不同建群蜂数对肿腿蜂子代性比的影响*
2022-10-15唐艳龙王丽娜王艳芹张彦龙王小艺
唐艳龙 王丽娜 王艳芹 张彦龙 王小艺 魏 可
(1.遵义师范学院生物与农业科技学院 贵州省赤水河流域动物资源保护与应用研究重点实验室 遵义 563002;2.中国林业科学研究院森林生态环境与自然保护研究所 国家林业和草原局森林保护学重点实验室 北京 100091)
对性比可调节的动物而言,性别分配行为是母代对产雄或产雌生殖权衡的结果,是决定种群两性数量结构的基础。关于动物的性比结构,Fisher最早提出两性数量均等是一种进化稳定策略(Fisher, 1930)。然而在寄生蜂类群中,自然界中满足雌雄等比结构的种类往往缺乏充足的证据(Odeetal., 2008),因为Fisher提出的这一理论需要建立在个体交配行为随机发生的基础上。事实上,多种寄生蜂的雌雄数量并非等比关系,特别是针对群居型寄生蜂(gregarious parasitic wasps),偏雌性比是它们更常见的生物学特征(King, 1987; Tangetal., 2014; Boultonetal., 2015; Kapranasetal., 2016; Sreenivasetal., 2016)。Hamilton在20世纪中后期提出了在空间受限时寄生蜂偏雌性比的自然选择和遗传基础。Hamilton(1967)认为在受限的空间中,雄性个体间存在竞争交配对象的情况,且这种竞争往往还发生在具有亲缘性的雄性间(兄弟间),即产生局部配偶竞争(local mate competition, LMC)。若雄性个体过多,且没有足够的雌性对象,未完成交配的雄性子代对亲代来说便是一种适合度损耗。因此,为了避免局部配偶竞争而引起亲代适合度降低,母蜂往往仅产出数量上可满足雌蜂交配的雄蜂即可。另一方面,空间限制也容易造成近交行为频发,这导致了母蜂与子代雌蜂亲缘性更强,故母蜂偏向于越来越多地产出雌性后代(Hamilton, 1967)。
群居型寄生蜂偏雌性比是较常见的生物学特征,但它们的性比并非是一个稳定不变的常量。例如,当在同一寄主上产卵母蜂(建群蜂)数量变化时,子代性比通常会发生变化。Hamilton(1967, 1979)基于寄生蜂的局部配偶竞争和近交行为提出了2个数学模型,分析了当建群蜂数量发生改变时寄生蜂子代的性别分配规律。在这2个模型中,建群蜂数量少时会得到更偏雌的子代性比,当建群蜂数量增加时子代性比偏雌程度会被削弱,即总体上认为子代雄性比与建群蜂数量正相关。Hamilton提出的LMC理论及其性比预测模型是寄生蜂性比特征研究的经典理论。LMC理论提出建群蜂数量变化时,子代性比会随之改变,体现了亲代母蜂对子代性比结构的调节能力。LMC模型在很多寄生蜂中得到了验证(Shukeretal., 2005; Shuker, 2006; Marteletal., 2010; Boultonetal., 2019),但其高度适用性的证据大多来源于无社会习性的膜翅目昆虫。
膜翅目昆虫中性别决定的祖先模式是产雄孤雌生殖,在此模式中二倍体雌性由受精卵发育而来,单倍体雄性由未受精的卵发育而来(Heimpeletal., 2008)。单双倍型的性别决定方式使得亲代可以仅通过控制卵是否受精就能实现子代性比调节。调节子代性比是亲代为了实现适合度最大化的一种生殖策略,是亲代对生存环境权衡结果的体现。研究寄生蜂的性比结构,以及其在不同建群蜂密度下性比的调整规律是评价寄生蜂适合度的重要方式。另外,在规模化繁育寄生蜂时,研究人员往往想繁育出更多的具有控害作用的雌蜂,因此性比也是评价繁育效率的重要依据(Wuetal., 2003; Odeetal., 2008; 唐艳龙等, 2020a)。尽管想要通过增加建群蜂的数量来获得更多的子代,然而建群蜂增加时可能会出现如局部配偶竞争模型所预测的雄性子代数量过度增多。上述这一现象在一些寄生蜂的工厂化繁育中常有发生(Heimpeletal., 2000; Pandeyetal., 2005)。因此,评价建群蜂数量与子代性比的关系对寄生蜂繁育也十分必要。
本研究选择了在我国天牛和吉丁虫类林木蛀干害虫防治中广泛应用的管氏肿腿蜂(Sclerodermusguani)、白蜡吉丁肿腿蜂(S.pupariae)和松墨天牛肿腿蜂(S.alternatusi)为研究对象,这一类寄生蜂具有典型的产雄孤雌生殖特征(杨忠岐等, 2014; 陈然等, 2019; Weietal., 2017),即亲代母蜂产雄或者产雌是通过控制卵是否受精而完成,亲代母蜂具备调节子代性比的能力。在众多可能影响寄生蜂子代性比的因素中,建群蜂数量是重要的因素之一(Lietal., 2014; Tangetal., 2014)。不同建群蜂数代表了亲代单个母蜂在固定寄主中可获得净资源的差异,由此可能会形成差异巨大的子代性比。本研究中,笔者系统地研究了3种硬皮肿腿蜂的性比特征,记述了当建群蜂数量变化时它们的子代性比的变化规律。同时,因Hamilton提出的LMC模型概括了寄生蜂性比随建群蜂数量变化的总体关系,但该模型是否能准确地表达不同建群蜂数量下肿腿蜂的子代性比变化规律尚且未知。因此,本研究将3种肿腿蜂在不同建群蜂数量下的性比实测值与LMC性比模型进行了对比研究,评价了该模型在预测肿腿蜂性比结构中的适用性和精度。研究结果对明确肿腿蜂的性别分配和性比调节规律,以及扩展局部配偶竞争理论具有理论意义,对肿腿蜂人工繁育亦有参考价值。
1 材料与方法
1.1 供试寄主和肿腿蜂接种 管氏肿腿蜂、白蜡吉丁肿腿蜂和松墨天牛肿腿蜂实验室种群均已连续繁殖10代,繁蜂寄主为麻竖毛天牛(Thyestilla gebleri)。选用5日龄已充分交配的雌性肿腿蜂和体质量在0.25~0.30 g的麻竖毛天牛幼虫供试,各处理间寄主体质量经检验无显著差异。将寄主单头装入玻璃指形管中(φ=1.0 cm,H=5.0 cm),随后在指形管内接入肿腿蜂雌蜂。前期研究发现肿腿蜂需补充营养(常为吸食寄主血淋巴)后才会产卵。补充营养后的肿腿蜂腹部可见明显膨大,腹部分节清晰可见。本研究分别在单头寄主上接种肿腿蜂1~8头,观察到所有母蜂腹部有明显膨大的样本才记入重复数,保证接蜂数即建群蜂数。每个建群蜂密度重复20次,即每种肿腿蜂计160次重复,3种肿腿蜂共计480次重复。在25 ℃,RH=50%~60%,L∶D=8 h∶16 h的人工气候箱内繁殖肿腿蜂。
1.2 不同建群蜂数量下子代性比及变化规律 接蜂约30天后子代蜂开始羽化,待羽化完成后分别统计不同建群蜂数量下3种肿腿蜂的子代雌蜂和雄蜂的数量。用雄性比(雄性子代数量占子代总数的百分比)计作子代性比,分析不同建群蜂数量下3种肿腿蜂性比的变化规律。同时,进一步统计在不同建群蜂数量下3种肿腿蜂的单雌子代数,并拟合单雌子代数与性比的数量关系。
1.3 LMC模型对肿腿蜂性比的预测精度 Hamilton以建群蜂数量变化会对子代性比产生影响为基础提出的2个LMC理论性比模型分别为:
(I)x= (n-1)/2n;
(II)x= [(c-1)×(n-1)]/2nc。
其中x为雄性比,n为建群蜂数量。模型I为未考虑近交时雄性比随建群蜂数量的变化规律; 模型II考虑了近交,c为近交系数,c= 4N-1,N为自然种群建群蜂数量。笔者首先用模型I预测不考虑近交发生时在不同建群蜂数量下肿腿蜂子代性比; 随后设定肿腿蜂的自然种群建群蜂数量分别为1头、2头和3头(即N=1, 2, 3)的情形,用模型II来预测当近交发生时的肿腿蜂性比。通过拟合在不同建群蜂数量下肿腿蜂实测性比的变化趋势,并同时比较实测性比与基于LMC模型的性比预测值的差异,评估LMC模型在肿腿蜂性比预测中的精确度和适用性。
1.4 数据分析 广义线性模型用于比较不同建群蜂数量和不同蜂种类间性比和单雌子代数的差异。S曲线模型[ln (y)=β0+β1/x] 被用于拟合建群蜂数量与性比的回归关系。数据统计用SPSS 20完成,柱形图和回归模型由GraphPad Prism 7绘制。
2 结果与分析
2.1 建群蜂种类和数量对子代性比的影响 3种肿腿蜂均为显著的偏雌性比。性比与肿腿蜂种类无显著相关性(χ2=1.76,df=2,P=0.414),但子代性比与建群蜂数量显著相关(χ2=55.25,df=7,P<0.001)。总体上,3种肿腿蜂子代性比随建群蜂数量的增加而增大(Sg: χ2=16.07,df=7,P=0.024; Sp: χ2=42.89,df=7,P<0.001; Sa: χ2=29.02,df=7,P<0.001)(图1)。当建群蜂数量为7头时,松墨天牛肿腿蜂子代性比显著高于管氏肿腿蜂和白蜡吉丁肿腿蜂(χ2=14.98,df=2,P=0.001),在其余相同建群蜂数量条件下3种肿腿蜂子代性比均无显著差异(图1)。
2.2 单雌子代数与性比的关系 3种肿腿蜂每窝平均单雌子代数与蜂种显著相关,管氏肿腿蜂和白蜡吉丁肿腿蜂单雌子代数显著高于松墨天牛肿腿蜂(χ2=382.03,df=2,P<0.001)。随建群蜂数量增加,3种肿腿蜂单雌子代数显著减少(χ2=1 468.65,df=7,P<0.001)(图2)。管氏肿腿蜂建群蜂数量为1头时其单雌所得子代数量显著高于其他处理,当建群蜂数量增加到3头之后单雌子代数趋于稳定(χ2=619.22,df=7,P<0.001)(图2)。白蜡吉丁肿腿蜂和松墨天牛肿腿蜂建群蜂数量为1头时其单雌所得子代数量显著高于其他处理,当建群蜂数量增加到6头之后单雌子代数趋于稳定(Sp: χ2=1 038.71,df=7,P<0.001; Sa: χ2=123.08,df=7,P<0.001)(图2)。拟合性比与单雌子代数量关系发现,3种肿腿蜂子代性比与单雌子代数均呈显著的负相关(图3)。
2.3 肿腿蜂性比实测值与LMC模型预测值差异 3种肿腿蜂子代雄性比虽随建群蜂数量增加呈小幅增加,但LMC模型并不能准确地描述肿腿蜂的真实性比。基于LMC模型(I、II)的性比预测值在建群蜂数量为1头时可达极小值(趋于0),3种肿腿蜂在建群蜂数量为1头时子代性比实测值介于2.00%~5.00%(图4)。未考虑近交发生时(模型I),建群蜂数量为2头时的预测性比为25.00%,且随着建群蜂数量增加性比预测值仍进一步增大,当建群蜂数量为8头时的预测性比为43.75%,但3种肿腿蜂在建群蜂数量介于2~8头之间的实测性比均小于LMC模型I的预测值(图4)。加入近交系数后(模型II),LMC模型的预测值仍极大地偏离了肿腿蜂的实测性比(图4)。依据3种肿腿蜂的实测性比与建群蜂数量关系的模型拟合发现,肿腿蜂性比随建群蜂数量增加表现为先增加后趋于稳定(图4)。
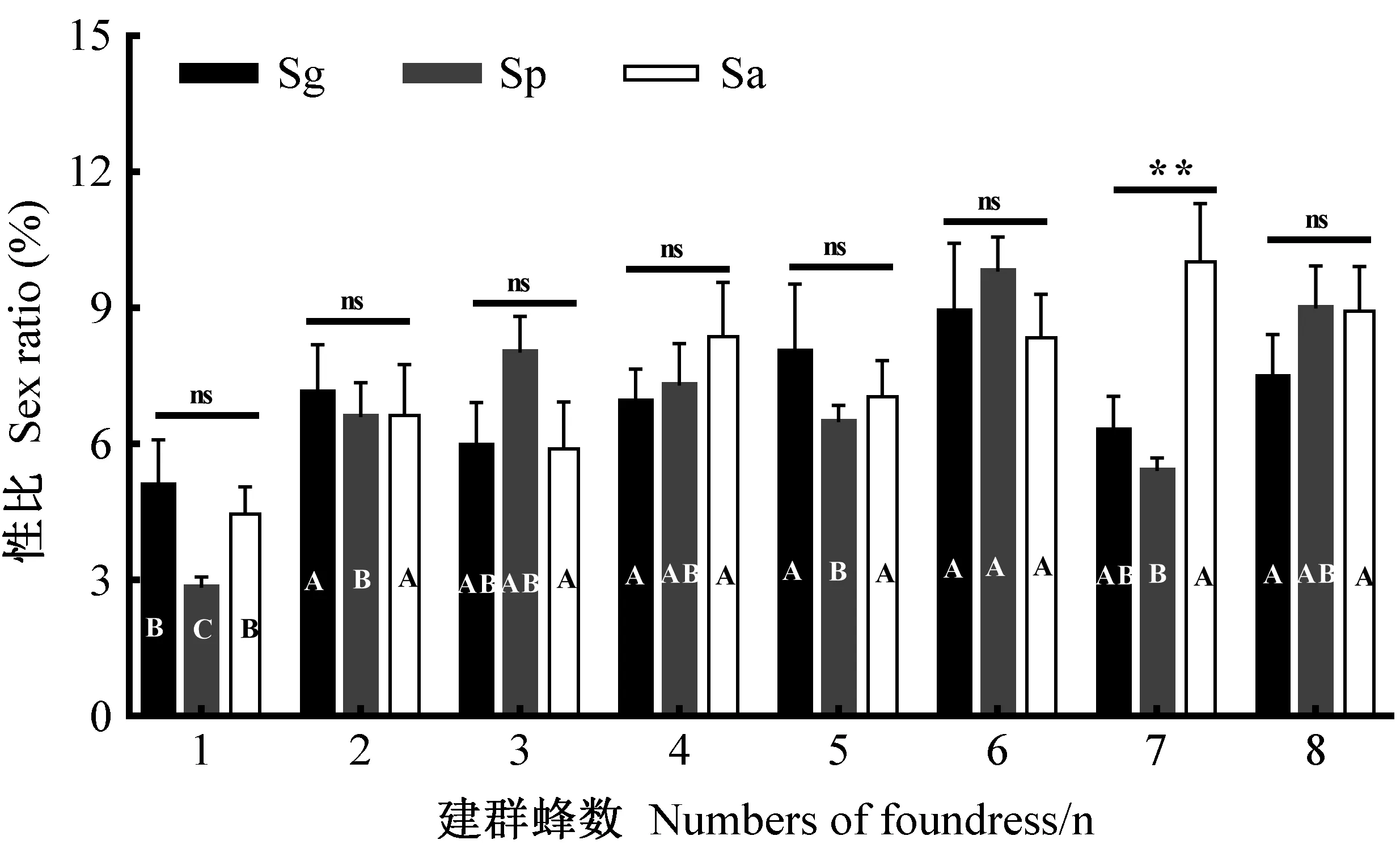
图1 不同建群蜂数量下肿腿蜂的性比Fig. 1 Sex ratio of the bethylid wasps under different foundress densitiesSg:管氏肿腿蜂S. guani; Sp: 白蜡吉丁肿腿蜂S. pupariae; Sa: 松墨天牛肿腿蜂S. alternatusi。同一颜色柱内字母表示同种肿腿蜂在不同建群蜂数量下子代性比差异,不同字母表示处理间显著差异 (α = 0.05), 同一建群蜂数量下3种肿腿蜂性比差异在柱顶标注,ns为无显著差异,**为差异极显著 (α = 0.01); 误差线为标准误。下同 。 Different letters in the same colored bars show significant difference of sex ratios among different foundress densities in a bethylid wasp, while the ‘ns’ and ‘**’ above the bars mean no significance and significant difference of sex ratios of three bethylid wasps under the same foundress density at α = 0.01, respectively; Error bars are presented as SEM. The same below.
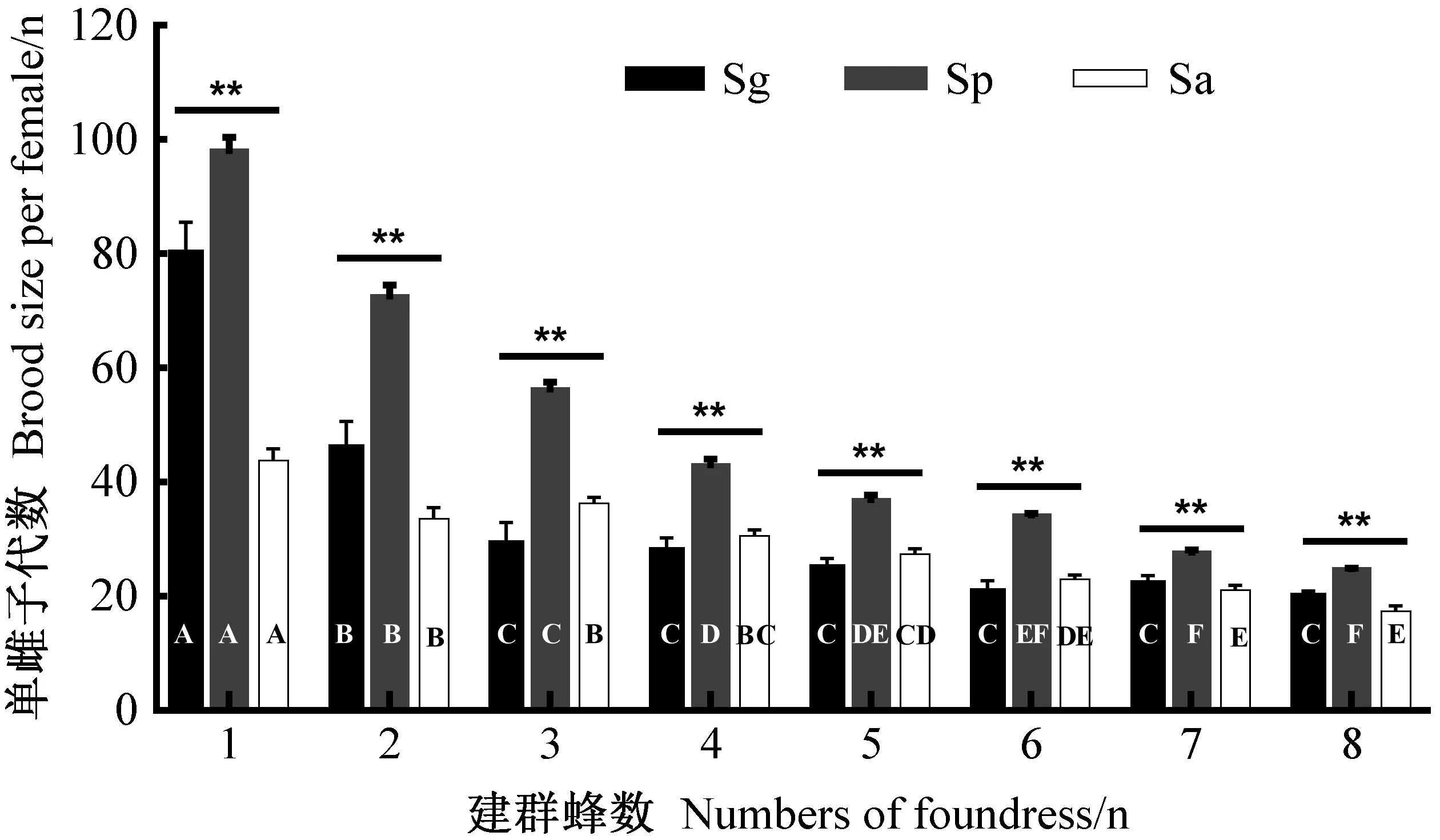
图2 不同建群蜂数量下肿腿蜂单雌子代数Fig. 2 Brood size per female under different foundress densities
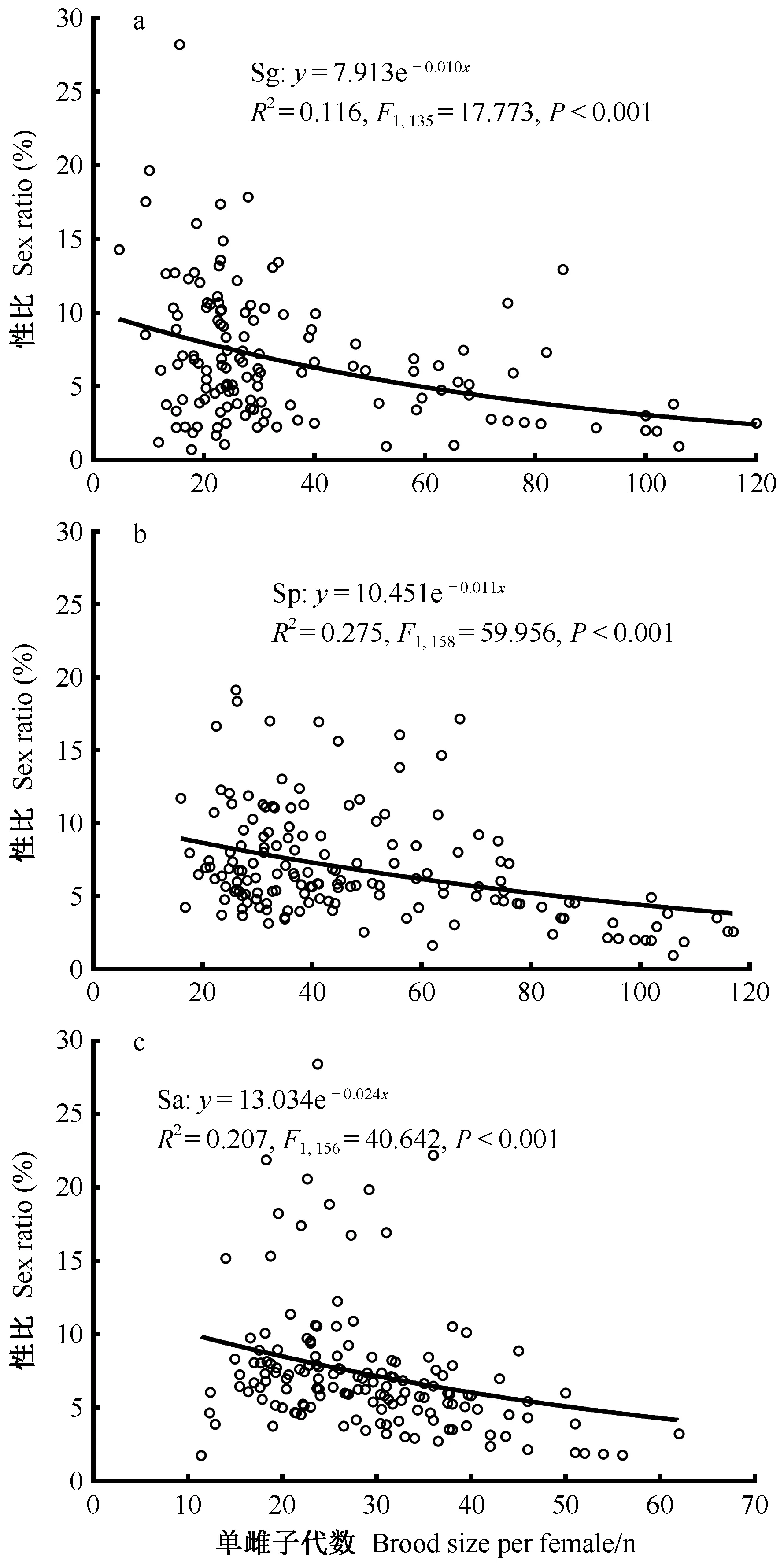
图3 肿腿蜂单雌子代数与性比的关系Fig. 3 The relationships between brood size per bethylid wasp and offspring sex ratio
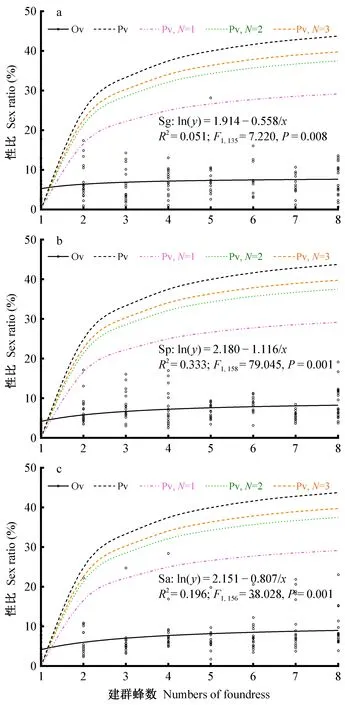
图4 不同建群蜂数量下肿腿蜂性比与基于LMC模型的性比预测值 Fig. 4 Measured and LMC predicated sex ratios of bethylid wasps under different foundress densitiesPv: 基于LMC理论的性比预测模型,其中黑色虚线所示为未考虑近交发生(模型I),彩色虚线所示为考虑近交发生,且分别设定自然种群建群蜂数为1、2和3头(模型II,N=1, 2, 3)。Ov: 性比实测值,以黑色圆圈表示,实线为实测性比与建群蜂数的拟合模型。Pv: Predicted sex ratio based on LMC model, the black dot line indicates the values without inbreeding (model I) and the colored dot lines indicate the vaules with inbreeding along with N=1, 2, 3 (model II); Ov: Observed values of offspring sex ratio, the black circles and full lines indicate the measured sex ratios and regressions on foundress number.
3 讨论
大多数膜翅目昆虫的性别由胚胎倍型决定,即未受精的卵发育成单倍体雄蜂,受精卵发育成二倍体雌蜂(王子龙等, 2019)。在膜翅目昆虫单双倍型的性别决定模式中,互补性性别决定机制(complementary sex determination,CSD)是被普遍认可的,即任意2个性别相关的不同等位基因结合(杂合子)诱导雌性发育,任意2个相同等位基因(纯合子)或单个等位基因结合(半合子)诱导雄性发育(van Wilgenburgetal., 2006)。意大利蜜蜂(Apismellifera)的csd基因首先被发现和克隆,功能性csd仅存在于杂合子中,作为初始信号参与了complementarysexdeterminer(csd)→feminizer(fem)→doublesex(dsx)调控级联,进而诱导雌性发育(Beyeetal., 2003; Hasselmannetal., 2008)。在这一通路中,fem是黑腹果蝇(Drosophilamelanogaster)transformer(tra)基因的直系同源基因,而tra→dsx通路在很多昆虫性别决定中高度保守(Paneetal., 2002; Halletal., 2015; Meccarielloetal., 2019)。有的膜翅目昆虫性别虽然仍是基于tra→dsx通路的调控,但tra上游却不存在csd基因。近期在丽蝇蛹集金小蜂(Nasoniavitripennis)基因组中鉴定到的waspoverrulerofmasculinization(wom)基因,它作为一个全新的上游指示基因激活tra→dsx通路介导雌性发育,并且wom在母本基因组中是沉默的,其仅转录自受精卵中父方提供的基因组(Zouetal., 2020)。csd和wom这些性别决定通路指示基因的相继发现,表明在膜翅目昆虫tra(或fem)的上游可能还进化出了其他功能类似的基因,它们可以使父本能够介导雌性子代的发育。
寄生蜂是膜翅目昆虫中的重要类群。单双倍型的性别决定方式使得寄生蜂可以通过受精作用来调整子代的性比,这一生殖机制决定了母蜂调节子代性比的便捷性。本研究发现,管氏肿腿蜂、白蜡吉丁肿腿蜂和松墨天牛肿腿蜂的性比均是偏雌性,这一特征符合局部配偶竞争(LMC)理论对群居型寄生蜂性比偏雌特征的描述。本研究中当建群蜂为1头时,3种肿腿蜂子代雄性比均在5%左右(Sg=5.07%; Sp=2.83%; Sa=4.46%)。笔者原始数据记录的在1头建群蜂时,3种肿腿蜂子代雄蜂仅有非常少的数量(平均值: Sg=3.53,♀=76.58; Sp=2.8,♀=94.80; Sa=1.85,♀=41.85),表明当1头建群蜂时,仅需很少数量的雄蜂就能与子代雌蜂完成交配。LMC模型预测建群蜂数量为1头时的子代雄性比为0,这实际是代表了雄性数量趋于无限小的可能。因此推测3种肿腿蜂在建群蜂为1头时子代性比的特征是符合LMC性比预测模型的。然而,当建群蜂数量增加时,3种肿腿蜂实测性比却较大地偏离了LMC模型的预测值。3种柄翅小蜂(Gonatocerusfasciatus、G.ashmeadi、G.triguttatus)的性比随建群蜂数量变化的关系就完全符合LMC模型的预测趋势,当建群蜂数量为1头时子代雄性比均低于10%,而建群蜂数量为5头时子代雄性比则超过40%(Irvinetal., 2006)。棱角肿腿蜂(Goniozusnephantidis)性比特征也基本符合LMC模型(Abdietal., 2020),LMC模型还在其他一些寄生蜂中得到了一定程度的验证(Marteletal., 2010; Boultonetal., 2019)。基于本研究的结果,认为LMC模型对硬皮肿腿蜂(Sclerodermusspp.)的性比结构预测精度低。
实验模型与理论模型的差异是源于多方面的。LMC模型对性比的预测是基于种群的第一性比(或称原始性比,primary sex ratio),而本研究观测的试验数据则是种群的第二性比(secondary sex ratio)。幼蜂在发育过程中的死亡会造成两类性比不对等。单寄生蜂在每头寄主上仅产出1粒卵,依据观测产卵时母蜂产卵器的动作并联系卵最终发育成熟时的性别,研究人员总结出一些经验性的依据母蜂产卵行为评价初级性比的方法,在姬蜂(Itoplectismaculator)(Cole, 1981)和螯蜂(Haplogonatopusatratus)(Yamadaetal., 1999)中得到应用。但这些经验性的估测方法不具备广泛的适用性,特别是对于群居型寄生蜂,它们在1头寄主上产多粒卵,产卵行为与卵性别间难以建立联系。一些分子标记手段被用于测定群居型寄生蜂卵的性别(Burton-Chellewetal., 2008; Khidretal., 2013),但一旦这些卵被用于遗传测定,它们势必不能再发育至成熟。所以在这种情况下获得了真实的第一性比(这里指具体某一窝卵),第二性比又仅能通过相同发育条件下其他窝的平均值来估测,其真实值难以获得。对寄生蜂初级性比的准确测定直到今天仍是一个较难的问题。本研究所测得的3种肿腿蜂的性比均是第二性比,源于第一性比和第二性比的差异可能是它们性比实测值与LMC模型理论值差异较大的原因之一。
另外,LMC理论模型在多种较为低等的膜翅目昆虫中得到证实,但在典型的社会性膜翅目昆虫中,其子代性别分配就与LMC理论模型相差甚远。Alexander等(1977)认为蜜蜂性别分配受多种因素影响因而用模型难以预测。社会性膜翅目昆虫母蜂不仅可以调节子代雄性的数量,还可以控制雄性出现的时间。肿腿蜂类寄生蜂是介于非社会性和社会性昆虫之间的类社会性昆虫(伍绍龙等, 2013),从试验结果来看,肿腿蜂这类寄生蜂的性比也不能用LMC理论模型进行预测,说明其比非社会性昆虫要进化。有证据表明,肿腿蜂这类群居型寄生蜂可能是社会性膜翅目昆虫进化的关键类群,其在生物学上表现的诸多现象(例如抚幼)与社会性昆虫相似,值得深入研究。
本研究发现建群蜂数量增加时,3种肿腿蜂子代雄性比均有一定程度的增加。母代对子代性比结构的调节,无一例外均是要实现适合度最大化(Westetal., 2002)。雌雄子代从胚胎发育至成蜂对寄主资源的需求差异促成了母蜂会通过选择产雌或者产雄来实现适合度最大化。一般认为,雄性发育至成蜂所需的寄主资源较雌性更少(Harveyetal., 2002; Weietal., 2014)。本研究发现随着建群蜂数量增加肿腿蜂平均单雌子代数减少。平均单雌子代数减少反映了建群蜂对寄主资源竞争的评估结果。当仅有1头产卵母蜂时,子代幼蜂之间的资源竞争更弱,这时雄性的数量仅用于满足交配即可,母蜂可以更多地产出雌性子代,这样即可在子一代就增加亲代的适合度。当有多头产卵母蜂时,寄主资源受到限制,若继续产出更多的雌性子代,势必会造成一部分雌子代不能发育至成熟。因此母蜂在这种情况下则选择增加雄性后代比例,这些雄性后代可与其他母蜂的雌性后代交配,如此亲代适合度可在子二代实现最大化。因此,当建群蜂数量从1头增加为多头时,肿腿蜂会感知到周围其他肿腿蜂也在产卵,判断寄主资源可能出现竞争,进而亲代产出更多的雄子代来保证自身繁殖适合度最大化。
本研究虽然明确了母蜂是通过控制产出的卵是否受精来调节子代的性比结构,但在这一过程中具体的行为模式仍然不明晰。对金小蜂(Pachycrepoideusvindemiae)的产卵行为和性别分配研究表明,当其卵子通过输卵管时,输卵管有25%的时间扭转方向,使得卵的受精孔遭到遮敝,这样卵便不能受精,从而导致了其子代偏雌性比降低(van Alphenetal., 1982)。另外,控制某一性别被产出的时间也可能是母蜂实现性比调节的途径之一。前期研究证实松墨天牛肿腿蜂在一次寄生产卵过程中会偏向于优先产出未受精的卵(即雄性子代)(唐艳龙等, 2020b)。推测在多建群蜂时,肿腿蜂面临着单雌产卵量减少,而雄子代又被优先产出(即总有雄蜂被产出)的情况,故其子代雄性比增加,实现性比调节。由于雄性先产而导致在建群蜂数量增加时子代雄性比同步增加在一些传粉榕小蜂中已经得到了证实(Chungetal., 2019; Greeffetal., 2020; Zhangetal., 2020)。
4 结论
1) 管氏肿腿蜂、白蜡吉丁肿腿蜂和松墨天牛肿腿蜂这3种肿腿蜂均为显著的偏雌性比。
2) 性比与肿腿蜂种类无显著相关性,但子代性比与建群蜂数量显著相关。3种肿腿蜂子代性比与单雌子代数均呈显著的负相关。
3) 3种肿腿蜂子代性比虽随建群蜂数量增加呈小幅增加,但与LMC模型的预测值差异较大。高度偏雌的子代性比不会因建群蜂数量的增加而被均衡化,LMC模型并不适用于对肿腿蜂这类亚社会性寄生蜂性比结构的预测。
4) 由于偏雌性比高度稳定,肿腿蜂繁殖中不用担心增加接蜂数量会导致过多的于生物防治不利的雄蜂被产出。
