湖南栎类天然林林分结构对林分生长的影响研究
2022-10-17李新建杨子铎何海梅邱子阳朱光玉
李新建,杨子铎,何海梅,邱子阳,朱光玉
(1.国家林业和草原局中南林业调查规划设计院,长沙 410014;2.中南林业科技大学 林学院,长沙 410004)
林分生长量是衡量森林质量和数量的重要指标,不仅反映了一段时间内森林能够产出的木材量、碳储量,而且可以作为评估森林生产力、环境质量和衡量林分生长健康情况的重要指标[1-2]。林分生长量,按照调查因子分为直径、树高、断面积和蓄积生长量等。然而,由于林分生长存在自然稀疏的现象,相比于树木“纯生型”的生长过程,林分“生灭型”的生长过程更加复杂[3]。目前,对于林分生长的影响因素已有大量研究,一般认为,立地条件和林分结构是关键的影响因子,而相比于立地条件,林分结构因素可调控性更高[4-6]。
林分结构是森林生态系统演替的驱动因子,是评价森林生态效应的重要因素之一,也是森林生态系统管理研究的理论基础。拥有合理的林分结构,才能充分发挥森林生态系统的各项功能[7-8]。优化林分结构,是确保森林生态系统生物多样性维持和增加的基础,是精准提升森林质量的有效途径[9]。目前,国内外学者对林分结构已经进行了大量的研究,但对于林分结构的分类仍有待进一步探讨。有的研究将林分结构分为非空间结构与空间结构两种[10]:非空间结构主要是指树种组成、树高结构、直径结构、林分密度、年龄结构等;林分的空间结构是指林分中树木的空间分布格局及其相关属性的分布方式[11]。赵中华等[12]从林分组成、大小分化多样性、空间分布多样性3个方面对林分结构指标进行了系统梳理和总结,而Miren等[13]将林分结构的指标分为8类。这些学者均认为林分结构对林分的生长有显著影响。
栎类林占中国天然林总面积的14%,是中国天然林的主要组成部分,也是湖南的一种典型森林群落,以中幼龄林为主。其在涵养水源、保持水土以及维持生态稳定等方面发挥了重要作用,具有很高的研究价值。以往关于该类型植被的研究主要集中在生长预估[14],林分结构对林下植被生长量的影响[15]及林分更新特征[16]等方面,而林分结构因子对林分生长的影响规律研究鲜见报道。有基于此,本研究根据在湖南设置的49块栎类天然林固定样地数据,依据Miren等[13]对林分结构指标的划分方法,将林分结构划分为林分密度、物种组成、水平结构、垂直结构、大小分布、林分年龄结构6大类,选取22个林分结构指标,采用平均树高、胸高断面积、蓄积量的总平均生长量3个指标,利用Pearson相关分析筛选影响林分生长量的关键因子,并在此基础上构建结构方程模型,探索林分结构对其生长量的影响规律,以期为天然林的科学经营管理提供参考。
1 研究区概况
研究区位于湖南省,地处长江中游,三面环山,与多个省份交界。全省年日照时长1 500h左右,多年平均降水量在1 450 mm,富含各种资源。土壤主要以红壤和黄壤为主。森林面积1 053万hm2,活立木蓄积4.61亿m3,森林覆盖率49.69%。湖南省植物种类多样,群种丰富,其中栎类林面积和蓄积约占全省阔叶林的12%和11%~14%。研究区主要乔木树种有锥栗(Castaneahenryi)、甜槠(Castanopsiseyrei)、石栎(Lithocarpusglaber)、枹栎(Quercusserrata)、青冈栎(Quercusglauca)、多脉青冈(Quercusmultinervis)、细叶青冈(Quercusgracilis)、光叶水青冈(Faguslucida)、香樟(Cinnamomumbodinier)、鹅耳枥(Carpinusturczaninowii)等。
2 研究方法
2.1 样地设置与调查
2015年12月—2016年8月,分别在湖南省平江县芦头林场、桑植县八大公山自然保护区、益阳市龙虎山林场、郴州市五盖山林场、宁乡市青羊湖林场5个地方进行外业调查,运用典型抽样技术在具有代表性的典型地段设立样地进行调查。设置了20m×30m的以栎类为主要优势群落的固定样地49块。对样地内胸径大于或等于5cm的活立木进行每木检尺,主要林分调查因子包括胸径、树高、冠幅、林分年龄、平均优势木高、郁闭度、地理坐标、海拔、坡向、坡度、坡位、土壤类型、土壤厚度等。样地中确定林分年龄的方法是利用生长锥钻取木芯计数年轮,以样地内树种组成最大的优势树种的平均木年龄代表混交林的年龄(T),样地具体信息如表1所示。
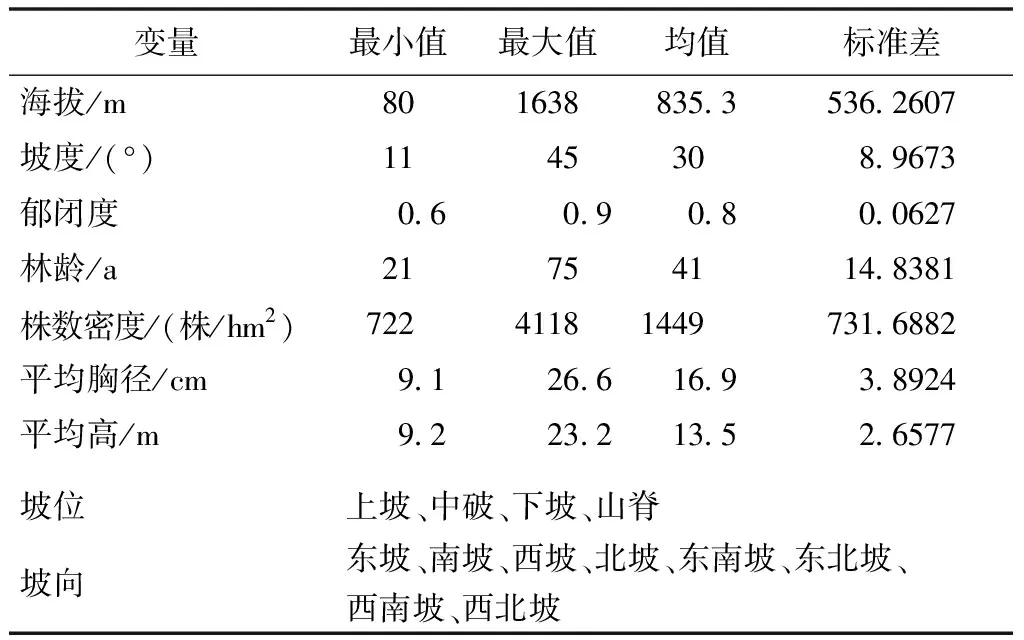
表1 样地的基本情况
2.2 林分空间结构单元的确定
本文采用4株木法确定林分空间结构单元。为避免核心木的相邻木落在样地之外,影响结构指数的计算,在20m×30m样地四周设置了3m的带状缓冲区以消除边界影响。
2.3 林分结构指标
选取株数密度(N)、林分密度指数(SDI)、拥挤度(K)、密集度(Ci)、郁闭度(N)代表林分密度指标;选Shannon-Wiener多样性指数(H′)、Pielou的均匀度指数(E)、Margalef丰富度指数(R1)、Simpson多样性指数(D)代表林分的物种组成;选取聚集指数(R)、角尺度(Wi)、混交度(Mi)代表林分的水平结构指标;选取树冠指数(CI)、开敞度(Bi)、林层比(Si)、林层差异化(Di)代表林分的垂直结构指标;选取竞争指数(Hegyi)、大小比数(Ui)、胸径大小变异系数(Dcv)、胸径分布偏度(sk)、断面积Gini系数(GC)代表林分的竞争及大小分化程度指标,以及林分年龄(T)共22个参数作为林分结构指标[12-13,17]。
2.4 林分生长量指标
采用林分平均树高、胸高断面积、蓄积量的总平均生长量(θH,θDg,θv)来表示树木的生长。以蓄积量为例,总平均生长量计算公式如下:
(1)
式中:Vt为样地蓄积总量,t为林龄。
2.5 林分结构对林分生长量的影响分析
研究利用Winkelmass、Excel 2016与R 4.0.5分析工具,计算固定样地林分结构指与林分生长量指标,并基于此,利用R 4.0.5与Amos 24.0分析林分结构与林分生长量的相关关系。
2.5.1Pearson相关分析
采用Pearson 相关系数判断林分结构指标与林分生长量指标之间的单因素相关性,并筛选出关键的影响因子,其计算公式为:
(2)
2.5.2结构方程模型的构建
结构方程模型(SEM)是基于变量的协方差矩阵,采用传统的多元数据分析方法来建立、估计和检验变量之间的相互作用关系的方法。该方法可以有效地处理多变量之间的作用关系强度和影响传导路径,相比于传统统计方法具有很大优势[18]。
结构方程模型构建前,先做预处理,利用Z-score方法对原始数据进行标准化处理,并以方差膨胀因子(VIF<10)为标准进行线性诊断,删去了存在多重共线性的变量。最终将物种组成、垂直结构、林分密度作为外源潜变量。将Shannon-Wierner多样性指数、Pielou均匀度指数、Simpson多样性指数作为物种组成的可测变量;将林层差异化、林层比、开敞度作为垂直结构的可测变量;将株数密度、密集度、郁闭度作为林分密度的可测变量;将平均树高、胸高断面积和蓄积量的总平均生长量作为与内生潜在变量有关的观测变量;将林分年龄、胸径大小变异系数、混交度作为独立变量加入模型,预设结构方程路径关系,构建了林分结构对林分生长影响的概念模型(图1)。
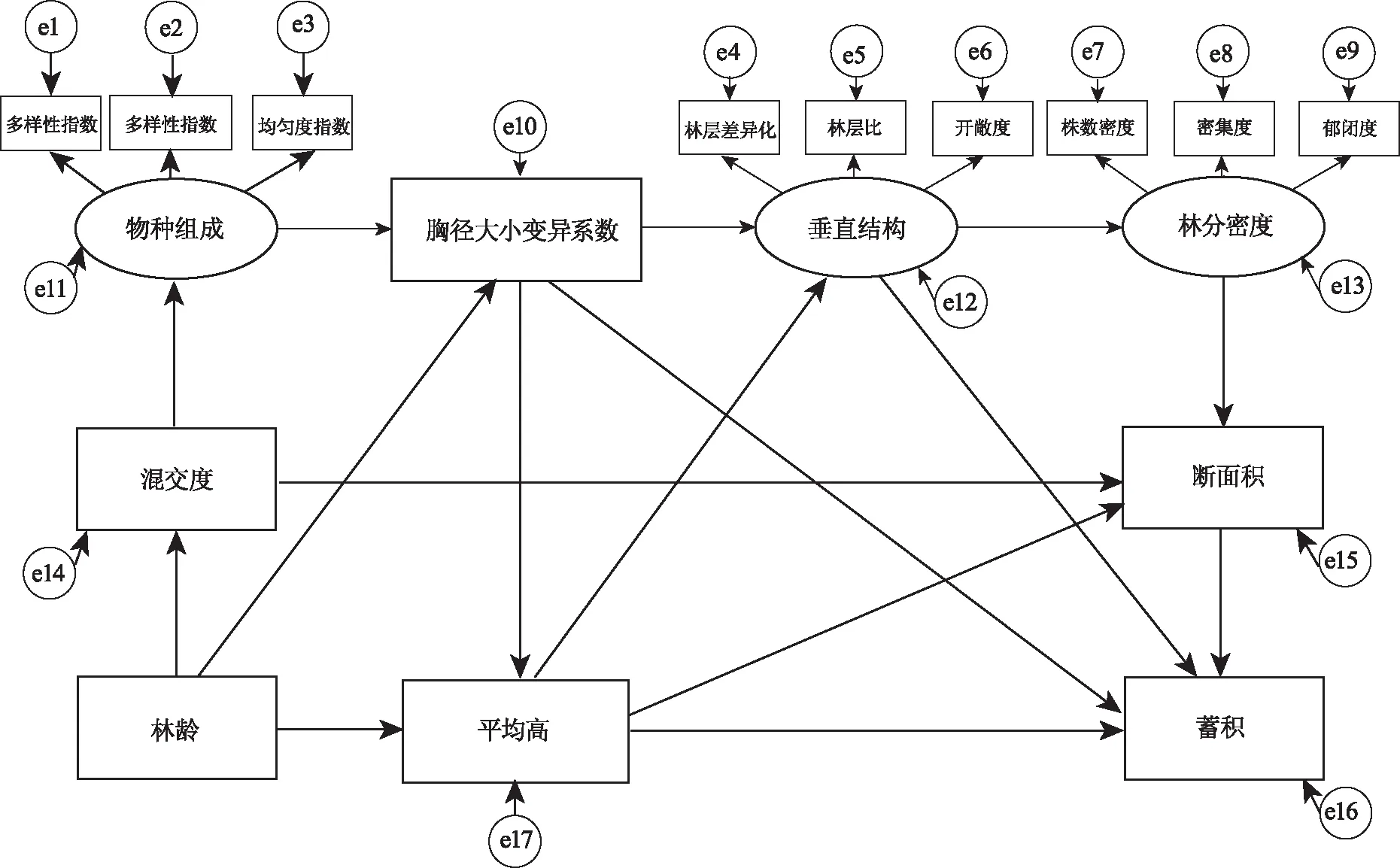
注:连接两个指标的直线表示两个指标间的直接影响,箭头表示影响方向,直线上的值表示为标准化路径系数;e1—e17为可测变量的误差变量。
2.5.3模型的适配度检验
为了确定最佳结构方程模型,研究采用卡方自由度比X2/df(Chi-square degrees of freedom)、标准拟合指数(Comparative Fit Index,CFI)、递增拟合指数(Incremental Fit Index,IFI)和渐近残差平方和的平方根(Root Mean Square Error of Approximation,RMSEA)等4个拟合指数来评价模型拟合效果[19]。X2/df介于1~3之间,表示模型适配度良好。CFI与IFI的数值介于0~1之间,其数值愈接近1,表示模型的适配度愈佳,一般的判别标准为CFI,IFI值大于0.9,表示模型的路径图与实际数据有良好的适配度。RMSEA值介于0~1间,其数值愈大表示模型的适配度愈差,其值为0时,表示模型有完美的适配度,一般而言,模型适配度可接受的范围为其值在0.1以下。本研究结构方程模型的构建及检验均使用Amos 24.0完成。
3 结果与分析
3.1 各项指标的计算结果与分异特征
由49块样地的林分结构指标与林分生长量指标统计信息可以看出,湖南栎类天然林各个样地间株数密度、Margalef丰富度指数、树冠指数、Hegyi竞争指数和胸径分布偏度的变化较大,变异系数CV分别为50.50%,49.66%,91.53%,44.07%和64.09%(表2)。

表2 49块样地林分结构与林分生长量指标的统计信息
3.2 林分结构与林分生长量的单因子相关分析
对22个林分结构指数和3个林分生长量指标进行Pearson相关性检验,结果如表3所示。
表3结果显示:
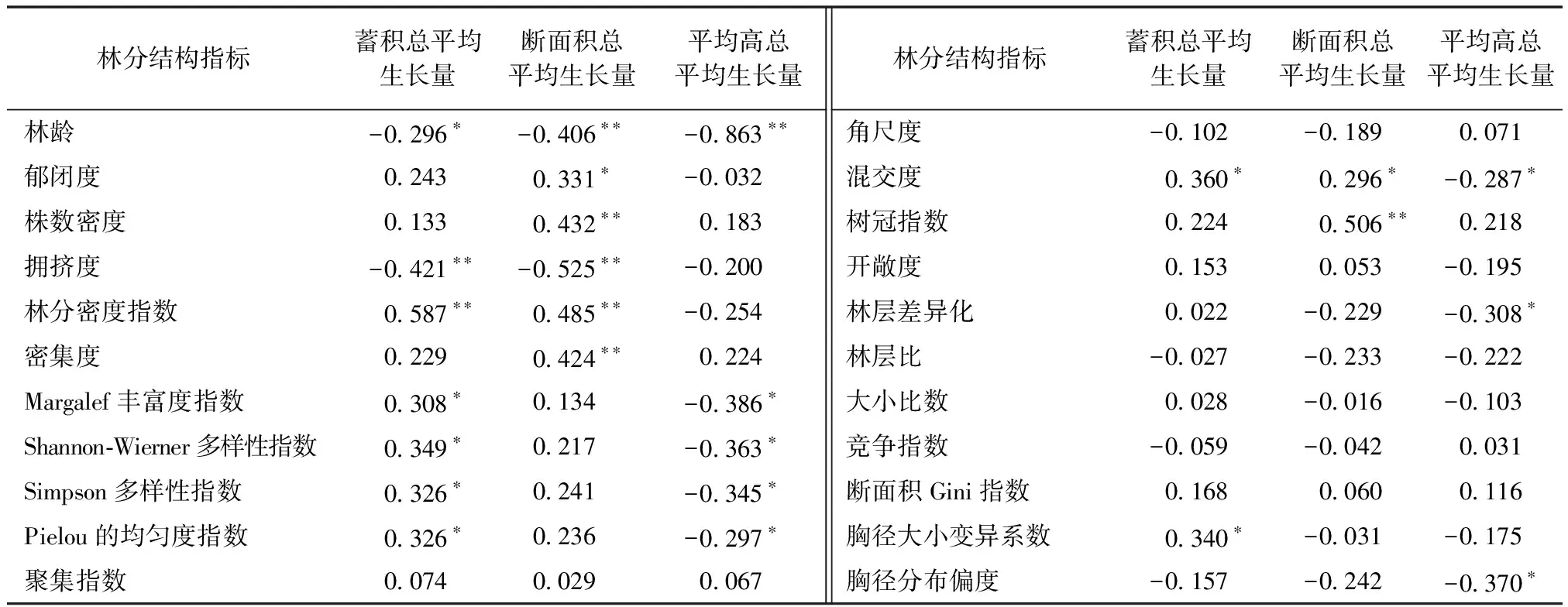
表3 林分结构与林分生长Pearson相关系数
1)林分年龄与蓄积总平均生长量呈显著负相关关系(P<0.05);拥挤度与蓄积总平均生长量呈极显著负相关关系(P<0.01);林分密度指数与蓄积总平均生长量呈极显著正相关关系(P<0.01);物种组成、混交度、胸径大小变异系数与蓄积总平均生长量呈显著正相关关系(P<0.05)。
2)林龄、拥挤度与断面积总平均生长量呈极显著负相关关系(P<0.01);郁闭度、混交度与断面积总平均生长量呈显著正相关关系(P<0.05);株数密度、林分密度指标、密集度、树冠指数与断面积总平均生长量呈极显著正相关关系(P<0.01)。
3)林龄与平均高总平均生长量呈极显著负相关关系(P<0.01);物种组成指标、混交度、林层差异化、胸径分布偏度与平均高总平均生长量呈显著负相关关系(P<0.05)。
3.3 结构方程模型结果与分析
由于林分结构间也存在复杂的相互作用关系,因此利用Pearson相关分析筛选出的关键因子,预设结构方程路径关系,构建结构方程模型,进一步分析林分结构对林木生长量的直接影响、间接影响和总影响。
模型的适配度检验结果显示,X2/df为1.087,CFI为0.990,IFI为0.991,RMSEA为0.043,说明构建的结构方程模型与数据的适配度良好,模型精度较高,模型拟合基本理想,确定初始模型为林分结构-林分生长的最优模型。
结构方程模型结果(表4)显示林龄对3种生长量指标均具有极显著的影响(P<0.001),影响系数分别达到-0.867,-0.42,-0.359;混交度、林分密度对断面积总平均生长量具有极显著的影响(P<0.001),影响系数分别达到0.489,0.351;混交度、胸径大小变异系数、林分密度对蓄积总平均生长量具有极显著的影响(P<0.001),影响系数分别达到0.515,0.443,0.253;物种组成对蓄积总平均生长量具有显著的影响(P<0.01),影响系数分别达到0.189。
结构方程模型路径分析结果(图2、表4)显示,混交度和物种组成是通过间接、直接影响胸径大小变异系数的方式来间接影响平均高总平均生长量;林龄、物种组成、胸径大小变异系数、垂直结构是通过直接影响平均高总平均生长量、间接影响林分密度的方式来间接影响断面积总平均生长量;林龄、混交度、林分密度、物种组成是通过直接影响平均高总平均生长量、断面积总平均生长量和胸径大小变异系数的方式来间接影响蓄积总平均生长量。

注:连接两个指标的直线表示两个指标间的直接影响,箭头表示影响方向,直线上的值表示为标准化路径系数;e1—e17为可测变量的误差变量;“*”表示路径检验P<0.05,“**”表示P<0.01,“***”表示P小于0.001。

表4 林分结构各指标对林分生长量的直接 间接和总影响
4 结论与讨论
4.1 Pearson相关分析
探索林分结构与林分生长的关系已成为林学领域研究的一个热点问题,了解林分结构如何影响天然林的演替与生长,对确定天然林森林经营与改造措施意义重大[20]。大量的研究表明,林分结构对林分生长的影响,不仅归因于林分密度和树种结构,还依赖于林分空间结构、林木大小分布状况以及林分生长发育阶段,但其对林分生长的相对重要性和影响机制仍不明确,值得进一步研究[13,17]。本研究利用Pearson筛选关键结构因子,用混交度表征林分水平空间结构,用林层指数、林层比和开敞度表征林分的垂直空间结构,用Shannon-Wierner多样性指数、Pielou均匀度指数、Simpson多样性指数表征物种组成,用株数密度、密集度、郁闭度表征林分密度,用胸径大小变异系数表征林木的大小分布,探讨了其对林分生长的影响的相对重要性以及作用机制,研究结果显示,混交度、林分密度是通过直接影响断面积生长来间接影响蓄积生长,且影响达到了极显著水平(P<0.001),这与张毅峰[21]、秦建华等[22]的研究结论基本一致。张毅峰[21]研究发现,天目山天然林林分断面积生长量随着混交度的变大而随之增大;秦建华等[22]研究发现,林分密度是影响林分生长与产量的主要因子。究其影响机制,多树种混交的林分,各树种在生态位上相互抑制,降低了林木的竞争程度,树木生长发育空间越充足,林木断面积越大,进而影响林分蓄积。而林分密度对断面积生长的影响则体现在林分中的植株数量越多,断面积越大。
4.2 结构方程模型
研究结果显示,胸径大小变异系数极显著地影响蓄积生长(P<0.001),这与刘铭波[23]的研究结论类似。刘铭波[23]研究发现,大小比数是辽东栎林生长的主要结构驱动因子,对辽东栎林应首先关注林木间胸径大小差异程度。这可能是林分大小分布与分化差异状况直接影响着林木间的竞争和生长潜力所致,一方面,林木大小分化程度越大的幼中龄林,小径级的树木越多,由于处于生长早期的小径级树木生长速度较快,其更易积累林分有机物质,也就更利于林分生长[24];另一方面,增加林木大小分化差异性,可以提高不同林冠层的林木填充密度,进而促进林分生长[25]。
研究中也发现,物种组成是通过直接影响林木大小分化来间接影响蓄积生长,影响达到了显著水平(P<0.01)。周健平[26]在进行阿什河流域天然林蓄积量影响因子分析时,也得到了相同的结论,即树种多样性增加对全林分蓄积量有着显著的正向影响。这可能与林分中不同树种间的相互作用有关。一方面,树种多样性的增加,特别是一些速生伴生树种的增加,促进了群落稳定性,进而促进林分生长;另一方面,植物群落中不同的物种会发生种间竞争,群落物种组成不同,竞争情况也会发生变化,这种竞争关系的变化会导致植株个体在大小上的差异分化,进而影响林分生长[27]。究其原理,物种组成对植物生长的影响取决于生态位的互补与分化(生境中不同物种在资源利用上的差异程度),生态位分化通过影响个体生长可塑性来改变植株的大小分化[28],进而影响植株生长。
森林是一个复杂的自然系统,描述林分结构对林分生长的影响关系时,需要考虑林分结构间的相互关系和林分结构影响林分生长过程间的相互作用,同时分析多个过程的影响因子和变化趋势,才能科学揭示森林生长过程的真实机理。相比传统一对一简单关系描述的统计方法,结构方程模型在解决生态系统耦合关系问题上更具有优势,它可以通过直接作用和间接作用描述系统间复杂的相互关系[29]。研究表明,结构方程模型能够从整体上直观地反映林分结构对林分生长的影响。结构方程模型不仅定量描述了产生的影响,也直观地展现了其影响路径,整体上比较科学地揭示了林分结构对林分生长影响的复杂作用。
4.3 讨论
树种混交度、林木大小分布状况、林分密度和树种组成是影响湖南栎类天然林林分生长的主要因素,其中混交度的影响最大。一方面林分混交程度越高,林木大小分化差异越大,林木竞争程度降低,其营养空间越充足,林木生长潜力越大;另一方面树种多样性的增加,特别是速生伴生树种的增加,促进了群落稳定性。因此,欲促进栎类天然林林分生长,应选择以调整树种混交度为主,合理控制林分密度和林木大小分布状况为辅,并适当增加树种多样性的综合经营措施。
