Sinomenine promotes differentiation of induced pluripotent stem cells into immature dendritic cells with high induction of immune tolerance
2022-10-08XiaoYanHuangZhanKuiJinMengDouBingXuanZhengXiangRongZhaoQingFengYangMengFengXiangLongDuanPuXunTianCuiXiangXu
Xiao-Yan Huang, Zhan-Kui Jin, Meng Dou, Bing-Xuan Zheng, Xiang-Rong Zhao, Qing Feng, Yang-Meng Feng,Xiang-Long Duan, Pu-Xun Tian, Cui-Xiang Xu
Xiao-Yan Huang, Xiang-Rong Zhao, Qing Feng, Yang-Meng Feng, Cui-Xiang Xu, Shaanxi Provincial Key Laboratory of Infection and Immune Diseases, Shaanxi Provincial People’s Hospital, Xi'an 710068, Shaanxi Province, China
Zhan-Kui Jin, Department of Orthopedics, Shaanxi Provincial People’s Hospital, Xi'an 710068,Shaanxi Province, China
Meng Dou, Bing-Xuan Zheng, Pu-Xun Tian, Department of Kidney Transplantation, The First Affiliated Hospital of Xi’an Jiaotong University, Xi'an 710061, Shaanxi Province, China
Xiang-Long Duan, Second Department of General Surgery, Shaanxi Provincial People's Hospital, Xi'an 710068, Shaanxi Province, China
Abstract BACKGROUND Immature dendritic cells (imDCs) play an important role in the induction of donor-specific transplant immunotolerance. However, these cells have limitations,such as rapid maturation and a short lifespan in vivo. In previous studies, induced pluripotent stem cells (iPSCs) differentiated into imDCs, and sinomenine (SN)was used to inhibit the maturation of imDCs.AIM To study the capacity of SN to maintain iPSC-derived imDCs (SN-iPSCs-imDCs)in an immature state and the mechanism by which SN-iPSCs-imDCs induce immunotolerance.METHODS In this study, mouse iPSCs were induced to differentiate into imDCs in culture medium without or with SN (iPSCs-imDCs and SN-iPSCs-imDCs). The imDCrelated surface markers, endocytotic capacity of fluorescein isothiocyanate-Dextran and apoptosis were analyzed by flow cytometry. The effects of iPSCs-imDCs and SNiPSCs-imDCs on T-cell stimulatory function, and regulatory T (Treg) cell proliferative function in vitro were analyzed by mixed lymphocyte reaction. Cytokine expression was detected by ELISA.The apoptosis-related proteins of iPSCs-DCs and SN-iPSCs-DCs were analyzed by western blotting. The induced immunotolerance of SN-iPSCs-DCs was evaluated by treating recipient Balb/c skin graft mice. Statistical evaluation of graft survival was performed using Kaplan-Meier curves.RESULTS Both iPSCs-imDCs and SN-iPSCs-imDCs were successfully obtained, and their biological characteristics and ability to induce immunotolerance were compared. SN-iPSCs-imDCs exhibited higher CD11c levels and lower CD80 and CD86 levels compared with iPSCs-imDCs. Reduced major histocompatibility complex II expression, worse T-cell stimulatory function, higher Treg cell proliferative function and stronger endocytotic capacity were observed with SN-iPSCs-imDCs (P < 0.05).The levels of interleukin (IL)-2, IL-12, interferon-γ in SN-iPSCs-imDCs were lower than those in iPSCs-imDCs, whereas IL-10 and transforming growth factor-β levels were higher (P < 0.05). The apoptosis rate of these cells was significantly higher (P < 0.05), and the expression levels of cleaved caspase3, Bax and cleaved poly(ADP-ribose) polymerase were higher after treatment with lipopolysaccharides, but Bcl-2 was reduced. In Balb/c mice recipients immunized with iPSCsimDCs or SN-iPSCs-imDCs 7 d before skin grafting, the SN-iPSCs-imDCs group showed lower ability to inhibit donor-specific CD4+ T-cell proliferation (P < 0.05) and a higher capacity to induce CD4+CD25+FoxP3+ Treg cell proliferation in the spleen (P < 0.05). The survival span of C57bl/6 skin grafts was significantly prolonged in immunized Balb/c recipients with a donor-specific pattern.CONCLUSION This study demonstrated that SN-iPSCs-imDCs have potential applications in vitro and in vivo for induction of immunotolerance following organ transplantation.
Key Words: Immature dendritic cells; Induced pluripotent stem cells; Sinomenine; Immune tolerance; Organ transplantation
lNTRODUCTlON
Although solid organ transplantation has become the most effective and promising treatment option for end-stage organ failure, it is limited by immune rejection. The advancement of medical technology and development of new immunosuppressive regimens have lowered the rates of acute rejection but appear to have had little impact on the incidence of chronic rejection; the principal cause of late graft loss[1,2].Therefore, it is necessary to seek a therapeutic approach with minimal side effects and high safety and effectiveness to inhibit chronic rejection, ultimately inducing a state of donor-specific tolerance.
Dendritic cells (DCs) are the most potent antigen-presenting cells and play a dual role in the immune response. Mature DCs participate in the priming of naïve T cells to initiate an immune response, while immature DCs (imDCs) induce immune hyporesponsiveness or tolerance[3]. Cumulative evidence has demonstrated that imDCs induce immunotolerance and immune hyporesponsiveness because they do not induce expression of costimulatory molecules or promote the proliferation of CD4+CD25+regulatory T (Treg) cells[4-6]. However, DCs constitute only 1% of human peripheral blood[7], and can be obtained from a limited number of sources. Additionally, DCs from external sources are prone to maturation after infusionin vivo.
Induced pluripotent stem cells (iPSCs) display the classic features of embryonic stem cells such as differentiation capacity and immunomodulation. Importantly, they have the main advantages of convenient acquisition, broad applicability, noninvasiveness, and lack of immunorejection and ethical issues[8]. They can also be induced into DCsin vitro[9], which solves the problem of limited sources for DCs. However, maintaining an immature state for DCs over a long period is another critical challenge to be addressed.
Sinomenine (SN), a Chinese medicinal product, is reported to be a potential therapeutic tool for inducing immunotolerance[10,11]. Some studies have confirmed that donor-derived DCs infused before transplantation were maintained in the immature stage by SN, which increased splenic FoxP3+Treg cells in recipient rats after renal allotransplantation[12] and prolonged the survival time of allografts[13]. In addition, SN reduced the expression of costimulatory molecules on DCs, such as CD86 and CD40[14],and enhanced the function of CD4+CD25+Treg cells[15]. However, whether SN can maintain the immature stage of iPSC-derived DCs to induce immune toleranceremains to be investigated.
We aimed to study the capacity of SN to maintain iPSC-imDCs (SN-iPSCs-imDCs) in an immature state as well as the mechanism by which SN-iPSCs-imDCs induce immunotolerancein vitro. Our findings should provide a theoretical and experimental basis for the application of SN-iPSCs-imDCs for the induction of immunotolerance following organ transplantation.
MATERlALS AND METHODS
Cell lines and reagents
iPSCs lines MiPS.5 were purchased from Shanghai Sisansai Biological Technology Co Ltd (Catalogue No. 0225-100) and maintained in serum-free and feeder-free medium (DMEM/F12, Neurobasal medium, 0.5% N2, 1% B27, 0.1% 2-mercaptoethanol (ME), 1% Penicillin-Streptomycin, 0.1‰StemoleculeTM PD0325901, 0.32‰ StemoleculeTM CHIR99021, 0.1‰ LIF). Mouse bone marrow stromal cells, OP9, were maintained in DMEM supplemented with 10% fetal calf serum (FCS) and seeded onto gelatin-coated dishes before use as feeder cells. Recombinant mouse interleukin (IL)-4 and granulocyte macrophage colony-stimulating factor (GM-CSF) were obtained from Perprotech (London, United Kingdom). Lipopolysaccharide (LPS) was purchased from Sigma Chemical (St. Louis, MO). Monoclonal antibodies of rabbit anti-mouse fluorescein isothiocyanate (FITC)-CD45, FITC-CD11b, FITC-CD80, FITCCD86, FITC-MHC-II, FITC-CD11c, CD4, CD25, FoxP3, Bcl-2, Bax, Caspase3, and poly(ADP-ribose)polymerase (PARP) were obtained from Biolegend (California, United States). SN was obtained from Xidiai Chemical Industrial Development Co Ltd (Shanghai, China).
SN optimal concentration screening assay
iPSCs were treated with different concentrations of SN (0, 50, 100 or 200 μM). The optimal concentration of SN was determined with the Cell Counting Kit-8 (CCK-8) assay. The optimal concentration of the drug was defined as the maximum administered concentration at which cell viability was > 95%.
iPSC identification
Polymerase chain reaction (PCR) Total RNA was extracted from resuscitated iPSCs, and reverse transcription was performed on ice to obtain cDNA. Afterward, quantitative PCR (qPCR) was performed using the Green PCR Master Mix System (Thermo Fisher Scientific). β-actin expression was used to normalize all the data. The primer sequences used are listed in Table 1.
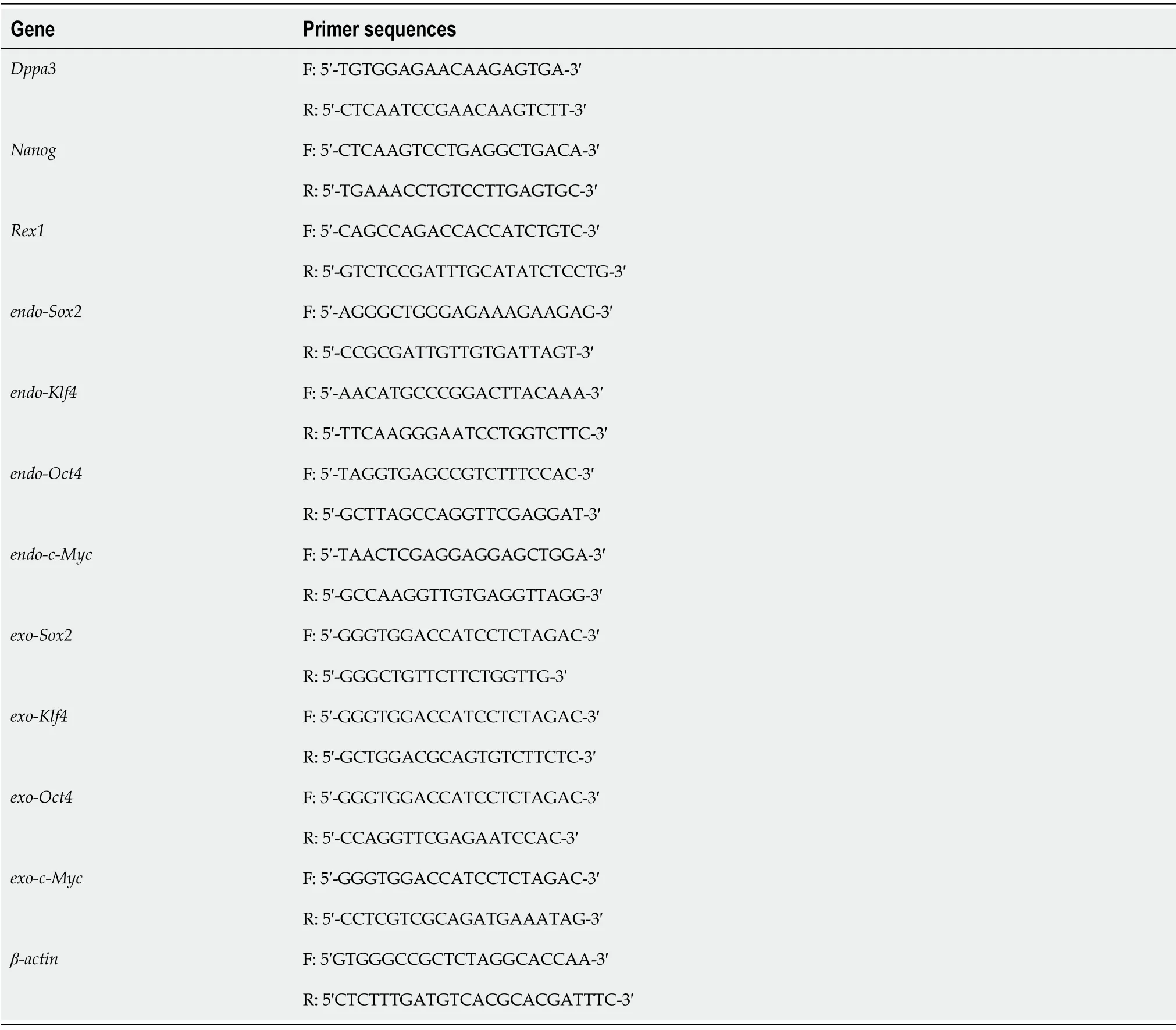
Table 1 Primer sequences of polymerase chain reaction
Alkaline phosphatase staining
Resuscitated iPSCs were cultured on sterile glass slides for 24 h, fixed, washed, and stained in a freshly prepared alkaline-dye mixture. The cells were exposed to the dark at 18-26 °C for 15 min, then observed under a microscope and photographed for analysis.
Karyotype analysis
iPSCs were collected and treated with 2% colchicine for 2 h, then treated with 0.075 M KCL in 37 °C for 30 min, and pre-fixed with a newly prepared fixative (methanol: Glacial acetic acid = 3:1). The supernatant was discarded, the cells were fixed again for 30 min, dropped onto slides, and treated with 0.125% trypsin solution for 2 s. After staining with Giemsa stain for 10 min, karyotype analysis was performed using chromosome karyotype analysis software (BEION V4.20 -C).
Teratoma formation
iPSCs (5 × 106) were injected subcutaneously into the right flank of Balb/c nude mice. The mice wereobserved to determine the time of teratoma formation and differentiation into ectoderm, mesoderm, and endoderm tissue.
Differentiation culture
The procedure for inducing differentiation of iPSCs into DCs comprised three stages[9]. Stage 1: In the presence or absence of 100 μM SN, iPSCs were suspended in α-minimal essential medium (MEM)supplemented with 20% FCS and seeded (105cellsperdish) onto OP9 cell layers. On day 7, the cells were collected for stage 2. Stage 2: In the presence or absence of 100 μM SN, cells harvested from the stage 1 culture were resuspended in α-MEM supplemented with 20% FCS, GM-CSF (10 ng/mL), and 2-ME (50 mmol/L) for 6 d. Stage 3: In the presence or absence of 100 μM SN, the cells were cultured in RPMI-1640 medium supplemented with 10% FCS, 100 μM SN, GM-CSF, and 2-ME without feeder cells. On day 5 or 6 after initiation of stage 3, the cells were stimulated with LPS (5 mg/mL) for 48 h then collected for subsequent analysis.
Mixed lymphocyte reaction
CD4+T cells from Balb/c mouse spleen were isolated with MACS immunomagnetic beads. The iPSCs-DCs and SN-iPSCs-DCs groups were treated with 25 μg/mL mitomycin. The control consisted of RPMI-1640 medium containing 5 μg/mL LPS. CD4+T cells were cocultured with iPSCs-DCs, SN-iPSCs-DCs, or controls in different proportions (DCs:T cells = 1:80, 1:40, 1:20 or 1:10) in a 96-well culture plate for 72 h.The inhibition rates of CD4+T cells by iPSCs-DCs and SN-iPSCs-DCs were analyzed with the CCK-8 assay based on the optical density (OD). Inhibition rate = (1 - OD of experimental group/OD of control group) × 100%.
Flow cytometry
The surface markers of iPSCs-imDCs and SN-iPSCs-DCs, the effects of iPSCs-DCs and SN-iPSCs-DCs on Treg cellsin vitroandin vivo, apoptosis, and uptake of fluorescein isothiocyanate (FITC)-Dextran in cells were analyzed by flow cytometry. Cells were collected and labeled with primary anti-CD45,CD11b, CD80, CD86, MHC-II, CD11c, CD25, CD4, and FoxP3, followed by secondary Alexa-Fluor488-conjugated goat anti-rabbit IgG antibodies. The cells were analyzed using BD FACS flow cytometry, and data were analyzed using FlowJo software.
ELISA
After stimulation with LPS at a final concentration of 5 μg/mL for 48 h, the iPSCs-imDCs and SN-iPSCsimDCs culture supernatants were collected. Expression of cytokines IL-2, IL-10, interferon (IFN)-γ,transforming growth factor (TGF)-β and IL-12 in the culture supernatant before and after LPS stimulation was detected by ELISA. IL-2, IL-10, IFN-γ, TGF-β and IL-12 expression in the peripheral blood of recipients at 14 d postgraft was also detected by ELISA.
Western blotting
The relative expression of apoptosis-related proteins caspase 3, Bcl-2, Bax, and PARP were analyzed by western blotting. After stimulating iPSCs-imDCs and SN-iPSCs-imDCs with LPS for 48 h, the total protein for the two groups was extracted with radioimmunoprecipitation assay lysis buffer and measured with a bicinchoninic acid protein assay kit. The same amount of protein from lysate samples was separated using SDS-PAGE and transferred to nitrocellulose membranes blocked with 50 g/L skimmed milk for 2 h. The membranes were incubated with primary antibodies overnight at 4 °C,followed by horseradish-peroxidase-conjugated secondary antibodies for 1 h at room temperature. The blots were visualized and detected using an ECL Western blot analysis system, and glyceraldehyde 3-phosphate dehydrogenase expression was used to normalize the data. The intensity of the western blotting bands was determined with Image J software version 1.46 (National Institutes of Health,Bethesda, MD, United States).
Skin transplantation
C57bl/6 mice were used as donors and Balb/c mice were used as recipients. Thirty Balb/c mice (6-7 week old, 19.9 ± 2.8 g) were randomly divided into six groups: (1) Control group: Allogeneic mouse skin graft model without treatment; (2) PBS group: 7 d before transplantation, 0.3 mL PBS was intravenously infused into the tails of the recipients; (3) iPSCs-imDCs group: 7 d before transplantation, 106iPSCsimDCs in 0.3 mL PBS were intravenously infused into the tails of the recipients; (4) SN-iPSCs-imDC group: 7 d before transplantation, 106SN-iPSCs-imDCs in 0.3 mL PBS were intravenously infused into the tails of the recipients; (5) SN group: 3 d before transplantation, 30 mg/kg SN was intravenously infused into the tails of the recipients, once a day for 3 d; and (6) Cyclosporin A (CsA) group: 3 d before transplantation, 30 mg/kg CsA was intravenously infused into the tails of the recipients, once a day for 3 d. Allogeneic skin transplantation was performed according to the protocol described previously[16].The donor mice were anesthetized with 50 mg/kg pentobarbital sodium (intraperitoneal), and back skin was harvested with blunt dissection. The donor animals were killed, and the connective tissue, fat tissue, and panniculus carnosus were removed from the skin flap to obtain 1 cm × 1 cm squares. The recipient mice were anesthetized with the same dose of pentobarbital sodium (intraperitoneal) plus 5 mg/kg carprofen (subcutaneous). Graft beds (1 cm × 1 cm) were created by dissecting the back skin,keeping the sarcolipid membranes and vessels intact. The allografted skin sections were stitched together using 4-0 silk, and 0.5 mg gentamicin was administeredpermouse to prevent infection. The weight of the animals was recorded every 7 d for 28 d. From day 4 post-transplant, the grafts in each group were scored once a day for 24 d. Statistical evaluation of graft survival was performed using Kaplan-Meier curves and compared using log-rank tests. On day 14 post-transplant, the allografts and recipients’ spleens were removed for analysis.
Treg cell detection in animal spleens
The recipient spleens in both groups were ground and filtered through a 70-μm cell strainer. The CD4,CD25, and FoxP3 markers on Treg cells were analyzed by flow cytometry. Using C3H mice as an unrelated third party, the reactivity of the recipient lymphocytes in the PBS, iPSs-imDCs and SN-iPSsimDCs groups to the donor and unrelated third party was detected.
Hematoxylin and eosin staining
The allografts were harvested, fixed in 4% paraformaldehyde for 24 h, dehydrated, and embedded in paraffin. Four-micrometer-thick sections were stained with hematoxylin and eosin (H&E) for morphological observation. Images were observed and photographed under the BX41 fluorescence microscope(Olympus Corporation, amplification: × 200).
Statistical analysis
All data were expressed as the mean ± SEM. The biotechnology experiments were repeated at least three timesin vitro. Intergroup deviations were assessed using one-way analysis of variance, withP< 0.05 indicating a significant difference.
RESULTS
iPSCs culture and identification
Morphology of iPSCs was observed after culturing for 2 d, 3 d, 5 d and 6 d, and fluorescence images showing expression of Nanog+-iPSCs colonies (Figure 1A). The specific genesNanog,Rex1andDppa3and endogenous genesSox2,Klf4,c-MycandOct4in the iPSCs remained stable as the number of passages increased, and no exogenousSox2,Klf4, c-MycandOct4gene expression occurred (Figure 1B).Alkaline phosphatase staining of the iPSCs was positive; the cells were stained purple-black, which is an important characteristic of pluripotent stem cells (Figure 1C). Karyotype analysis showed that the iPSCs possessed a normal diploid karyotype of 40XX (Figure 1D). The differentiation potential of iPSCs into various cell types in the three germ layers was determined by analyzing teratomas generated from mouse iPSCs. After H&E staining, the intestinal epithelium (endoderm), muscle tissue (mesoderm) and nerve tissue (endoderm) were observed under a microscope (Figure 1E). The experiment demonstrated that iPSCs possessed the potential to differentiate into three germ layer tissue.
SN enhances the differentiation of iPSCs into imDCs
The CCK-8 assay results showed the optimal concentration of SN was 100 μM. This was the maximum concentration at which cell viability was > 95% (Figure 2A). iPSCs began to differentiate at 3 d in the first stage; bone marrow hematopoietic progenitor-like cells appeared at 6 d in the second stage; and the cells differentiated into imDC-like cells at 13 d in the third stage. After 48 h of LPS stimulation, DC-like cells appeared in large numbers, with a uniform size, short burrs, and protrusions in individual cells.Scanning electron microscopy showed that the cells were wrinkled and uniform, with burr-like protrusions on the surface. Transmission electron microscopy showed that cells contained mitochondria,ribosomes and other organelles, as well as vesicles and lysosomes of varying sizes (Figure 2B). The iPSCs that differentiated on day 6 in the second stage were collected and detected with flow cytometry.The results showed that the cell surface highly expressed the hematopoietic cell marker antigen CD45 and monocyte marker antigen CD11b (Figure 2C).
SN-iPSCs-imDCs surface phenotypes in differentiation culture
The cells in the iPSCs-imDCs and SN-iPSCs-imDCs groups were collected on days 5, 10 and 13 in the third stage after 48 h of LPS stimulation and visualized by flow cytometry. On days 5, 10 and 13,expression of CD80, CD86, and MHC-II on the cell surface of the two groups was lower, and expression of CD11c was higher. After LPS stimulation of iPSCs-imDCs and SN-iPSCs-imDCs for 48 h, expression of CD80, CD86 and MHC-II increased, while expression of CD11c decreased. The positive proportion of CD80, CD86 and MHC-II on the surface of the SN-iPSC-DCs group was significantly lower than that in the iPSC-DCs group, while expression of CD11c was significantly higher (Figure 3).
SN-iPSCs-imDCs suppressed T-cell proliferation and enhanced Treg cell proliferation
The spleen cells of BALB/c mice were isolated. The purity of T cells detected by CD3 (FITC)-specific fluorescent antibody after magnetic bead separation was 93.42 ± 2.64% (Figure 4A). iPSCs-imDCs, SNiPSCs-imDCs and LPS were cocultured with sorted T-cells for 72 h, respectively. The ratio of DCs:T cells was 1:10, 1:20, 1:40 and 1:80 in each group. The proliferation of stimulated T cells in the SN-iPSCs-DCs group was significantly lower than that in the LPS group (Figure 4B). When the ratio of imDCs:T cells was 1:40, the proliferation of stimulated T cells in the SN-iPSCs-DCs group was significantly lower than that in the iPSCs-DCs group (Figure 4C). iPSCs-DCs and SN-iPSCs-DCs were cocultured with CD4+T cells at a ratio of 1:40 for 72 h, and the inhibition of CD4+T cells in each group was determined with the CCK-8 assay. The results showed that the ability of SN-iPSCs-DCs to inhibit proliferation of CD4+T cells was better than that of iPSCs-DCs (Figure 4D). Flow cytometry showed that iPSCs-DCs and SN-iPSCs-DCs had the ability to induce CD4+CD25+FoxP3+Treg cell proliferation. The ability of SN-iPSCs-DCs to induce proliferation of CD4+CD25+FoxP3+T cells was significantly higher than that of iPSCs-DCs(Figure 4E). The cell culture supernatants were collected, and ELISA was used to detect cytokines IL-2,IL-12, IFN-γ, IL-10 and TGF-β. The levels of IL-2, IL-12 and IFN-γ in the culture medium of the SNiPSCs-DCs group were significantly lower than those of the iPSCs-DCs group, whereas the levels of IL-10 and TGF-β were significantly higher in the SN-iPSCs-DCs group (Figure 4F).
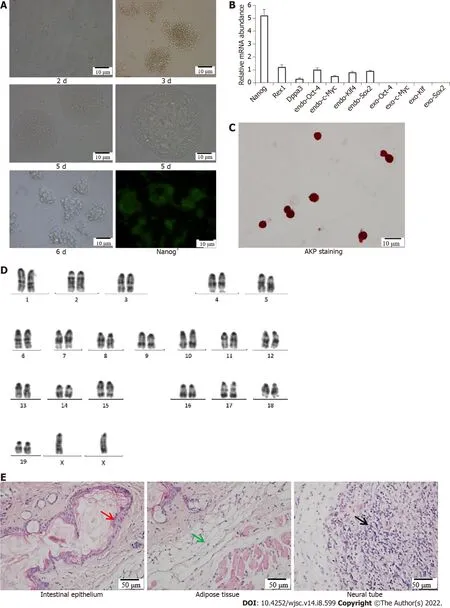
Figure 1 lnduced pluripotent stem cells culture and identification.
Endocytotic capacity in SN-iPSCs-imDCs and SN-iPSCs-DCs
iPSCs-imDCs and SN-iPSCs-imDCs were treated with LPS (5 μg/mL) for 48 h to obtain iPSCs-DCs and SN-iPSCs-DCs. The antigen uptake assays used FITC-Dextran to examine the endocytic ability in the generated SN-iPSCs-imDCs and SN-iPSCs-DCs. Antigen uptake was greater at 37 °C than 4 °C in the same group at the same time. Antigen uptake in SN-iPSCs-imDCs was greater than that observed in the iPSCs-imDCs at 37 °C. Endocytotic capacity of FITC-Dextran was also significantly higher in SN-iPSCs-DCs than iPSCs-DCs (Figure 5).
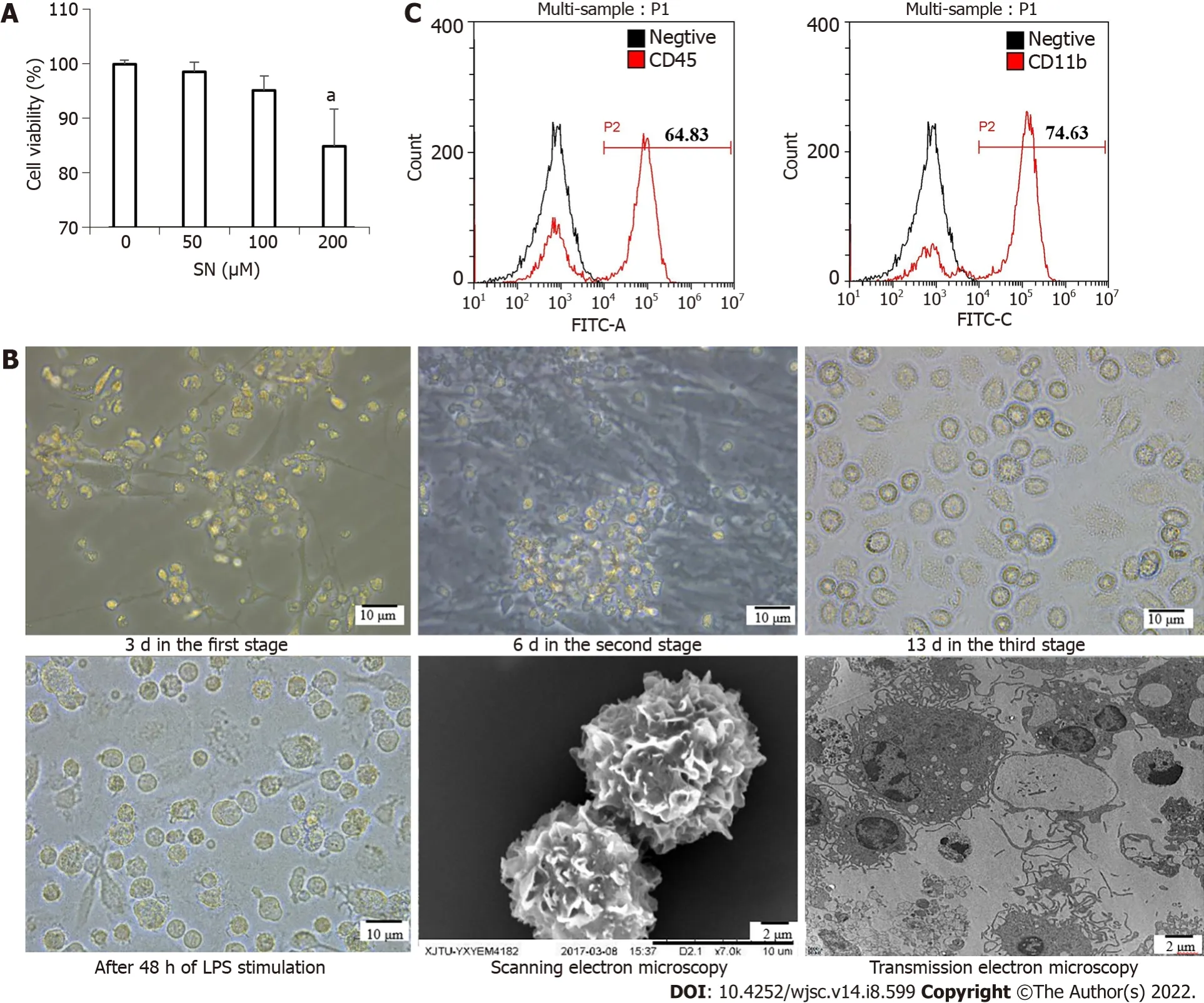
Figure 2 Morphological changes from induced pluripotent stem cells to induced pluripotent stem cells-dendritic cells.
Apoptosis of SN-iPSCs-DCs stimulated with LPS
iPSCs-imDCs and SN-iPSCs-imDCs were treated with LPS (5 μg/mL) for 48 h, and apoptosis was analyzed by flow cytometry. The apoptosis rate of SN-iPSCs-DCs was significantly higher than that of iPSCs-DCs (Figure 6A). Expression of apoptosis-related proteins caspase 3, Bax, Bcl-2 and PARP was analyzed by western blotting. Protein expression levels of cleaved caspase 3, Bax and cleaved PARP in SN-iPSCs-DCs was higher than in iPSCs-DCs, and the level ofBcl-2gene was lower (Figure 6B).
SN-iPSCs-imDCs immunization prolonged survival of allogeneic skin grafts
The weight of mice in the Sham group increased gradually, whereas the weight in other groups decreased from day 7 postgraft, especially in the CsA group. Compared with the iPSCs-imDCs group,the weight of mice in the SN-iPSCs-imDCs group increased significantly (Figure 7A). Histology showed lymphocyte infiltration around the grafted skin in the PBS group on day 7 postgraft. In contrast,lymphocyte infiltration was markedly reduced in the iPSCs-imDCs and SN-iPSCs-imDCs groups,especially in the latter group (Figure 7B). The graft rejection score was reduced in the SN-iPSCs-imDCs group than in the iPSCs-imDCs group (Figure 7C). We also found that donor SN-iPSCs-imDCs prolonged allograft survival [PBS control:n= 5, median survival time (MST) 8.2 d; iPSCs-imDCs:n= 5,MST 17 d; SN-iPSCs-imDCs: MST 22 d; SN:n= 5, MST 15.8 d; CsA:n= 5, MST 26.2 d] (Figure 7D).
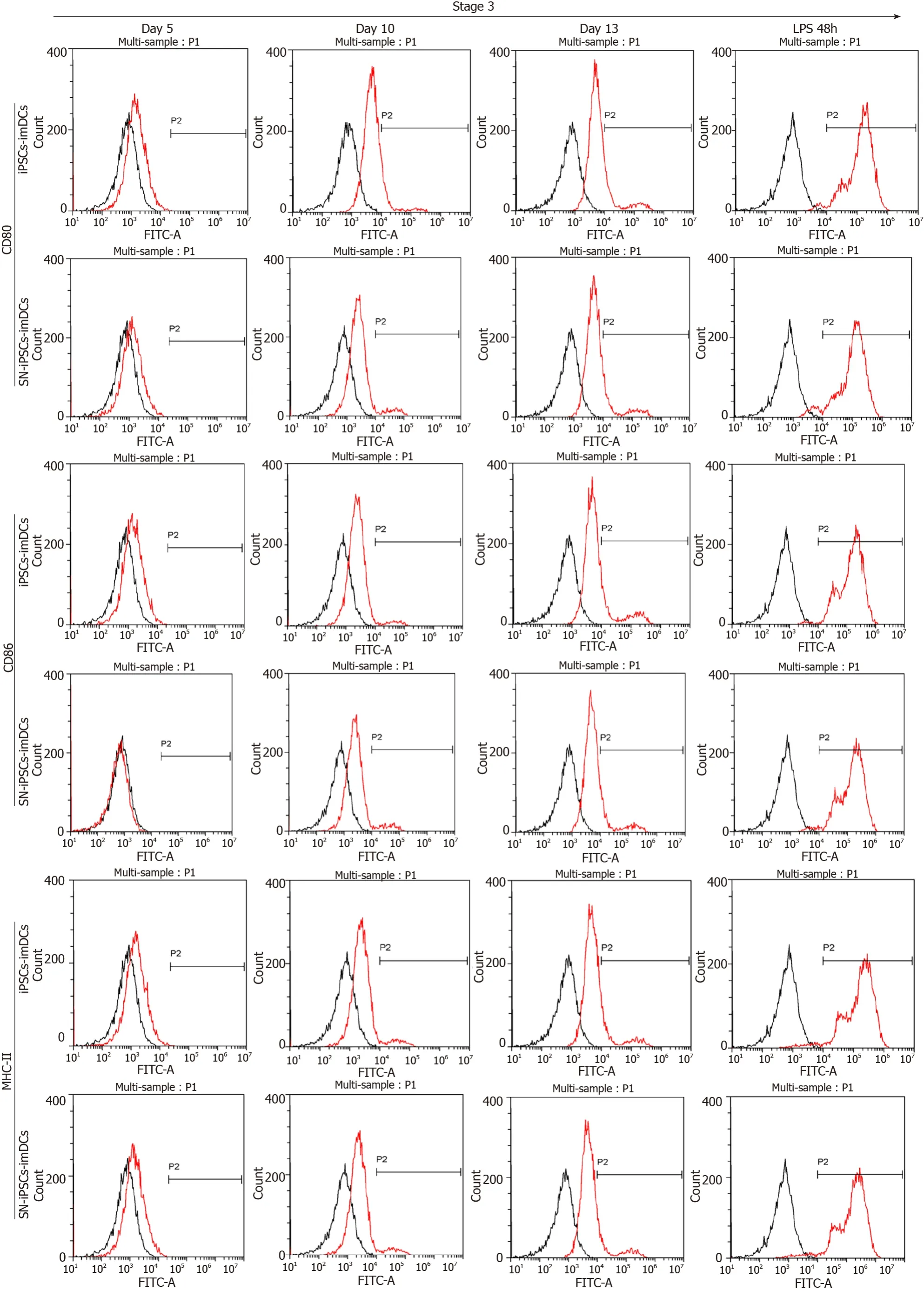
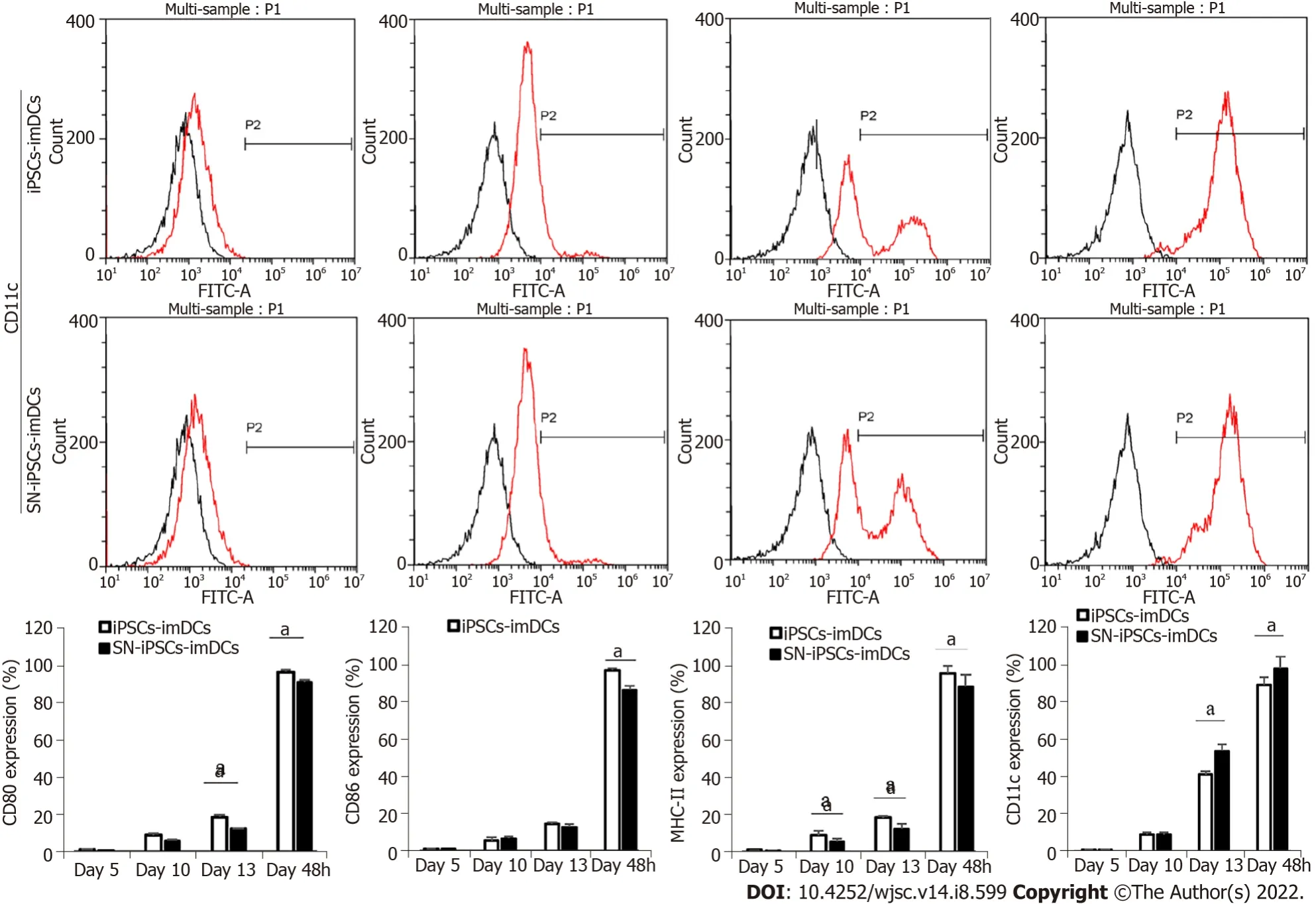
Figure 3 lmmature dendritic cells surface phenotypes in differentiation culture.
SN-iPSCs-imDCs immunization suppressed T cells and increased activated Treg cells in allografts
We found that the ratios of CD4+CD25+Treg cells/CD4+T cells in spleen and CD4+CD25+FoxP3+Treg cells/CD4+CD25+Treg cells were significantly increased in the iPSCs-imDCs and SN-iPSCs-imDCs groups than in the SN group. Compared with iPSCs-imDCs, the SN-iPSCs-imDCs group was higher(Figure 8A and B). The reactivity of spleen lymphocytes to donor mouse lymphocytes in the iPSCsimDCs and SN-iPSCs-imDCs groups was significantly lower than that in unrelated third-party C3H mice. The SN-iPSCs-imDCs group had significantly less reactivity to donor lymphocytes than the iPSCsimDCs group had (Figure 8C), which confirmed that the infusion of donor-derived SN-iPSCs-imDCs could induce donor-specific immune hyporesponsiveness and that SN-iPSCs-imDCs were significantly better than iPSCs-imDCs at this task. Proinflammatory cytokines such as IL-2, IL-12 and IFN-γ were downregulated, and anti-inflammatory cytokines such as IL-10 and TGF-β were upregulated in the serum from SN-iPSCs-imDC-treated recipients compared with the levels in iPSCs-imDCs-treated recipients (Figure 8D).
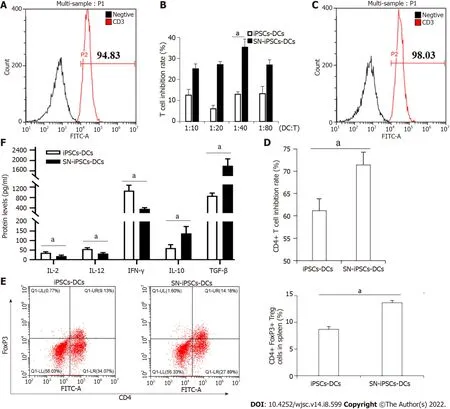
Figure 4 Sinomenine-induced pluripotent stem cells-immature dendritic cells suppress T cell proliferation and enhanced regulatory T cell proliferation.
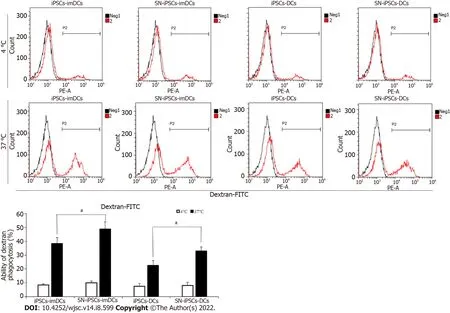
Figure 5 Endocytotic capacity in sinomenine-induced pluripotent stem cells-immature dendritic cells and sinomenine-induced pluripotent stem cells-dendritic cells.
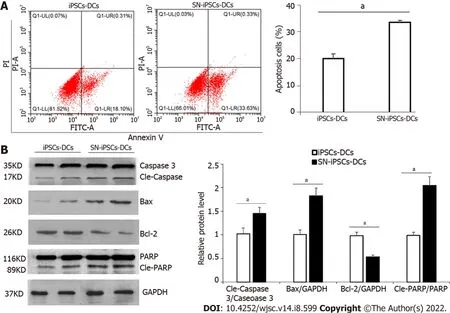
Figure 6 Apoptosis of induced pluripotent stem cells-dendritic cells and sinomenine-induced pluripotent stem cells-dendritic cells stimulated with lipopolysaccharide.
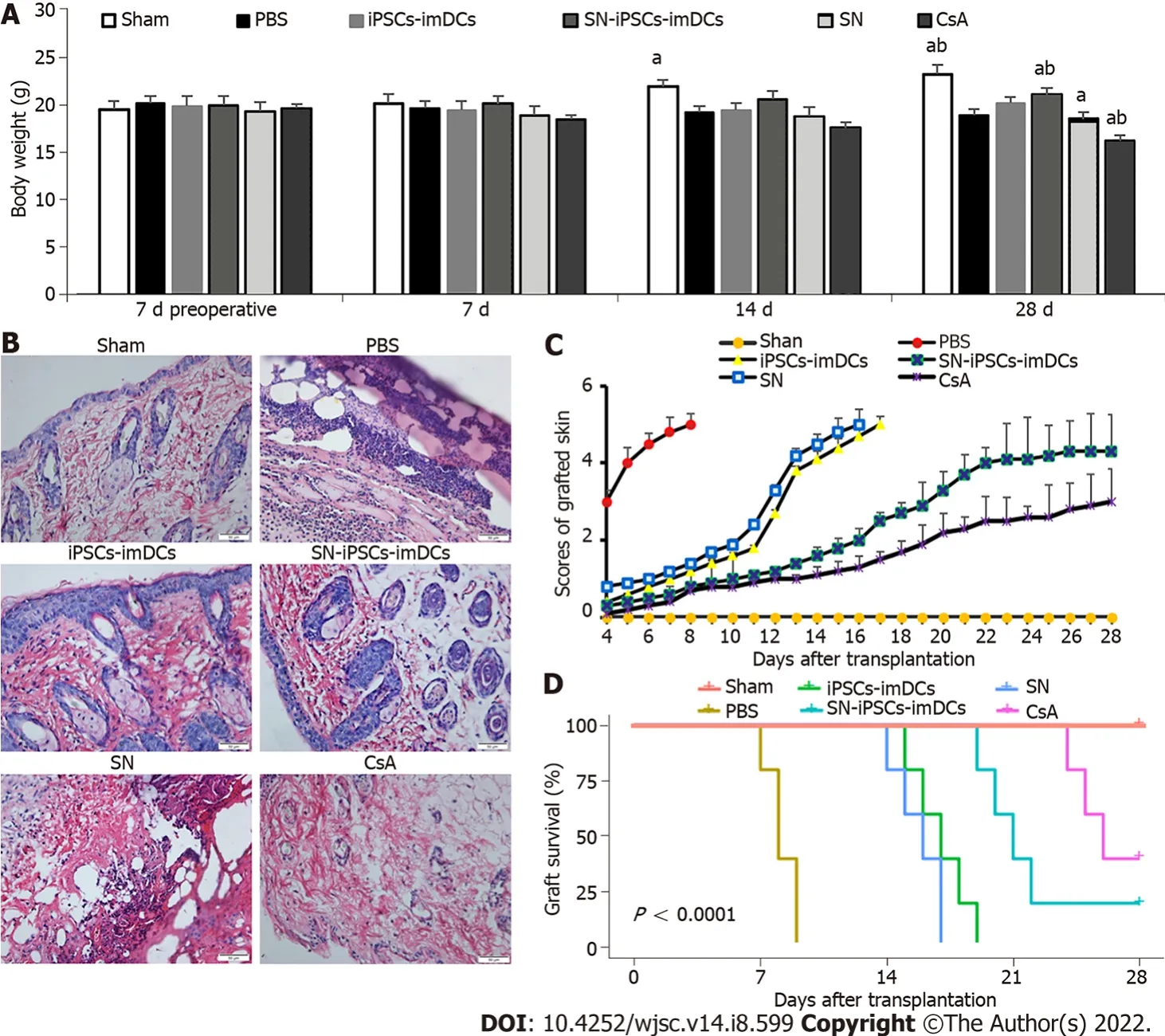
Figure 7 Sinomenine-induced pluripotent stem cells-immature dendritic cells immunization prolonged survival of allogeneic skin grafts.
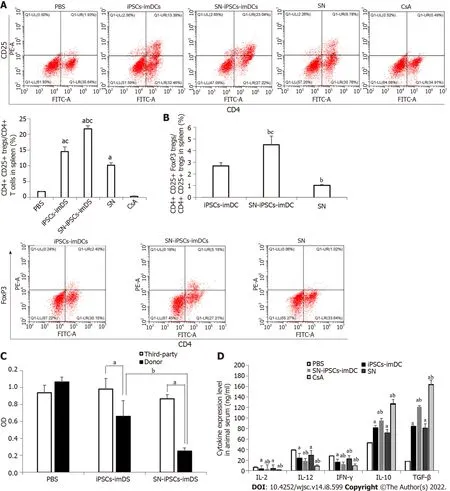
Figure 8 Sinomenine-induced pluripotent stem cells-immature dendritic cells immunization suppressed T cells and increased activated regulatory T cells in allografts.
DlSCUSSlON
imDCs play an important role in immune tolerance but are difficult to obtain in large quantities. At present, DCs can be obtained from bone marrow mesenchymal stem cells, but the process is difficult due to the significant trauma involved. DCs can also be obtained from cord blood and embryonic stem cells; however, ethical issues prevent the clinical application of these cells. iPSCs have been adopted in the biological and medical fields because they can be obtained from a wide range of sources and are free of ethical issues[17]. Previous research and our study confirmed that iPSCs cocultured with OP9 feeder cells could differentiate into imDCs under the influence of GM-CSF and IL-4[9,18].
The immune function of DCsin vivois closely related to their maturity. Mature DCs provide activation signals to initiate immune responses unlike imDCs, which cannot provide a signal for T-cell activation or induce T-cell clonal energy due to the lack of expression of costimulatory molecules on their surface[19,20]. imDCs induce T cells to differentiate into Th2 cells, inhibit secretion of inflammatory factors (IL-2, IFN-γ,etc.), and increase secretion of anti-inflammatory factors (IL-10, TSLP,TGF-β,etc.), eventually inducing an increase in the proportion of Treg cells[21-23]. The increased Treg cells subsequently downregulate expression of costimulatory molecules CD40, CD80 and CD86 in the NF-κB pathway; upregulate inhibitory receptors; induce production of tolerogenic DCs, including imDCs; and maintain the immature state of imDCs[24], thus forming a positive feedback loop that ultimately induces immunotolerance[25]. Due to the strong phagocytic function of imDCs, they mature quickly and lose the ability to induce immunotolerance under antigen stimulationin vivo. At present,the methods for inhibiting the maturation of imDCs mainly include: (1) Blocking the surface costimulatory molecules of imDCs by genetic modification[26-28]; (2) Blocking expression of specific molecules on the surface of imDCs with drugs[29]; (3) Using immunomodulatory factors to inhibit maturation of imDCs[30,31]; and (4) Using immunosuppressive agents to keep imDCs immature[32]. However, there are some inevitable flaws with these strategies. Firstly, the efficacy of gene modification is low, and there are potential biosafety and immunogenicity issues. Secondly, controlling the concentration of imDCs in the body through drug intervention is difficult due to large individual differences. Thirdly,when cytokines are infused into the body, allograft antigen stimulation, microbial infection, and other factors make it difficult for imDCs to maintain their immature state for a long time. Therefore, a strategy to stably maintain the immature state of imDCs has become a primary challenge at present.
In recent years, the potential of SN application in transplantation immunity has received increasing attention. It has been reported that SN can stimulate the differentiation of peripheral blood mononuclear cells into imDCs and further inhibit the maturation of imDCs[13]. SN can reduce the expression of CCR5 and CCR7 on DCs, inhibit secretion of chemokines CXCL9 and CXCL10, block expression of TLR2 and TLR4 in peripheral blood DCs, stimulate differentiation of monocytes into imDCs, and inhibit their further maturation[33]. Due to the mild drug properties of SN, short half-life, lack of toxicity below a certain dose, and lack of mutagenicity, the compound was used to induce bone marrow stem cells to differentiate into imDCs (SN-BM-imDCs). The results demonstrated that the infusion of donor SN-BMimDCs could significantly prolong the graft survival span in recipient rats and induce the proliferation of Treg cellsin vivo[13]. In this study, we successfully obtained SN-iPSCs-imDCs and confirmed these cells had the morphological structure and biological function of imDCs and retained a stable immature phenotype, even in the presence of LPS. Treg cells generated by SN-iPSCs-imDCs immunization were donor-specific and played a key role in tolerance induction and maintenance. However, the study did not explore the efficiency of directed differentiation in detail. In the future, we will apply SN at different stages of iPSCs differentiation and investigate the highest differentiation efficiency of SN-iPSCs-imDCs.
The apoptosis of DCs is closely related to immune status. It has been reported that some pathogens,tumors, and drugs promote the apoptosis of DCs, which can lead to a reduction in antigens presented to T cells. The results indicate that imDCs phagocytose apoptotic DCs, transform into tolerance DCs, and then induce naïve T cells to differentiate into FoxP3+Treg cells[34]. Some studies have also confirmed that SN could inhibit glioblastoma by inducing mitochondria-dependent apoptosis and autophagy by PI3K/AKT/mTOR and AMPK/mTOR pathway[33,34]. However, there is no report on SN-induced apoptosis of DCs. In this study, we found that apoptosis-related protein expression levels in SN-iPSCsimDCs were significantly higher than those in iPSCs-imDCs. This may be one of the mechanisms by which SN-iPSCs-imDCs induce immune hyporesponsiveness or tolerance.
Donor immune cell infusion therapy can enhance recipient immune negative regulation, which is the most effective method to resolve graft rejection[35,36]. In this study, we established a skin transplantation model of C57bl/6 as the donor and BALB/c mice as the recipient as an alternative to the common organ transplantation model. Balb/c mice recipients were immunized with SN-iPSCs-imDCs or iPSCs-imDCs 7 d before skin grafts, and Treg cells were significantly increased in Balb/c mice splenocytes after transplantation. The survival span of C57bl/6 skin grafts was significantly prolonged in immunized Balb/c recipients with a donor-specific pattern, which was consistent with the reports of DCs coordinating the development and homeostasis of organ-specific Treg cells[37,38]. The reason for the induction of tolerance may be that the donor-type SN-iPSCs-imDCs act as a primary vaccine, while the alloantigen acts as a secondary vaccine[16], thereby promoting the expansion and continuous activation of donor-specific Treg cells.
However, there were some limitations to our study. Firstly, the procedure for inducing the differentiation of iPSCs into DCs comprised three stages. Further research is required to establish at which stage intervention with SN improves induction. Secondly, cell density is an important factor determining cell fate. We aim to try different cell densities on different effects of immunotolerance in organ transplantation to obtain the optimal protocol in our future studies. Lastly, we combined SN and mouse iPSCs technology to obtain SN-iPSCs-imDCs. In the future, we will combine SN and human iPSCs technology to induce immunotolerance in a clinical setting.
CONCLUSlON
We combined SN and iPSCs technology for the first time and successfully obtained SN-iPSCs-imDCs.These cells could be acquired in large quantities, remained in an immature state for a long time, and induced immunotolerance by inhibiting CD4+T cells and increasing CD4+CD25+FoxP3+Treg cells. The results provide a theoretical basis for the use of SN-iPSCs-imDCs in the field of transplantation immunotolerance.
ARTlCLE HlGHLlGHTS
Research background
Immature dendritic cells (imDCs) play a vital role in the induction of donor-specific transplant immune tolerance. However, these cells have limitations, such as rapid maturation and a short lifespan in vivo.The induced pluripotent stem cells (iPSCs) could differentiate to imDCs and sinomenine (SN) could inhibit the maturation of imDCs. Therefore, the iPSCs-derived imDCs treated with SN are expected to become promising seeding cells for inducing immune tolerance in organ transplantation.
Research motivation
The immune chronic rejection after solid organ transplantation is serious and still lacks an effective treatment.
Research objectives
The objective of our study was to evaluate the therapeutic effects of SN-iPSCs-imDC induced immune tolerance in vitro and allogeneic skin graft Mouse model.
Research methods
Mouse iPSCs were induced to differentiate into imDCs in a culture medium with or without SN (iPSCsimDCs and SN-iPSCs-imDCs). The related surface markers, the effects on T-cell stimulatory function,regulatory T (Treg) cell proliferative function, cytokine expression levels, cell endocytic capacity and cell apoptosis of iPSCs-imDCs and SN-iPSCs-imDCs in vitro and in vivo were analyzed. The induced immunology tolerance of SN-iPSCs-DCs was evaluated by treated the recipients Balb/c skin graft mice.Statistical evaluation of graft survival was performed using Kaplan-Meier curves.
Research results
We successfully obtained iPSCs-imDCs and SN-iPSCs-imDCs. SN-iPSCs-imDCs exhibited higher CD11c levels, lower CD80, CD86 and MHC-II levels, worse T-cell stimulatory function, and higher Treg-cells proliferative function compared with iPSCs-imDCs. Additionally, the levels of interleukin (IL)-2, IL-12,interferon-γ in SN-iPSCs-imDCs were lower than those in iPSCs-imDCs, whereas IL-10 and transforming growth factor-β levels were higher. Moreover, the cell endocytic capacity and apoptosis rate in SN-iPSCs-imDCs was significantly higher. In Balb/c mice recipients immunized with iPSCsimDCs or SN-iPSCs-imDCs 7 d before skin grafts, Tregs were significantly increased in the spleen after transplantation and the survival span of C57bl/6 skin grafts was significantly prolonged in immunized Balb/c recipients with a donor-specific pattern in SN-iPSCs-imDCs treated group.
Research conclusions
The SN-iPSCs-imDCs have potential applications for the induction of immune tolerance following organ transplantation.
Research perspectives
The SN-iPSCs-imDCs could induce immune immune hyporesponsiveness, even immune tolerance,which may be an effective strategy to treat immune chronic rejection after organ tranplantation.
FOOTNOTES
Author contributions:Huang XY, Jin ZK, Tian PX and Xu CX conceived and designed the study, and drafted the article; Dou M, Zhao XR, Feng Q, Feng YM and Duan XL performed the experiments; Huang XY and Xu CX analyzed the data; Huang XY, Jin ZK, Dou M and Xu CX performed data analysis; all authors contributed to the collection of literature data, reviewed the manuscript, and approved the version to be published.
Supported byNational Natural Science Foundation of China, No. 81900686; Science and Technology Incubation Fund Project of Shaanxi Provincial People’s Hospital, No. 2020YXM-08; Technology Talent Support Program of Shaanxi Provincial People’s Hospital, No. 2021BJ-07; Key Projects of Shaanxi Provincial Department of Education, No.21JS038; and Medical Research Development Fund of Beijing Kangmeng Charity Foundation, No. 7B202010.
lnstitutional animal care and use committee statement:All of the procedures and ethics of animal use were reviewed and approved by the Biomedical Ethics Committee of Shaanxi Provicial People’s Hospital, No. 2021-198.
Conflict-of-interest statement:All the authors report no relevant conflicts of interest for this article.
Data sharing statement:No additional data are available.
ARRlVE guidelines statement:The authors have read the ARRIVE guidelines, and the manuscript was prepared and revised according to the ARRIVE guidelines.
Open-Access:This article is an open-access article that was selected by an in-house editor and fully peer-reviewed by external reviewers. It is distributed in accordance with the Creative Commons Attribution NonCommercial (CC BYNC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited and the use is noncommercial. See: https://creativecommons.org/Licenses/by-nc/4.0/
Country/Territory of origin:China
ORClD number:Cui-Xiang Xu 0000-0001-9498-4175.
S-Editor:Fan JR
L-Editor:A
P-Editor:Yu HG
杂志排行

World Journal of Stem Cells的其它文章
- Bone marrow mesenchymal stem cell treatment improves poststroke cerebral function recovery by regulating gut microbiota in rats
- How mesenchymal stem cell cotransplantation with hematopoietic stem cells can improve engraftment in animal models
- Combination of mesenchymal stem cells and three-dimensional collagen scaffold preserves ventricular remodeling in rat myocardial infarction model
- Metabolic determinants of stemness in medulloblastoma
- Pancreatic transplant surgery and stem cell therapy:Finding the balance between therapeutic advances and ethical principles