地下水矿化度对柽柳根系生长及构型的影响
2022-10-17彭广伟赵西梅夏江宝
彭广伟 孙 佳 赵西梅 房 颖 夏江宝
(1. 山东高速鸿林工程技术有限公司,山东 济南 250001;2. 滨州学院,山东省黄河三角洲生态环境重点实验室,山东 滨州 256603;3. 山东农业大学林学院,山东 泰安 271018)
根系构型描述了土壤中根系的形状和空间布局[1],是决定植物有效利用资源的重要因素。根系构型对基因和环境都具有很高的可塑性[2]。在单体植物的生命周期内,也可以形成不同的根系构型,因为根系各个部分发育和生长的速率可以通过短期环境信号而改变,包括土壤水分、养分和氧气的供应或病原体和害虫[2]。根系构型可以在特定的环境条件下(如干旱)提供生长优势,并对植物其他器官的生长产生直接影响。因此,根系构型研究是量化根系生长发育的重要手段。
黄河三角洲是黄河入海携带泥沙沉积而形成的冲积平原,地处河、海、陆交界处,生态系统类型独特,具有重要的生态功能[3-4]。黄河三角洲地区土壤盐渍化严重,地下水普遍埋藏较浅[5],地下水矿化度分为淡水、微咸水、咸水、盐水4种类型,其中咸水和盐水分布广泛[6-7]。浅层地下水是影响盐分迁移、积累和释放的主要因素,如地下水矿化度升高或地下水水位降低,会导致土壤含水量和含盐量显著升高[4,8-9],进而通过根系影响植被的生长发育、分布和演替[4,10-11]。在滨海湿地及部分三角洲地区,地下水水位和土壤水分、盐度的联合效应比其他区域更为显著[12-13]。目前对地下水与植物根系关系的研究主要集中在根系生长对地下水水位的响应特征[14-15]。刘深思等[14]研究了沙柳(Salix cheilophila)、柠条(Caragana korshinskii)等5种沙地建群灌木树种的生物量分配、根系分布及生理特征对地下水埋深变化的响应,发现不同树种对地下水埋深的响应存在明显差异,同时也具有一致性。日本沿海森林黑松(Pinus thunbergii)根系对地下水位具有较强的可塑性,由高水位至低水位,黑松根系由水平分布为主转变为以垂直分布为主,且根冠比降低[15]。目前,尚缺乏黄河三角洲植物根系对于地下水矿化度的响应研究,尤其是适宜植物幼苗生长关键期的地下水矿化度条件不清晰,在一定程度上导致植物幼苗栽植成活率较低。
柽柳(Tamarix chinensis)作为黄河三角洲地区的优势灌木种之一,对于提高该区域内的生态环境状况和海岸带生态稳定性具有重要作用[16]。研究表明,过高的地下水矿化度(大于盐水)严重抑制了柽柳的生长,且对根系的抑制作用大于对地上部的抑制作用;适当增加地下水盐度可显著提高柽柳的光合能力;柽柳幼苗在微咸水条件下保持了最高的光合效率、耗水能力、光能利用效率和水分利用效率[17]。目前,已对黄河三角洲地区柽柳根系特征进行了初步研究,随地下水水位的降低,柽柳根系拓扑构型由叉状分支向鱼尾形分支转变,根径、不同径级根系生物量、侧根数均呈现先升高后降低的趋势[18]。柽柳根系空间分布特征、拓扑结构、根冠异速生长关系以及根冠比等对滨海滩涂不同林分密度也具有一定的可塑性[19]。但目前尚不清晰柽柳根系适应地下水矿化度的生长分布规律和响应策略,这在一定程度上影响了对柽柳低效林的抚育提升管理和柽柳幼苗栽植的水盐管理。因此,本研究在1.8 m地下水水位条件下,模拟设置淡水、微咸水、咸水、盐水4个地下水矿化度,以3年生柽柳幼苗为研究对象,测定分析柽柳根系直径、扎根深度、生物量、拓扑指数、连接长度等生长指标和构型特征,探讨柽柳根系形态、构型对地下水矿化度变化的响应规律,探明适宜柽柳幼苗生长的地下水矿化度条件,以期为黄河三角洲地区柽柳幼苗的水盐栽植管理提供技术支撑。
1 材料与方法
1.1 试验材料
本试验在山东省黄河三角洲生态环境重点实验室的科研温室内进行(117°58′57″E,37° 22′56″N)。温室玻璃透光率大于90%,室内空气平均相对湿度为(45±6)%,平均大气温度为(25±4) ℃。在黄河下游滨海滩涂地带挖取粉砂壤土用于试验,粘粒占比为5.76%,粉砂占比为47.66%,砂粒占比为46.58%,土壤类型为潮土,其初始pH值为7.54,平均含盐量为0.1%,田间持水量为37.86%,土壤容重为1.32 g/cm3。柽柳苗木采用3年生实生苗,在材料选择上,选择生长、形态指标基本一致的柽柳幼苗,在栽植前,统一进行了截干处理,每株柽柳苗木有2~3个一级侧根,平均根径为6.0 mm。
1.2 试验设计
黄河三角洲潜水埋深较浅,平均埋深为1.14 m[20],矿化度多介于14.3~32.4 g/L[21],紧邻泥质海岸带的黄河三角洲莱州湾柽柳林场调查研究发现,潜水水位多介于0.3~2.0 m[9]。黄河三角洲地下水矿化度,随着距离海岸带远近的不同,呈现由微咸水到盐水的过度程度,而紧邻黄河岸堤附近的地下水以淡水生境为主,因此,模拟设置了4种地下水矿化度。本研究模拟1.8 m地下水水位,设置4个不同的地下水矿化度:淡水(A,0 g/L);微咸水(B,3 g/L);咸水(C,8 g/L);盐水(D,20 g/L)。地下水矿化度采用黄河三角洲以NaCl为主的海盐配制。每地下水矿化度均设置3个重复,共计12个栽植柽柳的土壤柱体模拟装置。
本研究模拟控制试验设计:在科研温室内,将柽柳栽植于PVC圆管(内径=0.3 m)中,将高、上口直径、底部直径分别为0.70、0.57、0.45 m的水桶作为模拟不同地下水矿化度的装置。为防止模拟地下水受环境温度变化太大,使地下水温度维持在相同水平,将水桶放置于挖好的深度为0.6 m的壕沟内。
栽植柽柳的PVC圆管具体设计参数为:1)根据PVC圆管高度为模拟地下水水位、实际淹水深度(0.55 m)和顶端的空隙层(0.03 m)之和,加工成高度为2.38 m的圆管。实际淹水区的0.55 m PVC管每隔10 cm打4个直径为1.0 cm的小孔作为进水口,并用透水布堵住,淹水区PVC圆管底部用透水布包住,铺反滤层以保证模拟地下水可从底部和四周的进水口充分进入土壤体。2)将土壤样品晾干、混匀后,按照20 cm一层填充土壤柱体,并用皮锤逐一压实。3)将长势均匀一致,基径平均为1.30 cm、苗高平均为0.60 m的3年生柽柳幼苗,栽植于PVC圆管中,每个PVC圆管先栽植3株,并从土壤表面浇灌淡水(4 L/次×3次)。4)30 d后,在每个PVC圆管内随机留下1株成活幼苗,开始进行不同地下水矿化度的模拟处理,并且不再从土壤表面浇水。为保持地下水水位及其矿化度的稳定,整个试验期间对模拟用地下水的盐度及水位每3 d检测一次,并补充相应的水分和盐分。在试验处理810 d后,2017年7月8日—7月20日进行栽植柽柳土壤柱体的水盐参数、根系生长和根系拓扑指数等的测定。具体装置模拟示意图见图1。
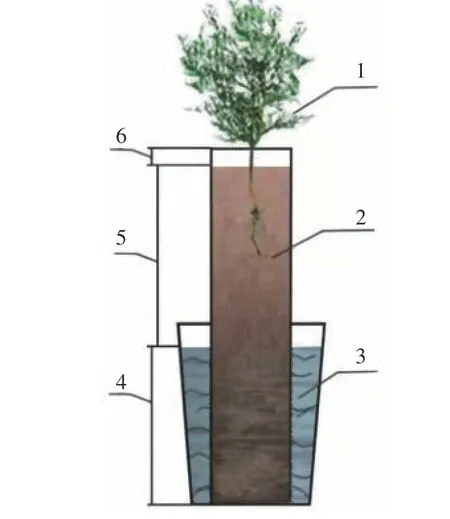
图 1 不同地下水矿化度下栽植柽柳土柱的模拟示意图Fig. 1 Schematic diagram with planting T. chinensis under different groundwater salinity
1.3 测定指标及方法
1.3.1 土壤水盐参数的测定
对栽植柽柳的土柱按照20 cm一层,分层取土壤样品,共采集到1.0 m深。每个PVC圆管每层随机取3个土壤样品混合作为该层的样品,每地下水矿化度处理分别取3个土柱样品作为3次重复。土壤含水量用烘干法测定,土壤盐分用残渣烘干法测定,水土比为5∶1。土壤溶液绝对浓度计算方法[22]如下:

1.3.2 柽柳根系生长参数的测定
将试验4个地下水矿化度处理的共12株柽柳整株挖掘出来。用自来水冲洗根系上附着的泥沙,待根系晾干后,利用电子游标卡尺和米尺等测定以下参数:1)根系内部、外部连接数量、连接长度;2)各级根系的直径,并测量根系长度;3)根系分级数及其生物量。根系生物量的测定:在105 ℃杀青,30 min后在80 ℃烘干至恒质量,称量各部分鲜质量和干质量。
1.3.3 根系拓扑的结构测定
根系拓扑结构不仅决定了植物根系的空间分布特征,而且对根系的营养吸收能力以及固定作用产生重要影响,能在一定程度上反映植物根系在不同生境条件下的觅养策略,尤其是侧根对养分的竞争能力[23]。Bouma等[24]和Fitter等[25]提出了鱼尾形分支和叉状分支2种根系拓扑结构极端类型,并且通过拓扑指数来反映不同植物根系的分支模式。拓扑指数计算公式为:

式中:M为根系所有外部连接的总数;A为最长根系通道内部连接的总数。TI=1.0,为鱼尾形分支,所有分支与主根直接相连,均为外部分支且不再继续分支;TI=0.5,为叉状分支,每一分支点的2个分支所产生的二次分支具有相同角度和数量。
1.4 数据分析
使用Microsoft Excel和Origin 8.0进行数据处理和绘图,采用SPSS 16.0统计分析软件对不同地下水矿化度条件下土壤水盐参数、根系指标进行单因素方差分析(one-way ANOVA),并对其进行T检验(α=0.05)。
2 结果与分析
2.1 不同地下水矿化度下栽植柽柳土柱的水盐参数
由表1可知,随地下水矿化度的升高,土壤含盐量显著升高(P<0.05),淡水、微咸水、咸水矿化度下土壤含盐量分别是盐水矿化度下的12.82%、15.38%、43.59%。随地下水矿化度的升高,土壤含水量呈现升高趋势,但差异不显著。盐水、咸水矿化度下土壤溶液绝对浓度显著高于淡水、微咸水(P<0.05)。

表 1 不同地下水矿化度下栽植柽柳土柱水盐参数Table 1 Water and salt parameters of soil columns planted with T. chinensis under different groundwater salinity
2.2 不同地下水矿化度下柽柳根系的生长指标
由表2可知,地下水矿化度对单株柽柳基径、总根长影响显著(P<0.05)。随地下水矿化度的升高,柽柳基径呈现先升高后降低的趋势。微咸水矿化度下柽柳基径达到最大值,淡水、咸水、盐水矿化度分别比其降低29.15%、17.47%、41.39%。淡水、微咸水矿化度下根长显著高于咸水、盐水矿化度,表现为微咸水>淡水>咸水>盐水。淡水、微咸水、咸水矿化度下柽柳一级侧根数差异不显著,但均显著高于盐水矿化度下侧根数(P<0.05)。
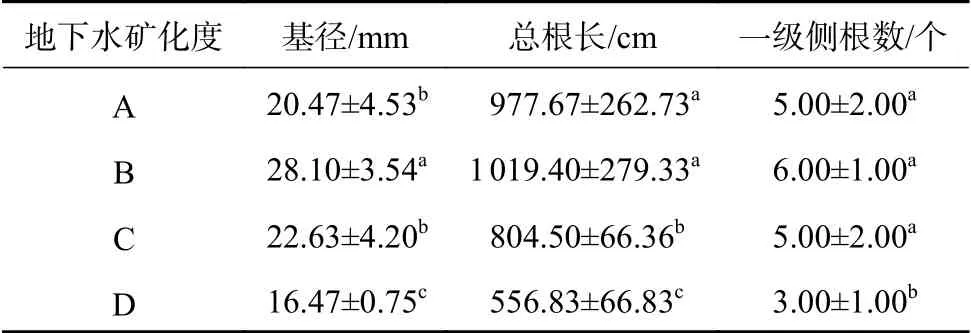
表 2 不同地下水矿化度下柽柳根系的生长指标Table 2 Root growth indexs of T. chinensis under different groundwater salinity
2.3 不同地下水矿化度下柽柳根系生物量的分配
由图2可知,随地下水矿化度的升高,单株柽柳主根生物量、一级侧根生物量呈现先升高后降低的趋势,在微咸水矿化度下达到最大值。淡水、咸水、盐水矿化度下单株柽柳主根生物量分别比微咸水矿化度下降低3.68%、42.43%、45.94%,一级侧根分别降低3.99%、47.81%、70.38%。随地下水矿化度的升高,单株柽柳二级侧根生物量、毛细根生物量整体呈现降低趋势,微咸水、咸水、盐水矿化度显著低于淡水矿化度(P<0.05),二级侧根生物量分别是其2.26、1.97、3.08倍,毛细根生物量分别是其1.64、2.13、2.18倍。

图 2 不同地下水矿化度下柽柳不同径级根系生物量Fig. 2 Root biomass of T. chinensis with different diameter classes under different groundwater salinity
由图3可知,淡水矿化度下单株柽柳地上部生物量显著高于咸水、盐水矿化度下(P<0.05),但与微咸水矿化度下差异不显著,微咸水、咸水、盐水矿化度下分别是淡水矿化度的1.35、1.76、2.12倍。随地下水矿化度的升高,单株柽柳根系总生物量呈现下降趋势。根冠比先升高后降低,表现为微咸水>淡水>咸水>盐水。
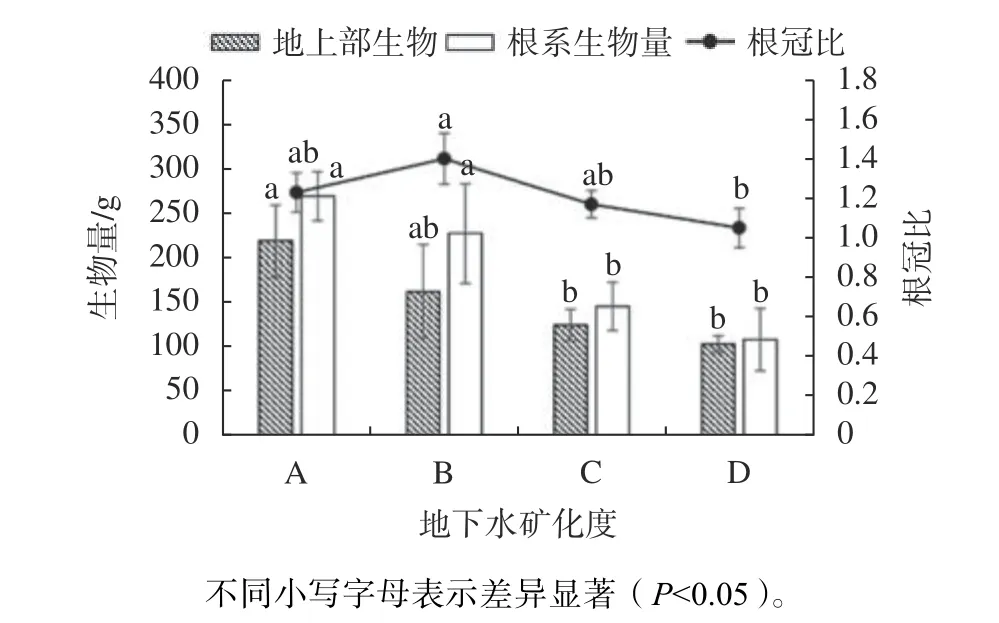
图 3 不同地下水矿化度下柽柳地上生物量、根系生物量及根冠比Fig. 3 Aboveground biomass, root biomass and root-shoot ratio of T. chinensis under different groundwater salinity
2.4 不同地下水矿化度下柽柳根系的拓扑指数
根系拓扑指数可以反映不同地下水矿化度处理下柽柳根系间拓扑结构的差异性(表3)。不同地下水矿化度处理下,柽柳根系拓扑指数差异显著(P<0.05),根系拓扑指数均值表现为盐水>咸水>微咸水>淡水。在淡水、微咸水低矿化度下,柽柳根系拓扑指数较小接近0.5,表明根系分支结构复杂,根系分支接近叉状分支模式;在咸水、盐水高矿化度下,柽柳根系拓扑指数较大都接近1.0,根系分支结构较为简单,趋向于鱼尾形分支模式。随地下水矿化度的升高,根系连接长度显著下降,微咸水、咸水、盐水矿化度下分别是淡水矿化度下的86.79%、55.97%、47.33%。

表 3 不同地下水矿化度下柽柳根系拓扑参数和连接长度Table 3 Topological parameter and link length of root system of T. chinensis under different groundwater salinity
3 结论与讨论
根系生长是反映幼苗受盐分胁迫的敏感指标[26-27]。微咸水矿化度下,单株柽柳基径、总根长、侧根数均高于淡水条件下,在微咸水矿化度下达到最大值。柽柳作为一种典型的泌盐性盐生植物,适当提高地下水矿化度可刺激柽柳根系的生长。在咸水、盐水等高矿化度下,柽柳基径和侧根数显著下降,高盐环境抑制了基径和侧根的生长。但也有研究发现,水盐胁迫下不同植物根系侧根数会显著增加或降低,这种差异可能与不同试验的植物材料、水盐胁迫强度及盐分类型等因素有关[28-29]。
自我调节生物量配置模式是植物适应盐胁迫的可塑性机制之一[30], 植物能通过改变根、茎、叶的生物量分配比例来适应不同盐分环境[31]。随地下水矿化度的增加,单株柽柳地上生物量、根系总生物量均呈现下降趋势,根冠比先升高后降低,但是根系生物量下降幅度要大于地上部生物量,可见根系对于土壤盐分的响应比植物地上部分器官更为敏感,受到更大的抑制。滨海盐碱地桑树(Morus alba)幼苗也被发现盐胁迫下根系比地上部反应敏感,尤其是根系的形态指标,较低的含盐量就能显著抑制根系的发育[27]。水盐逆境下柽柳通过对地上部器官和根部生物量分配的调控,可以使柽柳更好的平衡资源的获取和利用。 柽柳主根、一级侧根、二级侧根、毛细根生物量的变化,在根系固土能力、根系营养物质的吸收、植物生长发育方面发挥重要作用[32]。随地下水矿化度的升高,土壤含盐量和土壤溶液绝对浓度显著升高,柽柳主根、一级侧根生物量呈现先升高后降低的趋势,在微咸水矿化度下达到最大值,而二级侧根、毛细根生物量持续下降。这说明适当的低盐条件可促进柽柳主根、侧根的生长,而毛细根可能因为对盐分的敏感性更高而受到抑制。而高盐环境对柽柳主根、侧根和毛细根的生长均产生胁迫作用,根系作为植物吸收盐分的主要部位,为了保证柽柳在高盐环境下继续生长,通过减少根系生物量以降低对盐分的吸收,进而使得盐离子输送变得相对缓慢,有效避免了盐胁迫伤害[33]。但黄河三角洲芦苇(Phragmites australis)根系主根生物量受盐分影响并不显著[34],可见不同植物对于盐分胁迫的适应性差异较大。
Fitter等[25]提出利用拓扑指数(TI)大小来反映根系构型,并将根系构型划分为鱼尾形分支和叉状分支两种极端模式。在淡水、微咸水低矿化度下,土壤含盐量和土壤溶液绝对浓度均较低,水盐条件适宜,柽柳根长增加,根系分支趋向于叉状分支模式,主要通过扩大根系分布范围、增加次级分支形成一种网状结构,虽然根系分支丰富,但单位资源投入下根系延伸距离短,这样的根系分支特征更适合在资源相对丰富的土壤环境[35]。在咸水、盐水高矿化度下,土壤含盐量和土壤溶液绝对浓度均较高,受其影响,柽柳根长降低,根系趋向于鱼尾形分支模式。土壤盐分抑制土壤营养物质的转化与积累,同时高盐分条件下土壤酶活力降低抑制微生物活性,导致土壤贫瘠[35],造成根系拓扑指数显著升高。高地下水矿化度下,柽柳降低了根系分支的数量,以增加单位资源投入根系的延伸距离进行水分、养分的收集利用[36],这与Bouma等[24]的研究结果类似。
随地下水矿化度的增加,土壤盐分随之增加,柽柳根系长度增加幅度减小,根系连接长度显著下降,柽柳采取减少根系分布范围的避性机制,以最大限度地降低对高含盐量的毒害。随盐胁迫的增强,平欧杂种榛(Corylus heterophylla×C.avellan)幼苗根系长度下降,这与本研究结果相一致[37]。植物根系形态特征对盐分胁迫的响应规律与根系的生理生态调节机制密切相关[27,38-39],在遭受盐胁迫时,植物常常通过抗氧化系统中的酶类和非酶类抗氧化物质之间的协同作用来抵御盐胁迫引起的氧化胁迫[26]。研究发现,秋茄(Kandelia candel)幼苗是通过根系功能较强的抗氧化系统清除活性氧以提高植株的耐盐性[38]。盐旱胁迫下塔里木河下游铃铛刺(Halimodendron halodendron)、疏叶骆驼刺(Alhagi sparsifolia)、和胀果甘草(Glycyrrhiza inflata)根解剖结构中皮层厚度变薄、维管柱变厚[36]。柽柳根系生长及构型对地下水-土壤系统的水盐条件响应敏感,其内在生理生态过程也必然发生变化,在下一步的研究中,应对柽柳根系生长及生理生态变化过程开展系统性探讨,以丰富和完善柽柳根系生理生化指标在耐盐中的生理响应机制。
土壤含盐量、土壤含水量、土壤溶液绝对浓度受地下水矿化度影响显著,并对柽柳根系生长及构型产生影响。随地下水矿化度的升高,柽柳基径、根长、侧根数、主根生物量、侧根生物量均呈现先升高后降低的趋势,根系分支呈现由叉状分支向鱼尾形分支模式的转变。不同地下水矿化度下柽柳根系外部连接长度大于内部连接长度,柽柳根系均呈现向外扩张的生长策略。柽柳通过调整根系生长特征、拓扑结构以及生物量分配比例来适应不同的地下水矿化度条件,适当提高地下水矿化度至微咸水可促进柽柳根系的生长,而高于咸水矿化度则会抑制柽柳根系生长。
