滇西南山地农耕区小兽多样性空间分布格局及与寄生蚤关系*
2022-10-09琚俊科龚正达高子厚连宏宇权寿瑛段兴德冯锡光张丽云
琚俊科 李 栋 龚正达 高子厚 连宏宇 权寿瑛 段兴德 冯锡光 洪 梅 张丽云
(1.大理大学公共卫生学院, 云南大理 671000;2.云南省地方病防治所, 云南大理 671000;3.云南省寄生虫病防治所, 云南普洱 665099;4.安阳市第五人民医院,河南安阳 455000;5.北大医疗潞安医院, 山西襄垣 046204)
山地生态学是一门研究在山区这一特定环境中,不同生命层次的生态现象和过程及其相互作用的科学领域(方精云等,2004)。既往在云南横断山区的研究发现,小兽物种多样性以中山地带森林植被条件较好地带为高,而气候环境条件恶劣、植被条件较差的山顶部和人们生产活动频繁的山脚带山地农耕区小兽物种多样性和物种丰富度都相对较低,说明气候条件与人们的生产活动对物种多样性有重要影响 (龚正达等,2001a)。
山地农耕区是一种常见但特殊的生态环境类型,为了生存需要,人类对原自然环境中的森林进行砍伐、开垦和长期耕耘,导致当地部分物种的退出,或其他物种入侵定居,与周围山地自然环境和动植物区系有一定联系但又有很大差别,长期在人为影响和干扰下形成的一类生物群落即人工群落(连宏宇,2010;琚骏科等,2021)。云南西部山地广泛存在家鼠型鼠疫和野鼠型鼠疫两类型疫源地(俞东征,2009; 方喜业等,2012;龚正达等,2018),小兽及其寄生蚤是鼠疫的主要宿主和媒介,他们对鼠疫疫源地的长期保存、疾病的传播和流行起着重要的作用。因此,滇西南家鼠鼠疫疫源地山地农耕区小兽物种多样性空间分布特征与其寄生蚤类多样性之间的关系如何,是否同步或存在差异,以及导致它们之间差异的主要因素都是值得进一步研究和探讨的问题(琚骏科等,2021)。本研究以滇西南家鼠鼠疫疫源地(北纬25°50′以南)4州(市)8县(市)山地农耕区为调查取样的空间范围,以小兽及其体表寄生蚤为研究对象,在大量现场调查、数据整理、统计学处理和分析的基础上,就家鼠鼠疫疫源地农耕区小兽物种多样性空间分布格局、特征及其与寄生蚤多样性关系进行分析和研究,并对云南家鼠型鼠疫、野鼠型鼠疫与喜马拉雅旱獭鼠疫疫源地之间关系等进行初步探讨。
1 材料与方法
1.1 调查方法
1.1.1调查研究的对象和空间 据云南省农业区划对云南山区农用耕地划分为3种类型:高寒层(海拔2500 m以上)气候严寒、霜期较长,≥10 ℃的积温小于3 000 ℃,多一年一熟的农耕区;中暖层(海拔15 00~2 500 m),亚热带或暖温带等气候类型,≥10 ℃的活动积温从3 000~6 000 ℃,多一年二熟的农耕区;低热层(海拔1 500 m以下),属于南亚热带和热带,≥10 ℃ 的活动积温6 000~8 500 ℃左右,种植水稻、甘蔗、热带经济作物和水果的农耕区(连宏宇,2010)。
以农耕区小型兽类(简称“小兽”)及其寄生蚤为研究对象,在云南西南部的临沧市(耿马县、临沧市区)、德宏州(陇川县、盈江和梁河县)、保山市(龙陵县、腾冲市)和大理州(云龙县)共8县(市)为调查取样的空间范围(约北纬23°00′N~26°00′N,海拔高度800~3 000 m),各县市调查样区具体的地理位置和分布情况见图1。依据各县市山地农耕区的地理环境、海拔高度和不同温层农耕地类型及条件,调查区域可划分为4个垂直梯度带,以A~D表示,分别代表海拔800~1 500、1 500~2 000、 2 000~2 500、2 500 m及以上地区的取样区间。课题组于1998—2000年对耿马县、临沧市、陇川县、盈江县和龙陵县,2008—2011年对梁河县、腾冲市、云龙县8县(市)共36乡镇(调查点)农耕区小兽及其寄生蚤类进行了现场调查取样。
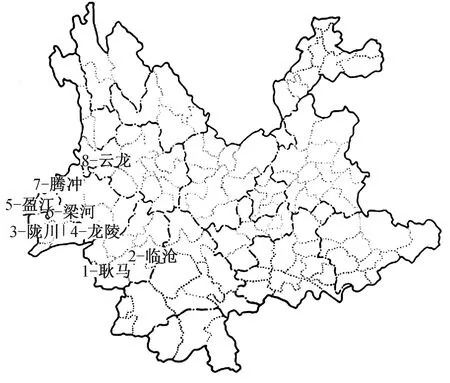
图1 云南西南部8县(市)山地农耕区小兽及寄生蚤调查区域的地理位置及分布
1.1.2小兽及其寄生蚤的调查取样 在旱季(雨季前、后期)使用大号鼠夹(夹夜法)晚放早收进行小兽和寄生蚤的调查取样。在每县(市)样区内各选4~6个不同海拔高度乡镇的农耕区为调查点,每个调查点捕鼠2晚,每晚置夹200个;此外,在农耕区居民点室内采用电猫(都乐牌电子灭鼠器,柳州市家用电器灭鼠器厂制造)进行辅助捕鼠取样。现场所获鼠体分别袋装,带回当地疾控中心实验室或乡镇卫生院工作室进行分类、记数,并分别采集其体外寄生蚤放入75%酒精内标记和保存,后期进行标本制作和分类计数。
1.2 蚤类的数据来源
本文中寄生蚤的物种组成、分布情况、物种多样性等数据源自本课题组近期发表的文章(琚骏科等,2021)。
1.3 统计与分析方法
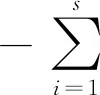
1.3.2聚类分析 以各物种在不同样区或梯度带(样区)的分布及其相对多度(构成比)情况,使用数值系数定性,运用系统聚类法(组平均法)进行聚类和分析(赵志模等,1990)。
1.3.3相关分析 应用SPSS 16.0软件进行Pearson线性相关分析。
2 结果
2.1 滇西南8县(市)山地农耕区小兽物种的组成及其多度分布
通过对耿马、临沧、陇川、龙陵、盈江、梁河、腾冲、云龙8县(市)共36乡镇(调查点)农耕区的调查,共获小兽7科19属35种 2 890 只,小兽的组成和分布情况按各县(市)样区归类形成表1,从所获小兽体表采获寄生蚤类6科16属27种(亚种)共2 840只。
由于地理和气候环境条件上的差异,各县市小兽物种和数量的组成各不相同,其中,耿马8种633只,临沧24种210只,陇川11种412只,龙陵12种595只,盈江14种312只,梁河18种191只,腾冲20种329只,云龙20种208只。各县(市)小兽的物种组成及其多度的分布情况详见表1。
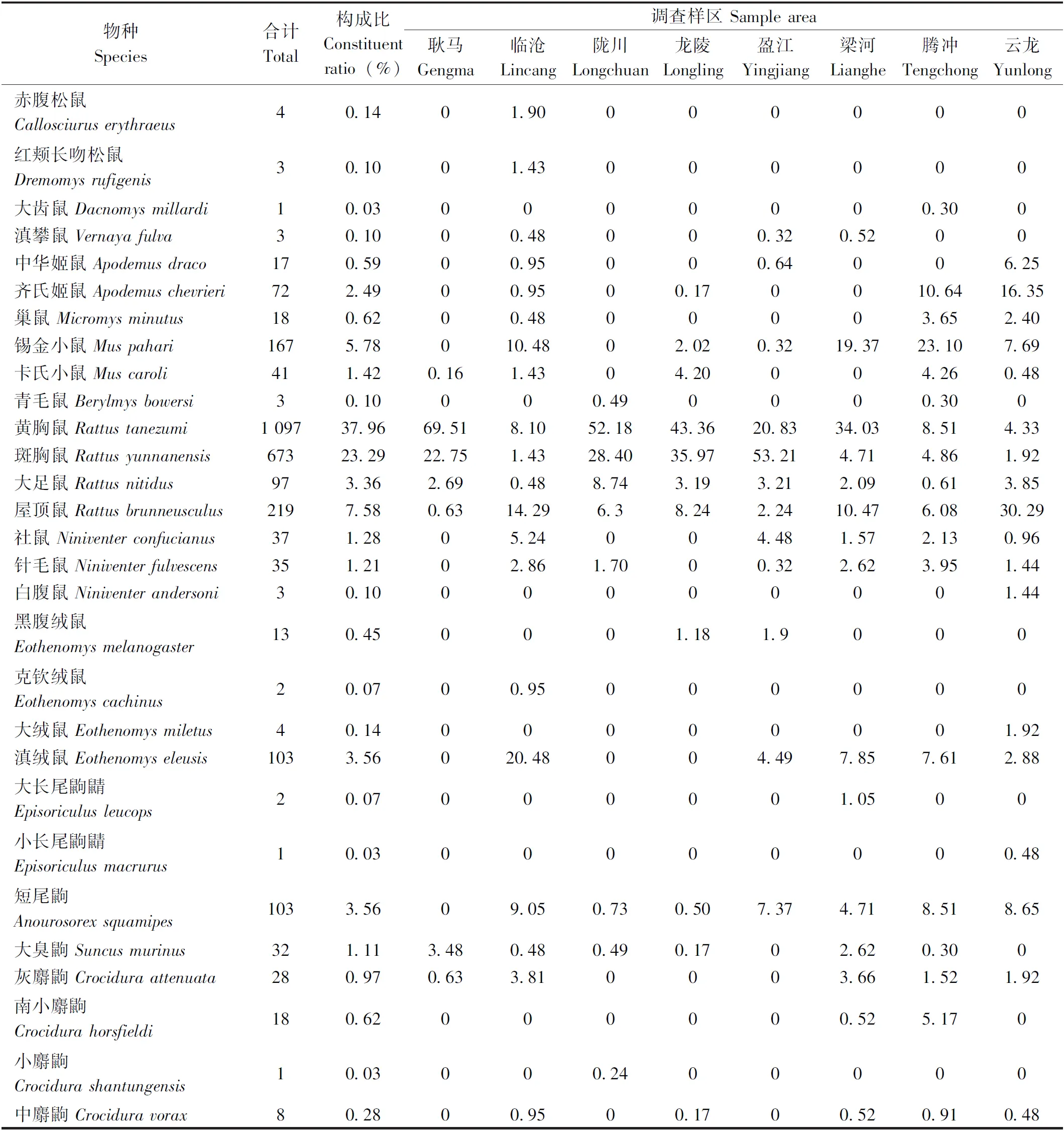
表1 滇西南8县(市)山地农耕区小兽物种的组成及多度分布
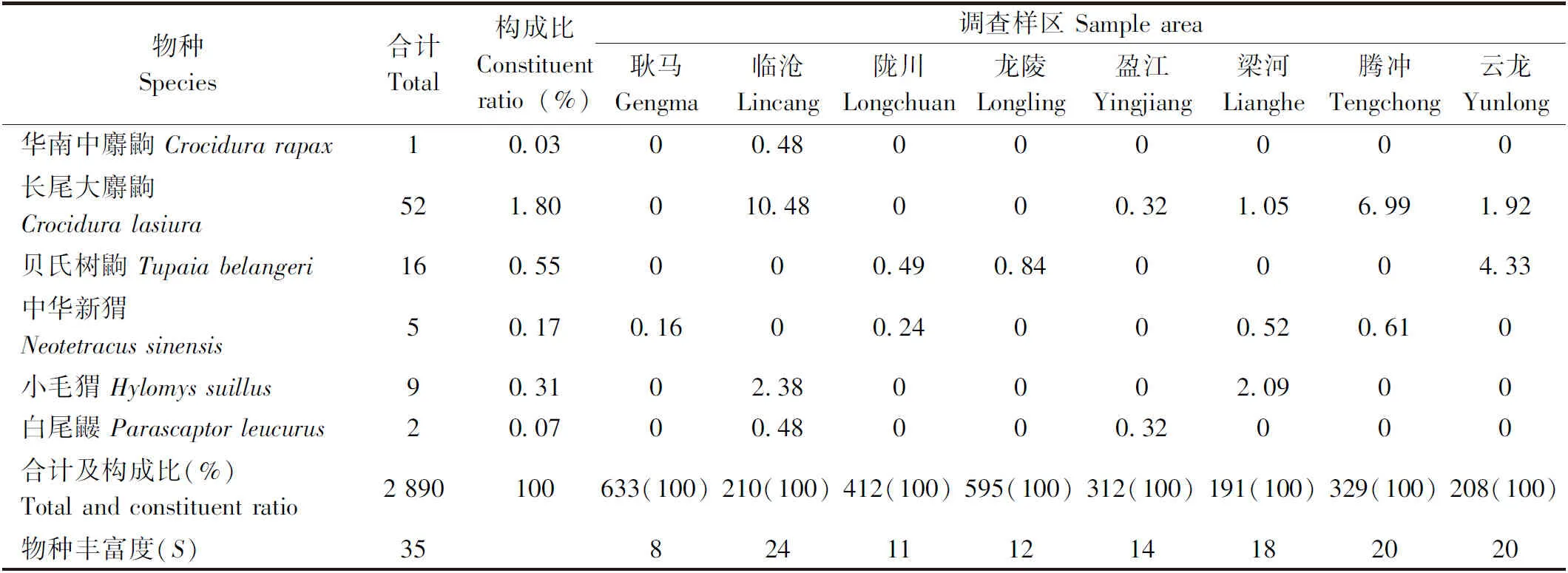
表1(续) Tab.l Continued
从表1中看出:8县(市)山地农耕区所获27种2 890只小兽中,不同物种小兽数量的构成比例明显不同,其中,黄胸鼠数量最多(占37.96%),其次是斑胸鼠(23.29%),该两种鼠类为当地小兽数量上的优势种;屋顶鼠 (7.58%)和锡金小鼠(5.78%)两种鼠类数量相对较多,为当地的常见种;而其余小兽的数量都相对较少。
由于调查区各县(市)地理位置和气候环境条件的差异,各地小兽的物种丰富度明显不同,其中,丰富度最高为临沧(24种),其次腾冲和云龙(各20种),最低为耿马(仅8种)。此外,各县(市)样区小兽的优势种及其多度也不尽相同,其中黄胸鼠是耿马(69.51%)、陇川(52.18%)、龙陵(43.36%)和梁河(34.03%)的优势种;而斑胸鼠(53.21%)、滇绒鼠(20.48%)、锡金小鼠(23.10%)、屋顶鼠(30.29%)分别为盈江、临沧、腾冲和云龙各县市数量上的优势种; 其余物种的多度相对较少或很少。
2.2 滇西南8县(市)农耕区小兽物种多样性的空间分布格局
2.2.18县(市)农耕区小兽物种丰富度(γ-多样性)随海拔梯度的变化趋势 受地理位置、气候环境条件和农耕地类型的影响,小兽物种丰富度沿海拔高度或不同气温层农耕区的分布情况也明显不同。其中,地理位置偏北,海拔偏高的腾冲和云龙两县小型兽类物种丰富度随海拔的升高呈先升高后降低的分布格局,峰值在1 500~2 500 m; 而地理位置偏南,海拔偏低的耿马、临沧、陇川、龙陵、盈江和梁河小兽物种丰富度随海拔的升高,均呈逐步降低的分布格局(图2)。在整个调查区域内,小兽物种丰富度随着海拔梯度的升高或不同的农业气温层(低热层→中暖层→高寒层)气候环境的改变,总体的变化趋势呈现了随海拔升高而逐步降低的分布特征(图2,表2)。
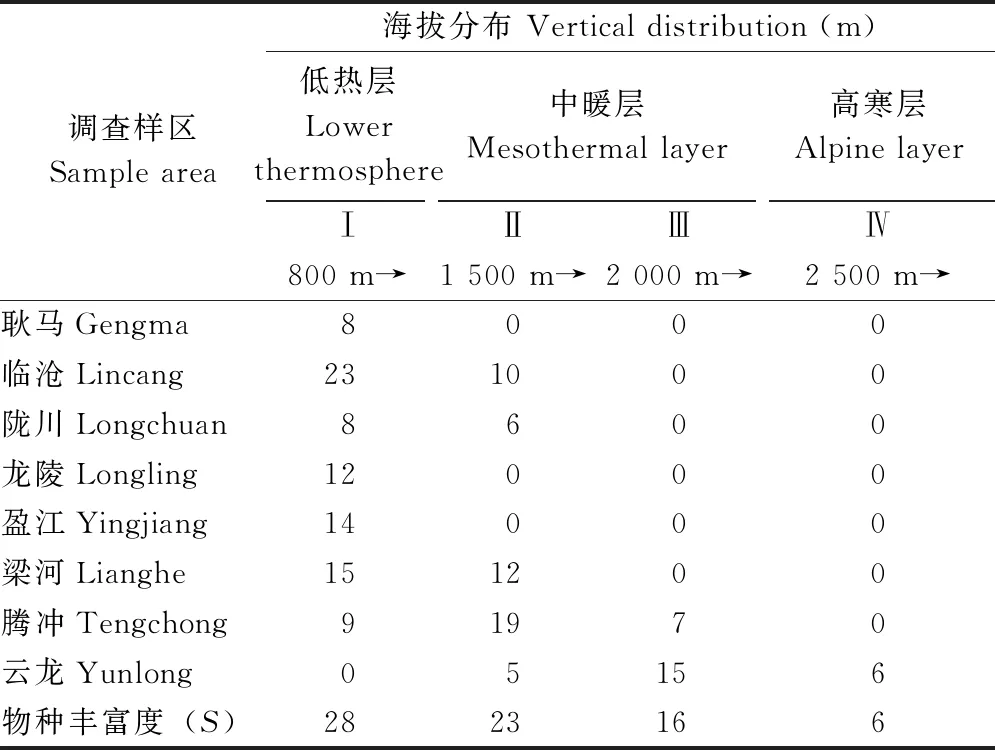
表2 滇西南8县(市)不同气温层农耕区小兽物种丰富度的空间分布
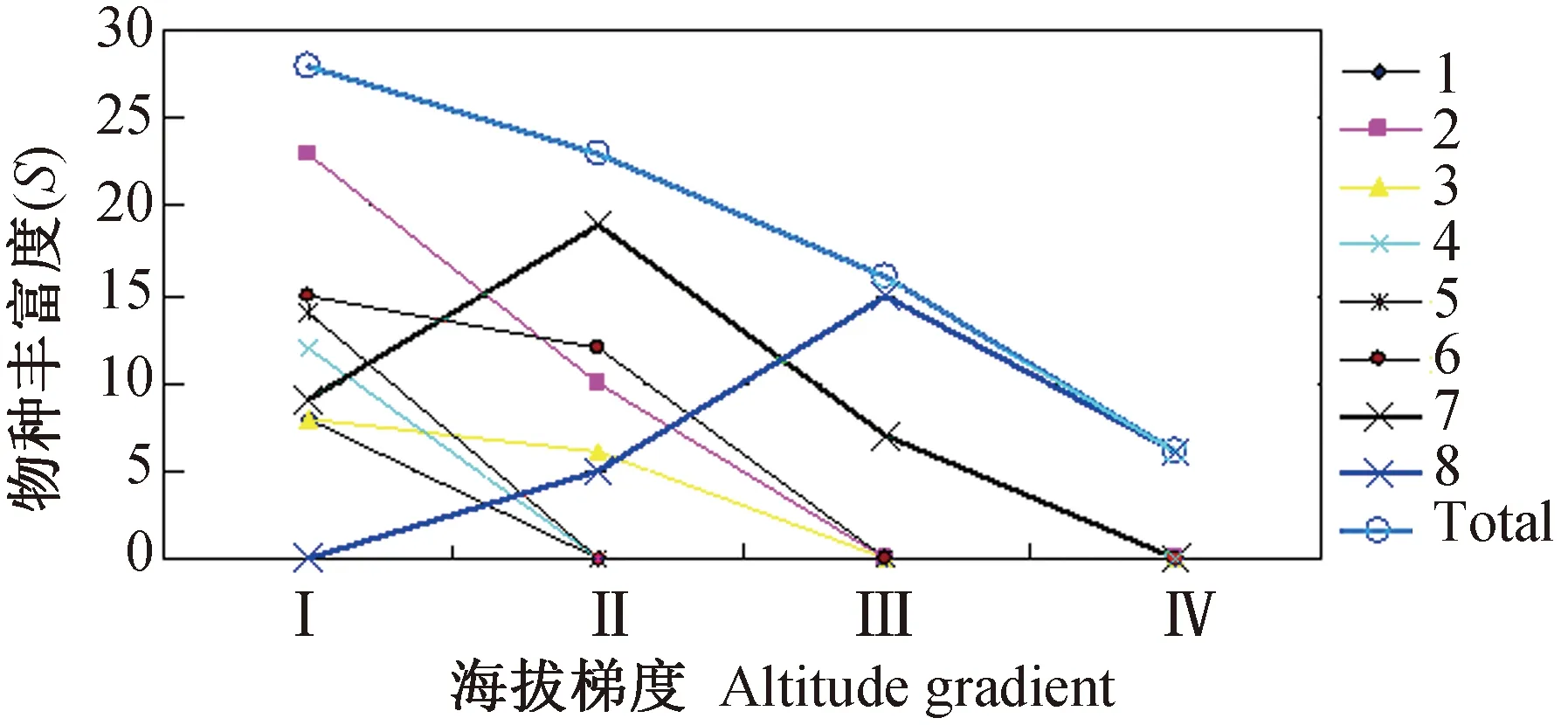
图2 滇西南8县(市)农耕区小兽物种丰富度(γ-多样性)随海拔梯度的变化趋势
2.2.2滇西南农耕区小兽β多样性随海拔梯度的变化趋势 农耕区小型兽类的Cody指数沿海拔梯度的增加呈现逐步升高的趋势(图3),最高点位于海拔C~D带(2 000~2 500 m)。而Sorenson指数随海拔增高呈逐步降低的分布格局,最高点位于A~B带(800~1 500 m)。显示了在高寒层(海拔2 500 m以上)暖温带气候类型农耕区小兽群落、物种和区系的组成发生了较大变化。
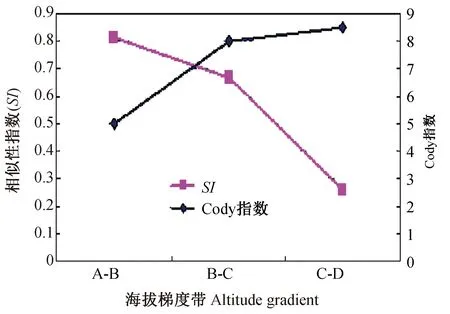
图3 滇西南农耕区小兽β多样性(Cody指数)随海拔梯度的变化趋势
2.2.38县(市)山地农耕区小兽的物种多样性(α多样性)及其与生态优势度的关系 根据8县(市)农耕区小兽物种组成及其物种多度的分布情况(表1),分别测定和统计出各样区小兽的物种多样性指数、物种丰富度、均匀度、生态优势度和优势种多度各项指标(表3)。各县市小兽物种多样性指数、物种丰富度、均匀度、生态优势度和优势种多度各不相同(表3),临沧、腾冲和云龙小兽的物种丰富度(分别为24、20和20种)、物种多样性指数(分别为2.5530、2.5345和2.3770)和均匀度指数(分别为0.8033、0.8460和0.7935)都相对较高,而生态优势度(分别为0.1061、0.1041和0.1445)和优势种多度(分别为20.48、23.10和30.29)都相对较低;与此相反,耿马县的物种丰富度(8种)、物种多样性指数(0.8879)和均匀度指数(0.4270)最低,而生态优势度(0.5369)和优势种的多度(69.51)最高。
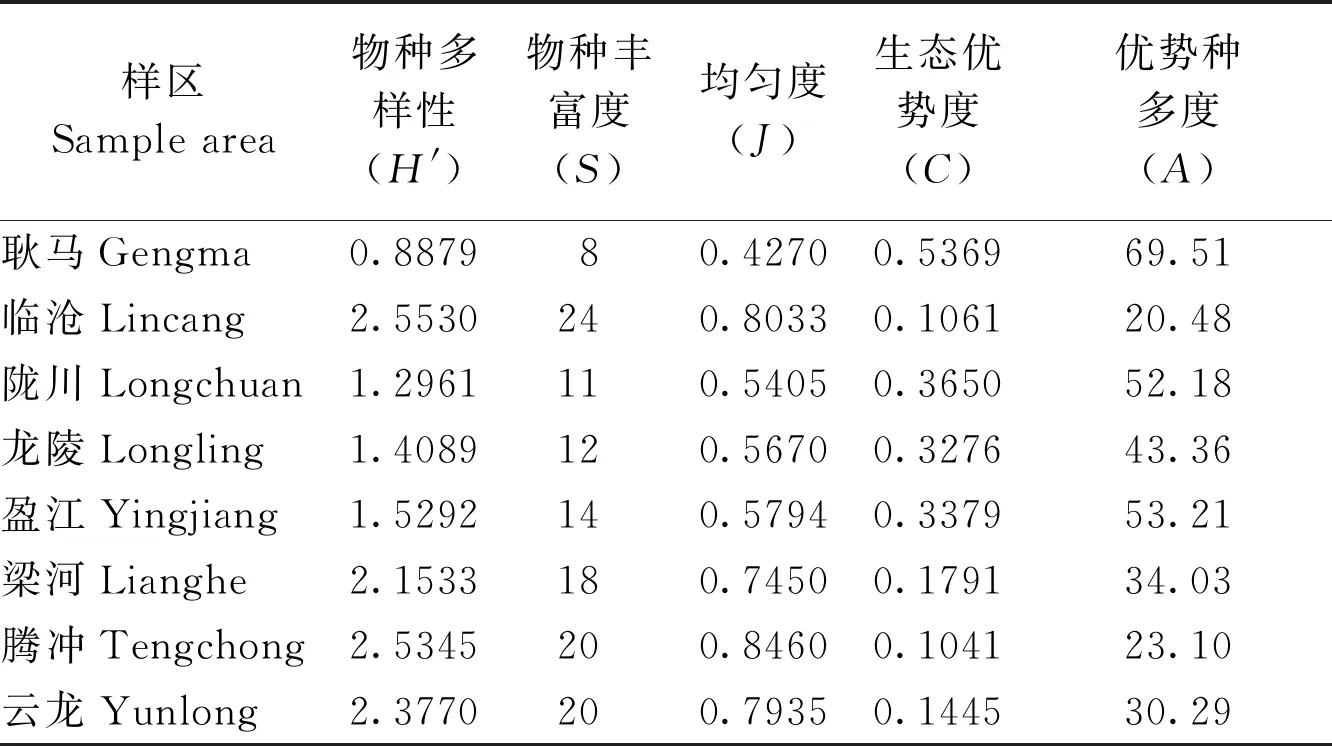
表3 滇西南8县(市)山地农耕区小兽群落生物学指标的测定
8县(市)小兽物种多样性与物种丰富度、均匀度和生态优势度之间的空间分布状况及相关分析显示(图4,5):小兽物种多样性与物种丰富度之间呈极显著正相关(r=0.979,P<0.01),而物种多样性与生态优势度之间呈极显著负相关(r=-0.985,P<0.01)。上述情况说明,8县(市)农耕区小兽的物种丰富度、生态优势度和优势种多度与物种多样性关系密切。

图4 滇西南 8县(市)农耕区小兽物种多样性与物种丰富度的空间分布及其关系

图5 滇西南8县(市)农耕区小兽物种多样性、均匀度和生态优势度的空间分布及其关系
2.2.4小兽物种多样性与优势种多度之间的关系 由于地理和气候环境条件的差异,8县(市)农耕区小兽优势种及其多度不尽相同,其中黄胸鼠是耿马、陇川、龙陵和梁河的优势种(其多度分别为69.51、52.18、43.36和34.03);而斑胸鼠、滇绒鼠、锡金小鼠、屋顶鼠分别为盈江、临沧、腾冲和云龙各地的优势种(各物种多度依序为:53.21、20.48、23.10和30.29)(表1)。根据表3数据,将上述各县(市)农耕区小兽物种多样性指数与优势种多度的分布状况及其关系绘制成图6。

图6 滇西南8县(市)各样区小兽物种多样性与优势种多度之间的关系
从图6 结果中可以看出,在8县(市)各样区内优势种多度高时,则物种多样性都较低,反之则高。各县(市)小兽物种多样性与优势种多度之间呈极显著负相关(r= -0.966,P<0.01)。上述结果说明,滇西南农耕区小兽优势种的组成及其物种多度的空间分布格局主导了该区域农耕区小兽物种多样性(α多样性)的变化趋势。
2.3 8县(市)山地农耕区小型兽类沿海拔梯度分布的聚类分析
根据8县(市)不同海拔高度农耕区所获35种小兽的分布情况,应用数值系数进行了聚类分析,结果发现: 农耕区小兽沿海拔梯度的分布可分为3个主要类型,低海拔、中海拔和高海拔区域(图7),他们沿海拔梯度的垂直分布特征与当地不同气温层农耕区的构成相一致,反映了海拔高度、气候环境条件和耕作区类型对小兽物种、群落的组成及其空间分布格局具有重要影响。

图7 滇西南8县(市)山地农耕区小兽沿海拔梯度分布的聚类分析图
2.4 滇西南8县(市)山地农耕区小兽与寄生蚤多样性的特征及其关系
从8县(市)农耕区所获小兽7科19属35种,小兽体表采获寄生蚤6科16属27种(亚种)。两者科、属和种不同阶元丰富度相比较,小兽科、属和种的丰富度均较寄生蚤类的丰富度高一些,两者之间的差距随着分类阶元的变小而呈现逐步增大的趋势。由于地理和气候环境条件和农耕区类型不同,在所获27种(亚种)2 840只蚤类中,各地蚤类物种及其数量的组成情况亦不相同,其中,耿马4种308只,临沧12种511只,陇川9种372只,龙陵7种578只,盈江8种679只,梁河10种112只,腾冲13种85只,云龙11种195只(琚骏科等,2021)。
2.4.1不同海拔梯度及温层农耕地的小兽与寄生蚤物种丰富度(γ-多样性)的变化趋势 小兽物种丰富度随海拔升高总的趋势呈逐步降低的分布格局,寄生蚤物种丰富度随海拔升高的总趋势呈先增高后降低的分布格局,两者的最大峰值不同:小兽在800~1 500 m间,而蚤类在 1 500~2 000 m间(表4, 图8),但两者物种沿海拔的增高和气候环境的变化,物种丰富度总体呈现为逐步下降的趋势(图8)。
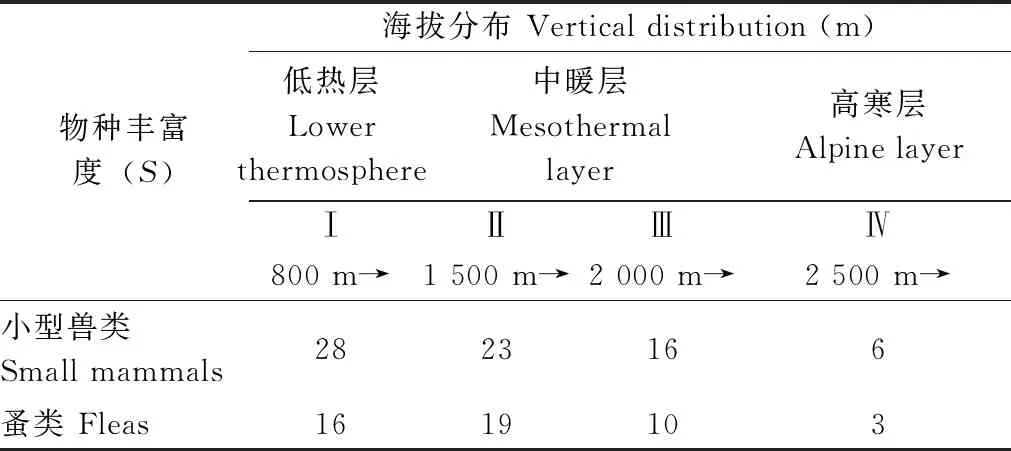
表4 小兽及其寄生蚤物种丰富度(γ-多样性)随不同海拔梯度和温层农耕地的变化

图8 小兽及其寄生蚤类物种丰富度沿海拔梯度的变化趋势
2.4.28县(市)山地农耕区小兽寄生蚤群落生物学指标的测定 通过对8县(市)农耕区小兽寄生蚤类的物种多样性、物种丰富度、均匀度、生态优势度进行测定和统计,结果见表5。从表5看出,在8县(市)调查区中以腾冲市蚤类物种多样性最高(1.9178),其次为梁河县(1.6425)和云龙县(1.6391),而最低为耿马县(0.2753)。
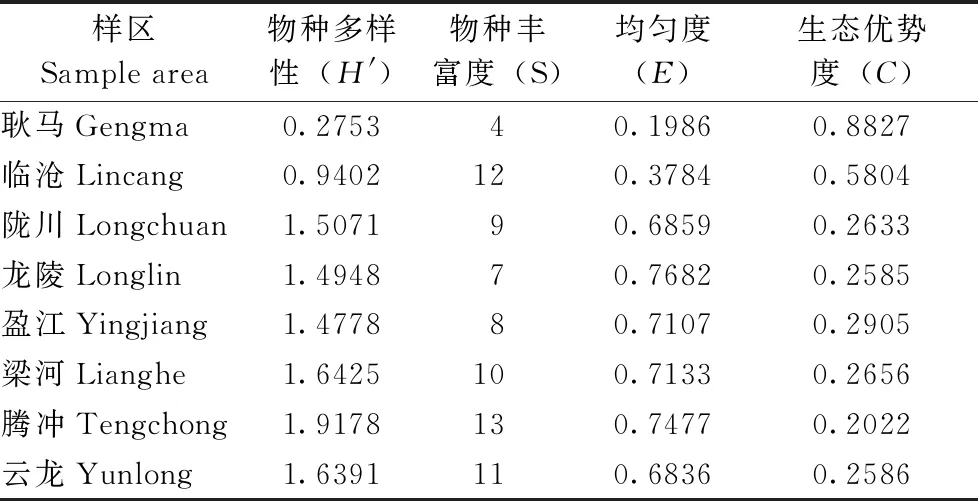
表5 8县(市)山地农耕区小兽寄生蚤群落生物学指标的测定(琚骏科等,2021)
2.4.38县(市)农耕区小兽与寄生蚤类物种多样性之间的关系及分析 对8县(市)农耕区小兽(表3)与寄生蚤(表5)的物种多样性、丰富度、均匀度和生态优势度之间关系进行了相关分析和比较,结果发现:小兽物种多样性指数与寄生蚤丰富度之间呈极显著正相关(r= 0.937,P<0.01)(图9);小兽物种丰富度与寄生蚤丰富度之间呈极显著正相关(r= 0.899,P<0.01)(图10);小兽均匀度与寄生蚤丰富度之间呈极显著正相关)(r= 0.946,P<0.01)(图11);小兽生态优势度与寄生蚤丰富度之间呈极显著负相关(r= -0.954,P<0.01)(图12);小兽与寄生蚤的物种多样性(r= 0.51,P>0.05)(图13)和生态优势度之间不甚相关(r= 0.52,P>0.05)(图14)。
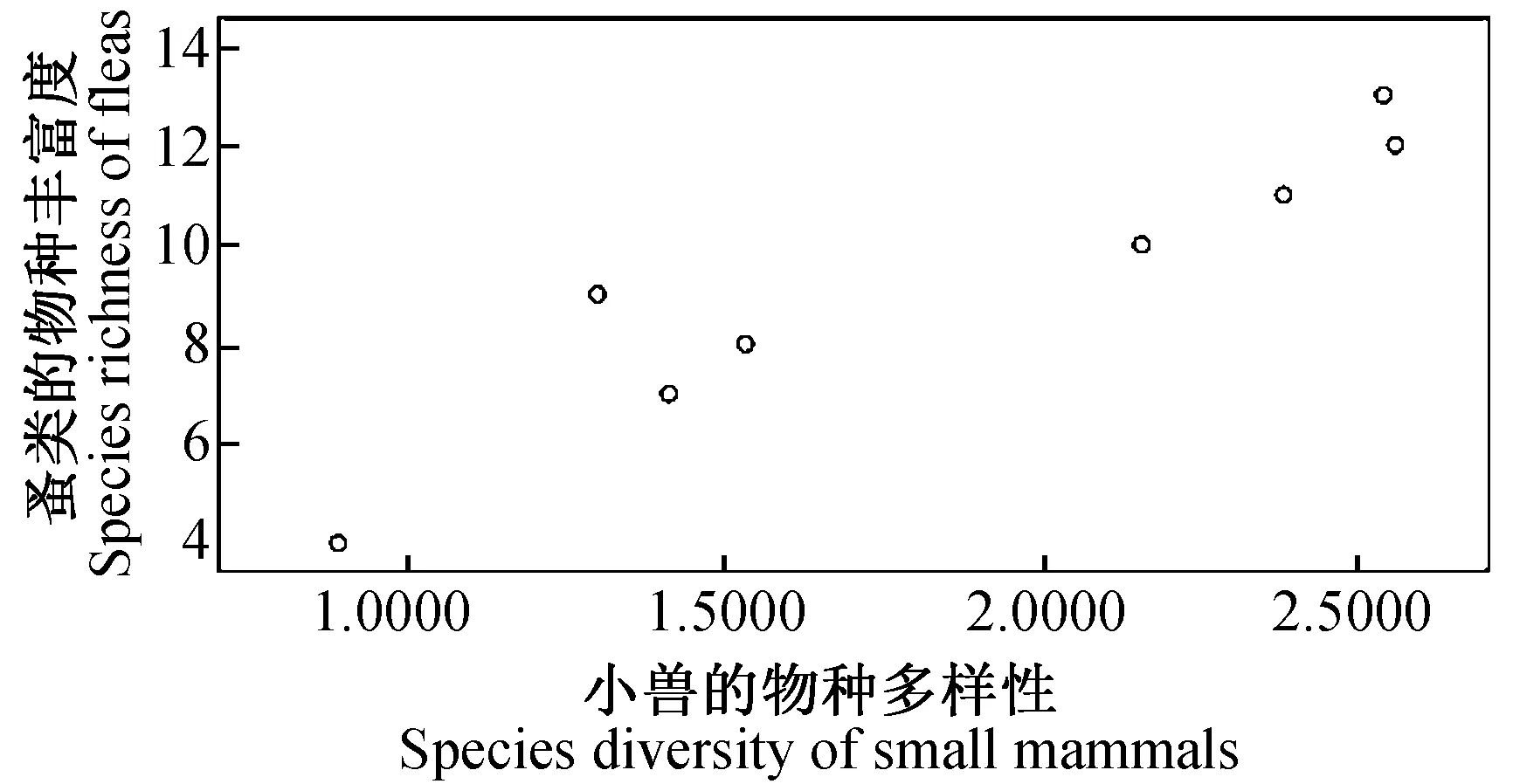
图9 小兽多样性指数与蚤类物种丰富度的关系
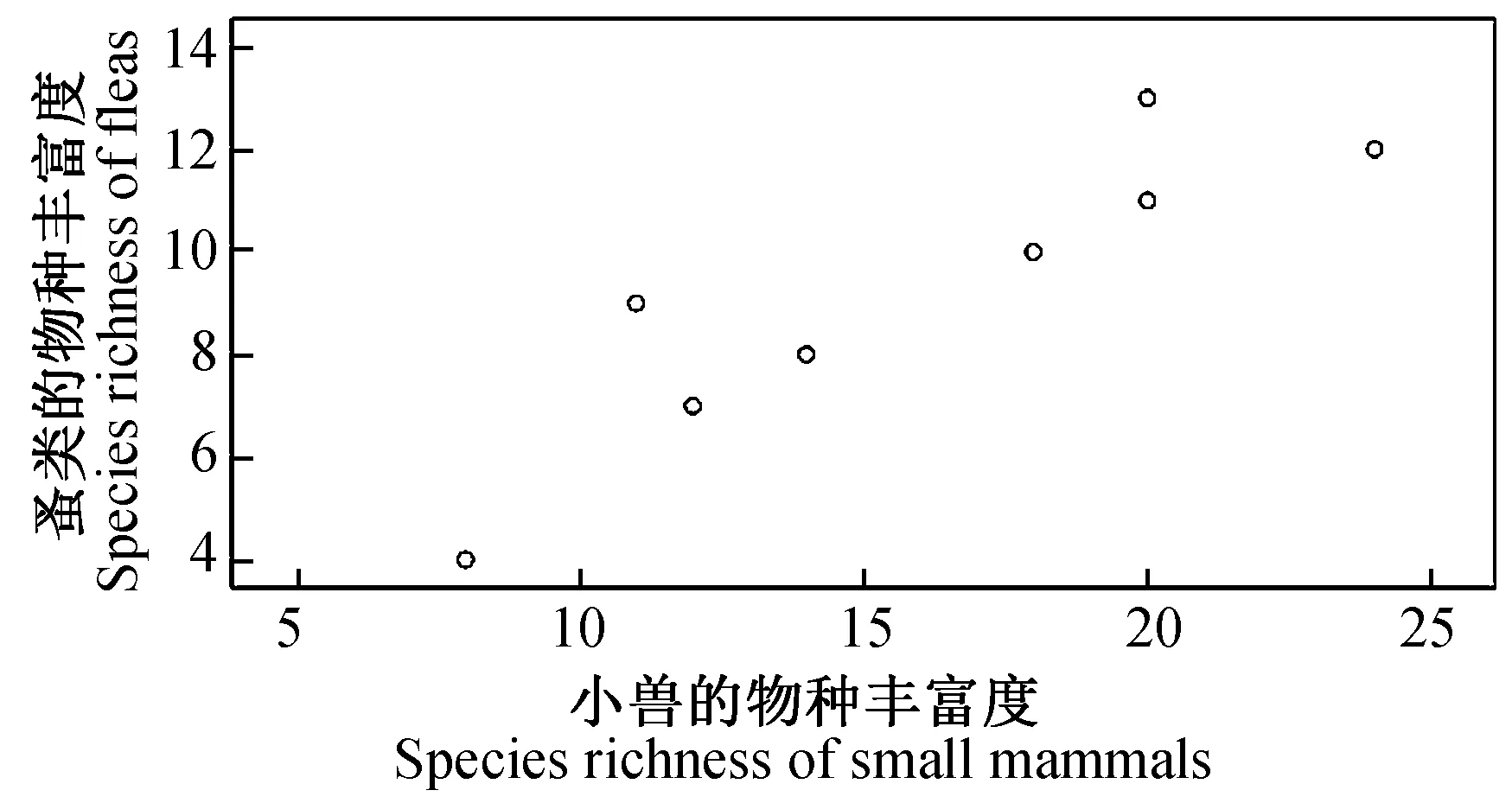
图10 小兽与寄生蚤类物种丰富度之间的关系
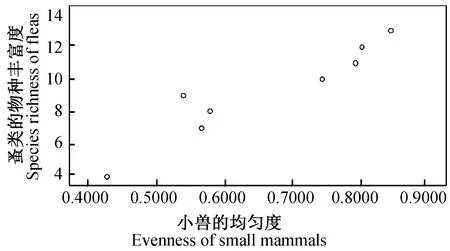
图11 小兽均匀度与蚤类物种丰富度之间的关系
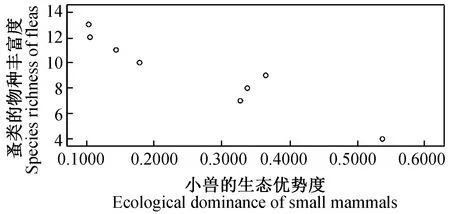
图12 小兽生态优势度与蚤类物种丰富度之间的关系
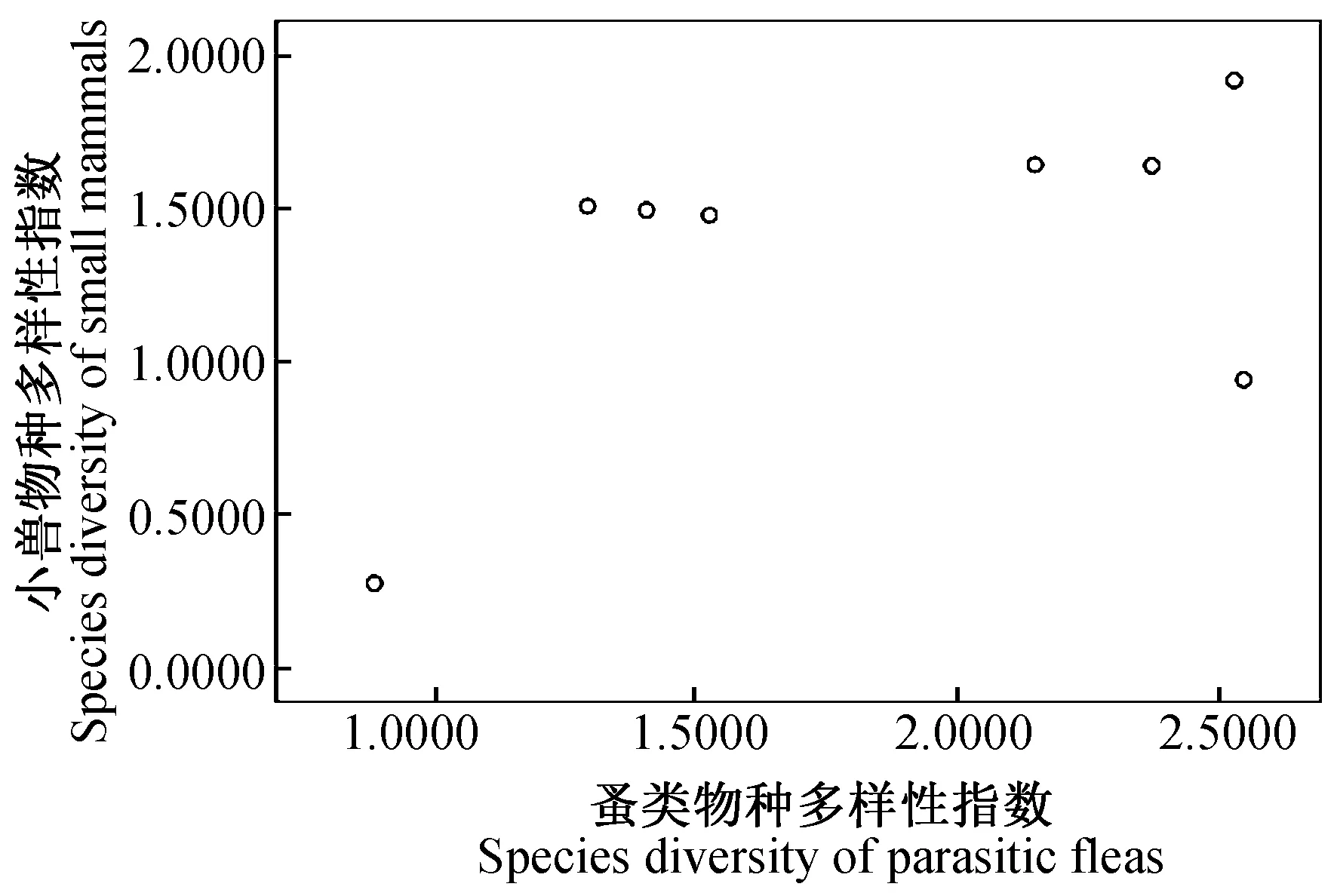
图13 小兽与蚤类物种多样性之间的关系
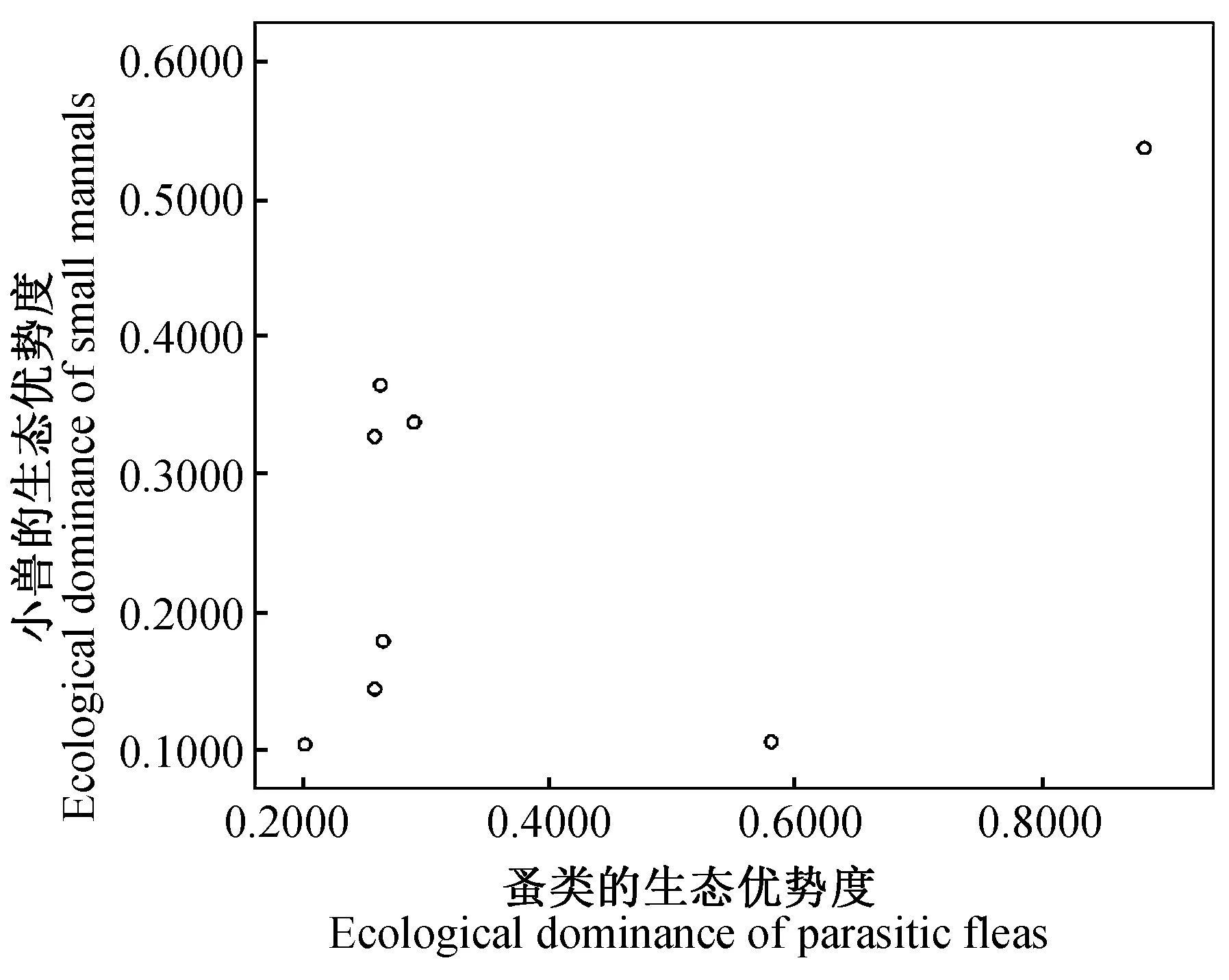
图14 小兽与蚤类生态优势度之间的关系
综上所述,该区域小兽物种多样性、物种丰富度、均匀度与寄生蚤丰富度之间的关系颇为密切,同时也说明了当地小兽与寄生蚤类的物种及其多样性之间存在的协同进化与共存的关系。但是,小兽与寄生蚤的物种多样性之间(r= 0.51,P>0.05)和生态优势度之间(r= 0.52,P>0.05)的关系并不很密切,因此也反映出两者物种多样性和优势种多度的空间分布格局及其特征之间存在一定的差异。
3 讨论
3.1 山地小兽及其蚤类物种多样性空间分布特征及其关系
云南西部横断山区由于存在家鼠型鼠疫和野鼠型鼠疫两类型自然疫源地(俞东征,2009; 方喜业等,2012;龚正达等,2018),而小兽及其寄生蚤是鼠疫的主要宿主和媒介,它们对鼠疫疫源地的长期保存、疾病的传播和流行起着重要的作用。既往对该区域小兽和寄生蚤类的区系分布特征、物种多样性空间分布格局及其与地理环境因素关系等内容进行过一系列的研究(龚正达等,2001a; 龚正达等,2001b, 2005和2007;权寿瑛等,2009)。此外,还对蚤类科、属和种丰富度之间空间分布特征及其关系进行了分析和讨论(李栋等,2014)。
多年来,有不少研究者对云南不同地区或环境类型的小兽和寄生蚤类多样性也开展了许多的调查研究,如:滇南山地8县(市)小兽及其体表寄生蚤物种多样性的测定和比较(张胜勇等,2006);滇西北剑川、兰坪和鹤庆三县野鼠鼠疫疫源地农田鼠、蚤类多样性及空间分布格局、特征及与地理环境因素关系等问题进行分析和探讨(连宏宇,2010);在西双版纳、临沧和普洱三州(市)的鼠、蚤类多样性及空间分布格局、特征及与地理环境因素关系的调查分析(刘霖,2014);滇西南6县居民区、农耕区和林区3种不同景观小兽及其寄生蚤的调查(黄涛华等,2016);滇西和滇南地区17县(市)居民区鼠体寄生蚤多样性的调查(高子厚等,2014);云南5个不同地理小区内43个县(市)居民区小兽和寄生蚤的多样性、丰富度和密度的关系分析(刘正祥等,2016);云南22县农耕区小兽的群落结构及多样性空间分布特征的分析(陈星等,2019);应用DNA条形码技术对临沧大雪山小型兽类的分类鉴定(马英等,2020)等。
近期,我们在滇西南家鼠鼠疫疫源地8县(市)山地农耕区所获小兽7科19属35种,寄生蚤6科16属27种(亚种)调查研究基础上,就当地蚤类物种多样性空间分布格局与人间鼠疫之间的关系进行分析和探讨,结果发现:蚤类物种多样性、均匀度与人间鼠疫均呈显著负相关(r=-0.910,P<0.05;r=-0.836,P<0.05),而生态优势度与人间鼠疫呈显著正相关(r=0.905,P<0.05);但总蚤指数与物种多样性、均匀度、生态优势度、物种丰富度和病例构成比均不相关。研究认为滇西南家鼠鼠疫疫源地农耕区蚤类物种多样性和优势种印鼠客蚤Xenopsyllacheopis物种多度的空间分布格局和特征与当地人间鼠疫的发生与否、流行强度都存在重要的关系(琚骏科等,2021)。但是,上述系列研究均未涉及山地农耕区小兽与寄生蚤多样性之间关系的研究内容。
本文在滇西南8县(市)山地农耕区所获小兽7科19属35种及其寄生蚤6科16属27种(亚种)蚤类调查数据的基础上,对当地小兽与寄生蚤物种多样性之间关系进行了分析,结果显示:小兽的物种多样性、物种丰富度、均匀度和生态优势度与蚤类丰富度之间(相关系数分别为:r= 0.937,P<0.01;r= 0.899,P<0.01;r= 0.946,P<0.01;r= -0.954,P<0.01)均呈极显著相关,说明当地小兽与寄生蚤之间存在着协同进化与共存的关系; 但是,小兽与蚤类的物种多样性之间(r= 0.51,P>0.05)和生态优势度之间(r= 0.52,P>0.05)的关系不甚密切,因此也反映出两者物种多样性和优势种及其多度的空间分布格局、特征之间存在的差异。这种差异的形成,主要是因为横断山区蚤类与主要宿主动物ɑ多样性的地理分布趋势及它们与重要环境因素的关系均有所不同,前者(蚤类)与水湿(降水量和雨季长短)条件密切相关(龚正达等, 2001a), 而后者(小兽类)主要受纬度的影响,其次是水湿条件(龚正达等, 2001b)。生态优势度指数是反映群落内各物种数量变化情况的测定,生态优势度指数越大,说明群落内物种数量分布越不均匀,优势种的地位越突出,群落的物种多样性越低。该区域山地农耕区小兽寄生蚤物种多样性与生态优势度之间密切相关(r= -0.994,P<0.01)(琚骏科等,2021);小兽物种多样性与生态优势度密切相关(r= -0.966,P<0.01),他们共同反映了优势种的组成及其多度的空间分布格局影响和主导了该区域农耕区小兽及寄生蚤类物种多样性(α多样性)的变化趋势。此结果与滇西北金沙江流域蚊类多样性空间分布格局与优势种多度关系的研究结论(李栋等,2011)类似。
3.2 山地农耕区小兽和寄生蚤多样性及其与鼠疫关系
农田生物多样性(farmland biodiversity)通常特指农田生态系统中的生物多样性,可以理解为在农田进行生长、繁殖、栖息、取食、迁移、避难等活动的所有物种及其相互复杂的营养级联系(Fahrigetal, 2011)。鼠疫疫源地生物群落是一个有机整体,包括鼠疫主要宿主和媒介以及鼠疫菌主要基因组型,它们在进化中互相依赖、互相制约、相互适应、同步进化,形成了鼠疫自然疫源地(龚正达等,2012)。
滇西南家鼠鼠疫疫源地的农耕区小兽寄生蚤类物种多样性、均匀度与人间鼠疫均呈显著负相关,而生态优势度与人间鼠疫呈显著正相关;在蚤类物种多样性、均匀度高而优势度较低的区域,人间鼠疫发生率较低或未发生鼠疫;反之,人间鼠疫的发生率则较高(琚骏科等,2021);此外,优势种印鼠客蚤物种多度及其空间分布特征与人间鼠疫的发生与否和流行强度之间都具有重要关系,这总体反映出滇西南家鼠鼠疫疫源地农耕区蚤类物种多样性和优势种印鼠客蚤物种多度的空间分布格局和特征与当地人间鼠疫的发生与否、流行强度都存在重要的关系(琚骏科等,2021)。由于小兽种类多,分布广,迁移性较小,不同种类的组成及空间分布特征又多与地理环境、气候条件、生态适应能力及鼠疫等自然疫源性疾病的关系密切。因此,进一步开展农田小兽和寄生蚤多样性空间分布格局和特征及当地鼠疫流行或疫源地长期保存机制之间关系的研究有重要意义。
云南西部横断山区已知广泛存在家鼠型鼠疫和野鼠型鼠疫两类型疫源地,其中,以黄胸鼠为主要宿主及其寄生蚤印鼠客蚤为主要媒介的家鼠鼠疫疫源地分布于北纬25°50′以南,滇西南横断山脉区南部和滇南等海拔2 000 m以下的山间盆地城镇或乡村居民区;以齐氏姬鼠、大绒鼠或玉龙绒鼠为主要宿主及其寄生蚤特新蚤指名亚种Neopsyllaspeicalisspeicalis、棕形额蚤指名亚种Frontopsyllaspadixspadix等为主要媒介的野鼠鼠疫疫源地主要局限于北纬26°00′以北,滇西北横断山中部,以剑川县沙溪石龙村为分布中心,长期固着于海拔2 300~3 500 m的山间小盆地、溪流交汇处的林缘或亚高山草甸的农耕区一带(Guoetal.,2000;俞东征,2009; 龚正达等,2018;琚骏科等,2021)。但是,近年从滇西北玉龙县鼠疫疫源地农耕区野鼠体内分离到的鼠疫菌与云南剑川菌株(野鼠型)及云南家鼠型菌株生化特性不同,而与西藏旱獭鼠疫菌的生化特性接近,属于喜马拉雅旱獭青藏高原生态型(郭英等,2008)。喜马拉雅旱獭鼠疫疫源地的主要宿主是喜马拉雅旱獭Marmotahimalayana,其主要媒介为谢氏山蚤Oropsyllasilantiewi和斧形盖蚤Callopsylladolabris,而近年在滇西北香格里拉市山区的调查也证实了当地喜马拉雅旱獭体表和洞穴内有谢氏山蚤和斧形盖蚤的存在和分布(洪梅等,2013)。鉴于目前上述宿主、媒介和病原存在的问题与情况,我们认为滇西北横断山“三江并流”山区高寒草原地带也可能存在喜马拉雅旱獭鼠疫疫源地。因此,进一步开展滇西北野鼠型鼠疫疫源地农耕区小兽和寄生蚤多样性空间分布格局与特征的研究,探讨家、野鼠疫疫源地之间,以及他们与藏东南、川西南地区喜马拉雅旱獭鼠疫疫源地之间的关系均具有重要的理论与实践价值。
