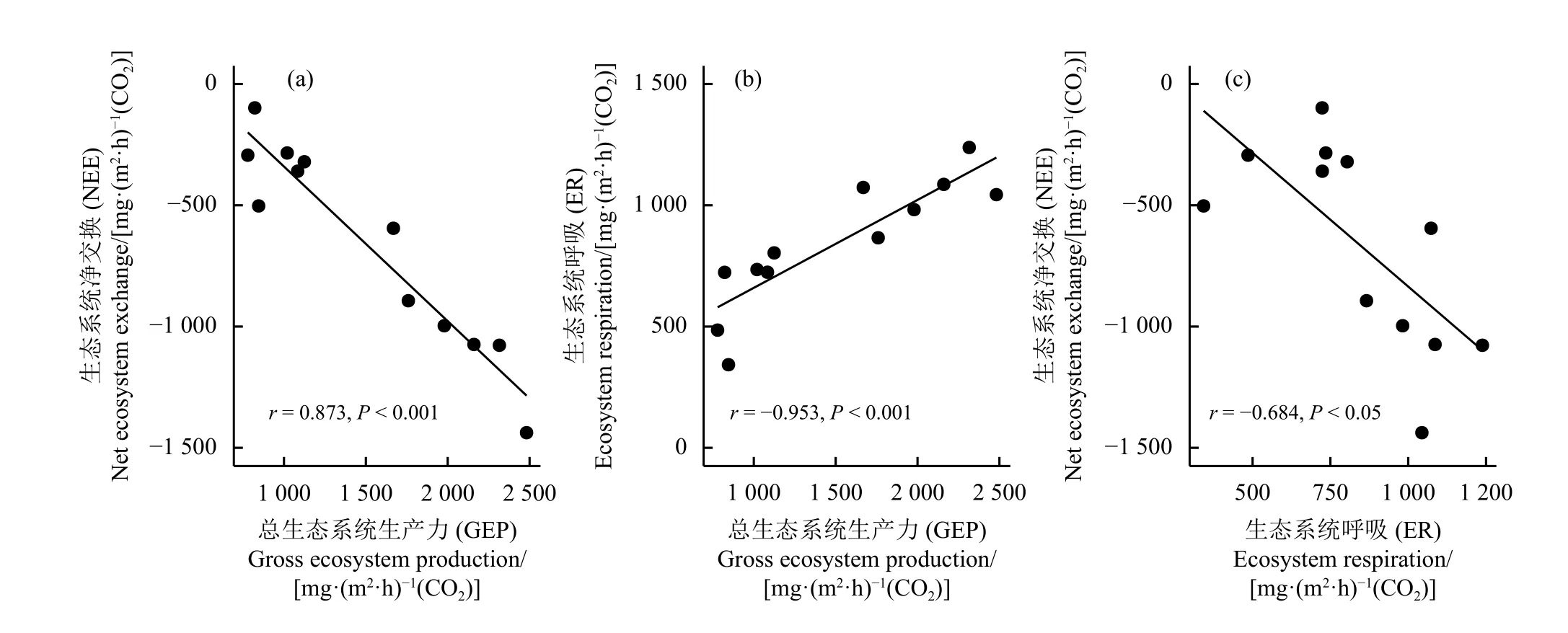
图3 二氧化碳通量组分的相关性分析Figure 3 Correlation tests among the components of CO2 fluxes
2.2 环境因子对高寒草甸碳通量的影响
不同退化程度高寒草甸的GEP、ER 和NEE 与VC 显著相关(P< 0.1),而CH4通量与AGB 显著相关(图4)。高寒草甸的0-10、10-30 、30-50 cm的SOC 含量分别为35.56、17.61、11.41 g·kg-1(表2)。0-50 cm 平均SOC 含量为轻度退化(34.58 g·kg-1) >对照(25.23 g·kg-1) > 中度退化(16.67 g·kg-1) > 重度退 化(9.64 g·kg-1)。轻 度 退 化 样 地0-50 cm 平 均SOC 含量比对照高37%,但从轻度退化到中度退化SOC 含量下降了52%。不同退化程度高寒草甸的NEE与各层SOC 含量均显著负相关,GEP、ER 仅与30-50 cm 的SOC 含量显著正相关,CH4与SOC 含量无显著相关性(图4)。GEP、ER、NEE 和CH4通量与不同深度的SWC、土壤pH 和表层土壤温度无显著相关关系(图4)。

表2 不同退化程度高寒草甸土壤和植被特征Table 2 Soil and vegetation characteristics in alpine meadows with different degradation degrees

图4 高寒草甸碳通量组分与环境因子的相关性热图Figure 4 Heatmap for the correlation between CO2 flux components and environmental factors
3 讨论
3.1 夏季高寒草甸的碳汇作用
本研究结果表明,玛沁高寒草甸在夏季表现为显著的CO2汇,同时也能吸收少量CH4(图2a,d),与大多数的研究结果一致[12,24]。与相同区域基于涡度相关法的研究比较,玛沁县极度退化的“黑土滩”测 定7 月 的NEE 为-232.73 mg·(m2·h)-1(CO2)[25],与本研究中重度退化样地[-272.69 mg (m2·h)-1] (CO2)接近。建植1 年和4 年的垂穗披碱草(Elymus nutans)栽培草地NEE 一年中最大日值发生于夏季,分别为-363.61[26]和-270 mg·(m2·h)-1(CO2)[27]。考虑到基于涡度相关法测定的NEE 包含了昼夜波动,垂穗披碱草栽培草地的夏季NEE[26-27]应高于本研究中的中度退化样地测得的昼间值[-372.30 mg·(m2·h)-1] (CO2),可能与轻度退化样地[-828.35 mg·(m2·h)-1] (CO2)相当。基于以上多个研究得出,不同退化程度的高寒草甸在夏季均表现出较强的碳汇作用。
在气候变化和人类活动的双重影响下,高寒草甸退化引起碳汇功能发生改变[28]。本研究中NEE随着草地退化加剧而减弱,且从轻度退化到中度退化阶段的降幅最大达55% (图2),同时SOC 含量下降约50%,说明SOC 可能在这一阶段大量释放,削弱了生态系统的碳汇功能[29]。另一方面,与轻度退化的样地相比,植物物种结构从莎草科为优势种转为杂草为优势种(表1),植物功能群变化使地下根系组成变化[30],旧根系分解可能也是削弱生态系统碳汇的因素之一。
3.2 退化高寒草甸不同碳通量组分变化
与对照样地相比,本研究轻度和中度退化的高寒草甸NEE 和GEP 显著相关(图3a),说明高寒草甸在退化过程中碳汇减弱主要是由于GEP 减弱引起的。王斌等[25]的研究也发现高寒草甸退化过程中生态系统全年总初级生产力的降幅大于ER,使全年NEE 从负值转为正值。高寒草甸夏季GEP 受光合有效辐射和叶面积指数共同调控[15],退化样地中较低的植被盖度使光合作用强度减弱,从而降低了GEP (图4)。本研究中未发现AGB 与GEP 的相关性(图4),与前人的研究不一致[31-32],可能是中度退化样地中的杂草繁殖体占据了较大生物量比例导致。本研究调查的中度退化样地中的田葛缕子(Carum buriaticum)为优势种之一(表1),该植物正处于盛花期,白色伞形花序对光合作用贡献甚微,使AGB 和GEP 的相关性分析呈不显著。除去中度退化样地,本研究中对照样地、轻度退化样地和重度退化样地的AGB 和GEP 呈显著正相关关系(r=0.817,P< 0.01)。
随着生态系统退化ER 呈下降趋势,中度退化样地的ER 均值最低(图2b)。ER 与GEP 及VC 的显著相关性(图3b,图4)说明植物呼吸可能在ER 的变化中占主导作用,前人研究表明高寒草甸中植物呼吸占ER 的50%~80%[33-34]。随着退化高寒草甸中VC 降低,植物呼吸也随之降低[35-36]。SOC 与ER显著正相关(图4),可能是土壤中充足的碳底物促进微生物异养呼吸[37-38],对ER 的变化有一定贡献。本研究发现调查的样地中SOC 在轻度退化的样地中最高,尤其是0-10 cm SOC 比对照样地高1 倍(表2),可能是牧场的牛粪输入引起的。轻度退化的草地SOC 升高促进微生物异养呼吸,弥补了因植被减少而降低的植物呼吸,使对照和轻度退化样地的ER 差异不显著(图2b)。然而,随着高寒草甸的进一步退化,SOC 快速流失造成了从轻度退化到中度退化样地ER 显著减弱。
本研究中发现SWC 对NEE、GEP 和ER 的作用不明显(图4),与部分已有研究结果不同[39-40]。从对照到中度退化的样地,土壤SWC 均在20%左右且无明显降低(表2)。本研究调查当月降水量仅为76 mm (图1),低于往年平均水平100 mm,而玛沁县7 月的蒸发量为130~140 mm,高于降水量近1 倍。因此,调查期间玛沁可能正经历轻度干旱。夏季降水是高寒草甸植物生长和微生物活动的主要水分来源[41-42],进而驱动了碳循环过程。本研究中可能是研究地SWC 正处于较低水平,从而对NEE、GEP和ER 的影响较弱。另外,本研究也未发现表层土壤温度与碳通量有相关性(图4),可能是由于较高的土壤温度促进土壤水分蒸发,加剧环境干旱胁迫,使其与对微生物活动的促进作用抵消[43]。
4 结论
本研究的调查发现,夏季不同退化程度的高寒草甸总体上为CO2汇,范围为-98.45~-1 437.55 mg·(m2·h)-1(CO2),随着退化加剧生态系统CO2汇能力减弱。同时高寒草甸也呈微弱的CH4汇,平均CH4吸收速率为-0.029 mg·(m2·h)-1,CH4通量随退化程度无明显趋势。植被盖度和土壤有机碳是影响高寒草甸碳汇能力的主要因素。高寒草甸从轻度向中度退化演替时系统碳汇能力下降55%,可能是这个阶段植物群落组成变化,土壤有机碳快速流失引起的。因此,在恢复治理退化高寒草甸时,需加强防止草地从轻度退化到中度退化演替,将有助于维持高寒草甸的碳汇功能。
