Seasonal variations in gonad morphology and hypothalamic GnRH-I and GnIH in Eurasian Tree Sparrow,a multi-brooded passerine
2022-10-03LiminWngGhulmNiLiZhngDnLiuMoLiJuyongLiKixinShiIrhimAhmdYuefengWuJohnWingfieldDongmingLi
Limin Wng ,Ghulm Ni ,Li Zhng ,Dn Liu ,Mo Li ,Juyong Li ,Kixin Shi,Irhim M.Ahmd,Yuefeng Wu,John C.Wingfield,Dongming Li,*
a Ministry of Education Key Laboratory of Molecular and Cellular Biology,Key Laboratory of Animal Physiology,Biochemistry and Molecular Biology of Hebei Province,College of Life Sciences,Hebei Normal University,Shijiazhuang,050024,China
b Department of Neurobiology,Physiology and Behavior,University of California,Davis,CA,USA
Keywords:Eurasian Tree Sparrow Gonads Hypothalamus Reproduction Seasonality
ABSTRACT The hypothalamic-pituitary-gonadal (HPG) axis ubiquitously regulates seasonal reproduction,following the rhythmicity of a suite of environmental cues.Birds display prominent seasonal variations in gonad size regulated by two hypothalamic peptides,gonadotropin-releasing hormone-1 (GnRH-I) and gonadotropin-inhibiting hormone (GnIH).How multi-brooded avian species adjust GnRH-I and GnIH to regulate gonadal morphology seasonally remains largely unknown.Here,we studied the variations in the hypothalamic immunoreactivity (ir)of GnRH-I and GnIH,gonadal proliferation,and apoptosis in a typical multi-brooded species,the Eurasian Tree Sparrow(Passer montanus),across the pre-breeding(PB),first breeding(FB),second breeding(SB),pre-basic molt(PM),and wintering stages (WS).Our results showed that both sexes had higher preoptic area (POA)-GnRH-I-ir but lower paraventricular nucleus(PVN)-GnIH-ir neurons during the breeding stages(FB and SB)relative to other life-history stages,with no significant differences between two broods.The testes and ovaries were significantly larger during the breeding stages.Testicular volume increased during the two broods due to anincreased diameter of seminiferous tubules.Furthermore,there were more testicular apoptotic cells in PB and WS stages than in breeding stages,and in PB stage than in PM stage.Males had higher POA-GnRH-I expression than females during the breeding stages,but both sexes had comparable PVN-GnIH expression throughout the annual cycle.Both sexes of the sparrows may undergo a similar pattern of life-history stage-dependent variation in the hypothalamic GnRH-I,GnIH,and gonadal morphology,except that during breeding stages,males may display higher expression of POA-GnRH-I relative to females.The higher expression of POA-GnRH-I-ir in breeding male sparrows may be critical for male-dependent parental care.
1.Introduction
In birds,the hypothalamic-pituitary-gonadal (HPG) axis regulates gonadal steroidogenesis and gametogenesis that in turn induces the development of secondary sexual traits and facilitates reproductive behaviors (Dawson,2008;Cui et al.,2021;Ioannidis et al.,2021).Gonadotropin-releasing hormone-I (GnRH-I) promotes the release of gonadotropins from the anterior pituitary gland whereas gonadotropin-inhibiting hormone (GnIH) does the opposite.These two hypothalamic peptides regulate reproductive functions in response to energy demands and environmental signals such as the annual change in day length(photoperiod)and temperature(Deviche et al.,2011;Surbhi et al.,2015;Banerjee and Chaturvedi,2017;Dixit and Byrsat,2018;Nabi et al.,2020;Tsutsui and Ubuka,2020;Hanlon et al.,2021).Seasonal changes in gonadal size in birds are remarkable among vertebrates and include a series of neuroendocrine and biological processes regulating cell proliferation and apoptosis in the gonads(Deviche et al.,2011;Dixit and Singh,2013).Previous studies have reported the inter-relationship of the seasonal variations in gonadal morphology with GnRH-I and GnIH actions,and the involvement of the preoptic area (POA) and paraventricular nucleus (PVN) of the hypothalamus (Jenkins et al.,2007;Peixoto et al.,2012;Surbhi et al.,2015;Akhtar et al.,2020).However,unlike single-brooded avian species,multi-brooded species exhibit a unique neuroendocrinologic mechanism to orchestrate their morphological,physiological,and behavioral changes to maximize reproductive output(Li et al.,2012;Halupka and Halupka,2017).How free-living multi-brooded avian species adjust their hypothalamic GnRH-I and GnIH to regulate gonadal morphology seasonally remains largely unknown.
In birds,GnRH-I is the primary secretagogue of gonadotrophins,and thus regulates reproductive physiology (Deviche et al.,2011;Ubuka et al.,2013).The GnRH-I-ir cell bodies are bilaterally clustered in the preoptic area(POA;Surbhi et al.,2015),and their ir-fibers extend to the median eminence (ME;van Gils et al.,1993).The GnIH neurons are localized in the PVN and project to the ME,where they are in close contact with the GnRH-I(Surbhi et al.,2015;Tsutsui and Ubuka,2020).During the non-breeding stages in the Eurasian Tree Sparrows (Passer montanus),gonads regress and gonadal cells undergo apoptosis.In temperate environments with distinct seasons,day length is a reliable indicator of food availability.Therefore,seasonal environments can exert strong selective pressure on reproductive physiology,allowing birds to reproduce during peak seasonal food availability (Hurley et al.,2020).GnIH regulates the timing of seasonal reproduction by inhibiting GnRH-I activity via the GPR147 receptor (Surbhi et al.,2015;Teo et al.,2021).The gonado-inhibitory effects of GnIH suppress the synthesis and secretion of gonadotropins,leading to gonadal regression and suppression of reproductive behaviors(Bauer et al.,2018;Tsutsui and Ubuka,2021).
Among the avian species,hypothalamic GnRH-I mRNA expression varies with photoperiod,suggesting a pivotal role of GnRH-I and GnIH circuitry in regulating seasonal reproduction (Ottinger et al.,2004;Stevenson and MacDougall-Shackleton,2005;Surbhi et al.,2015).Increased hypothalamic GnRH-I and decreased GnIH expression during the breeding stage are well-documented in most avian species(Dawson et al.,2002;Stevenson et al.,2012;Dixit and Byrsat,2018).In contrast,decreased GnRH-I and increased GnIH secretion during the winter season causes gonads to mature slowly or remain in an immature state to enhance survival in harsh environments (Dawson et al.,2001;MacDougall-Shackleton et al.,2009;Kriegsfeld et al.,2015).However,lower GnRH-I levels are associated with the onset of absolute photorefractoriness after breeding in some passerines (Hahn and Ball,1995;Meddle et al.,2006),but not in some Galliformes (Teruyama and Beck,2000;MacDougall-Shackleton et al.,2009).The pre-basic molt (PM)period after breeding is also an energy demanding stage;however,the extent to which the hypothalamic GnRH-I and GnIH systems are regulated during this period is largely unknown(Dawson et al.,2001).
Testes develop maximally during the breeding season,followed by a phase of gonadal regression leading to photorefractoriness,which is governed by the hypothalamic activity of GnRH-I (Dawson et al.,2001;Dawson,2003,2008;Dixit and Singh,2013;Dixon et al.,2021).A complex molecular mechanism regulates the decrease in testicular size during regression resulting from apoptosis in Sertoli cells,germ cells,seminiferous tubules,and somatic cells mediated by GnIH(Young et al.,2001;Ubuka et al.,2006;Jenkins et al.,2007;Deviche et al.,2011;Banerjee et al.,2016;Banerjee and Chaturvedi,2017).The ovaries contain primordial,primary,secondary,and atretic follicles at different life cycle stages,but the follicular diameter and wall thickness reach their maximum limits during the breeding season (Dixit and Singh,2013).After egg-laying,the follicles regress,and the ovary becomes quiescent and diminishes in size (Dixon et al.,2021).Since egg-laying and incubation periods have higher energy costs,females may rely on supplementary environmental cues unique from males in many aspects of reproductive morphology and physiology(Ball and Ketterson,2008).To date,limited studies have been conducted on seasonal variations in the histophysiology of gonadal morphology and the corresponding regulation of hypothalamic GnRH-I and GnIH expression in female birds,especially multi-brooded species.
The Eurasian Tree Sparrow(Passer montanus)isa phenotypically plastic,multiple-brooded,and small granivorous passerine (approximate body weight:20 g)that is widely distributed across Eurasia from sea level to the Qinghai-Tibet Plateau(Sun et al.,2016;Li et al.,2020a;Ding et al.,2021).Even in highland areas,Eurasian Tree Sparrows can breed at least twice annually(Li et al.,2012,2013),which is regulatedat both the HPG levels by photoperiod and by endogenous circannual rhythmicity(Dixit and Byrsat,2018).In the multi-brooded Eurasian Tree Sparrow,meeting the increased energy requirements for both mating and parental care is fundamental for optimizingreproductive output(Lietal.,2017,2019;Nabietal.,2021).Our previous results have shown that plasma corticosteroid-binding globulin and testosterone levels are higher during the first brood than during the second brood,but testis sizes are comparable between the two (Li et al.,2012),indicating that the HPG axis is decoupled from the proliferation and apoptosis of gonads.In the present study,we studied the seasonal variations in hypothalamic GnRH-I,GnIH,and gonadal morphological features during pre-breeding (PB),first breeding (FB),second breeding (SB),pre-basic molting (PM),and wintering (WS) stages in the multi-brooded Eurasian Tree Sparrow.Our aim was to define the annual regulation of the HPG axis in both sexes and the corresponding neuroendocrinological mechanisms in a typical multi-brooded species.
2.Materials and methods
2.1.Animal collection
Free-living adult Eurasian Tree Sparrows were captured using mist nets from April to mid-October 2013 between 06:00 and 10:00 in Shijiazhuang(38°3′28′′N,114°32′47′′E),Hebei Province,China.Male and female birds were further divided into PB,FB,SB,PM,and WS stages based on the life stages determined by previous studies(Li et al.,2012).The captured tree sparrows were placed in cages (38 cm×27 cm×30 cm) and sent to the laboratory of Hebei Normal University for further sampling within 2 h.All protocols were approved by the Ethics and Animal Welfare Committee,the Institutional Animal Care and Use Committee of Hebei Normal University,and the Department of Wildlife Conservation(Forestry Bureau)of Hebei Province,China.
2.2.Gonadal size and histology analyses
Birds were weighed and then anesthetized using 2% sodium pentobarbital(3 mg/kg body weight).They were then transcardially perfused with 0.1 M phosphate buffered solution (PBS) buffer (pH 7.4) and 4% paraformaldehyde (PFA).The brains were immersed in PFA until sectioning.
After incising the abdominal wall,the length and width of the testes(n=50)and ovaries(n=70)were measured in situ.Testicular volume was calculated using the formula 4/3 πab2,whereaandbdenote the long(length) and short (width) axes,respectively (Perfito et al.,2011).The testes and ovaries were rapidly removed from male and female sparrows,respectively.One part of the testes and ovaries were immersed in Bouin's fluid,and the other part was immersed in PFA.After 3-day fixation in Bouin's fluid,testes and ovaries were dehydrated in a graded series of ethanol (50%,70%,80%,90%,95%,and 100%),treated with xylene,and embedded in paraffin wax.Sections(6 μm)were cut using a paraffin microtome (RM2125 RT,Leica,Germany) and processed for hematoxylin-eosin (HE) staining.Finally,sections were dehydrated using ascending grades of ethanol (50%,70%,80%,90%,95%,95%,100%,and 100%),cleared in xylene,and cover-slipped in neutral balsam.Histological sections of the testes and ovaries were viewed and photographed using a microscope(FV-1000,Olympus,Japan).The number of seminiferous tubules(sample size,n=29),the diameter of seminiferous tubules (n=28),and the size of the largest follicle (n=28) were determined in five sections per testis/ovary using Image-Pro Plus software(V6.0,Media Cybernetics,USA).
2.3.Apoptosis staining
The testes and ovaries were fixed in PFA for three days at 4°C and then dehydrated in 15% and 30% sucrose solutions.Serial transverse slices of the testes and ovaries were cut at 30 μm intervals using a freeze microtome (CM-1850,Leica,Wetzlar,Germany).The slices were mounted about gelatin-coated glass slides and stored at -20°C.The sections were dried for 20 min and incubated with 0.3%H2O2.After three rinses with PBS,sections were incubated with terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL) reaction mixture (TUNEL Assay kit,Roche,Switzerland)in the dark for 1 h at 37°C.After rinsing thrice with PBS buffer(0.1 M,pH 7.4),sections were incubated with 4′,6-diamidine-2′-phenylindole dihydrochloride (DAPI).All sections were photographed using a laser confocal microscope (FV-1000,Olympus,Shinjuku,Japan).The number of TUNEL-immunoreactive (TUNEL-ir)cells in the testis(n=25)was measured bilaterally using Image-Pro Plus software(V 6.0,Media Cybernetics,MD,USA).
2.4.Immunohistochemical analysis of GnRH-1 and GnIH
Brains were removed from male and female sparrows,fixed in PFA for three days,dehydrated in 15%and 30%sucrose solution,and then serial transverse slices were cut at 30 μm intervals using a freeze microtome(CM-1850,Leica,Germany).The slices were mounted about gelatincoated glass slides in sequence and stored at -20°C.Sections were dried and immersed in 0.1 M PBS buffer and then incubated in 0.3%H2O2dissolved in methanol for 20 min under dark conditions.After washing 3 times in PBS,the sections were blocked in 5% bovine serum albumin (BSA) blocking solution (containing 0.2%Triton X-100) for 30 min.For GnRH-I(male:n=21;female:n=21)and GnIH(male:n=25;female:n=25)immunohistochemical staining,sections were incubated with primary antibodies,either rabbit anti-GnRH(1:3000)or rabbit anti-GnIH (1:1000)at 4°C overnight.The primary antibody for GnRH-I and GnIH were made by a polyclonal rabbit antiserum (anti-Eurasian Tree Sparrow GnRH-I or GnIH)in the Institute of Biology,Hebei Academy of Sciences,Shijiazhuang,China,according to the translation of nucleotide sequences of Eurasian Tree Sparrows (Nabi et al.,2020).The staining specificities of GnRH-I and GnIH antibodies were assessed by substitutions of the control serum for the antiserum.The next day,three washes in PBS were followed by incubation for 2 h in a biotinylated goat anti-rabbit IgG secondary antibody(1:200,Boster,Wuhan,China).After washing,the sections were incubated for 1 h in an avidin-biotin complex(SABC-AP Kit;Boster,Wuhan,China).Finally,all sections were washed with PBS and treated with diaminobenzidine solution in dark (DAB,Boster,Wuhan,China).The sections were immediately transferred into dH2O to stop the reaction after obvious color was observed.The sections were washed in distilled water and dehydrated using ascending grades of alcohol(50%,70%,80%,90%,95%,95%,100%,and 100%),cleared in xylene,and cover-slipped in neutral balsam.All sections were observed and photographed using an optical microscope (FV-1000,Olympus,Shinjuku,Japan).The number of GnRH-I-immunoreactive (GnRH-ir)cells in the POA and GnIH-immunoreactive (GnIH-ir) cells in the PVN were measured bilaterally using the “cell counting” function of Image-Pro Plus software(V6.0,Media Cybernetics,MD,USA).
2.5.Statistical analysis
All data were checked for normality using a one-sample Kolmogorov-Smirnov test.One-way analysis of variance (ANOVA) was used to investigate variations in gonadal histomorphology of males or females across different life-history stages,and two-way ANOVA was used to investigate variations in body weight and the number of GnRH-ir and GnIH-ir cells by considering life-history stage and sex as fixed factors.In addition,an independent-samplest-test was used to investigate the largest follicle diameter between the two breeding stages.Post hoctests between groups were performed using Tukey's test.The figures were generated using GraphPad Prism v8.0 (GraphPad Software Inc.,CA,USA).All statistical procedures were performed using SPSS V 20.0(SPSS Inc.,Chicago,USA).Differences were considered significant atP<0.05.All data are presented as the mean±SEM.
3.Results
3.1.Variations in body weight across different life-history stages
There were significant differences in body weight between the sexes and among life-history stages,and a significant interaction existed between sex and life-history stage(Table 1).Post hocanalysis revealed that male Eurasian Tree Sparrows were significantly heavier in the PM and WS,followed by the PB and FB relative to that in the SB(Fig.1;Appendix Table S1).However,female sparrows did not exhibit significant changes in body weight across different life-history stages (Fig.1;Appendix Table S2).In addition,male sparrows were significantly heavier than females in the PM (P<0.001) and WS (P<0.001),but significantly lighter than females in the SB(P=0.023)stage(Fig.1).
3.2.Testicular structure and apoptosis across the life-history stages
The testes of Eurasian Tree Sparrows appeared rounded to ovalshaped (Fig.2A),and testicular volume and the number and diameter of the seminiferous tubules varied significantly with the life-history stage(Table 1).Post hocresults showed that testicular volume was significantly larger in the PB,FB,and SB stages than in the PM and WS stages.There were no significant differences in testicular volume between the two breeding stages(FB and SB),but that in FB was significantly larger than that in PB (Fig.2B;Appendix Table S1).The number of seminiferous tubules was significantly lower in the PB,FB,and SB stages than in the WS stage,and in the FB and SB stages relative to the PM stage(Fig.2C;Appendix Table S1).In contrast,the diameter of the seminiferous tubules was greater at the PB,FB,and SB stages than at the PM and WS stages(Fig.2D;Appendix Table S1).
In the testes,the TUNEL-positive apoptotic cells were predominantly present in the lumen and interstitial space (Fig.2E).The number of testicular apoptotic cells varied significantly with life-history stages(Table 1).Post hocresults showed that there were significantly more apoptotic cells in the PB and WS stages than in the FB and SB stages,and in the PB stage relative to the PM stage,although there were no significant differences between the PB and WS stages(Fig.2F).
3.3.Ovarian structure and apoptosis across the different life-history stages
The size and structure of the ovary varied significantly with the lifehistory stages (Table 1;Fig.3A).Both primary and secondary follicles were found in the breeding(FB and SB)stages,but only primary follicles were present in the non-breeding (PB,PM,and WS) stages.Post hocresults showed that the length and width of the ovary were significantlyhigher at the FB and SB stages,followed by the PB stage relative to those at the PM and WS stages(Fig.3B and C;Appendix Table S2).There were no significant differences in the length and width of the ovaries between the FB and SB stages or between the PM and WS stages.There were no significant differences in the diameter of the largest follicles between the FB and SB stages(Fig.3D).
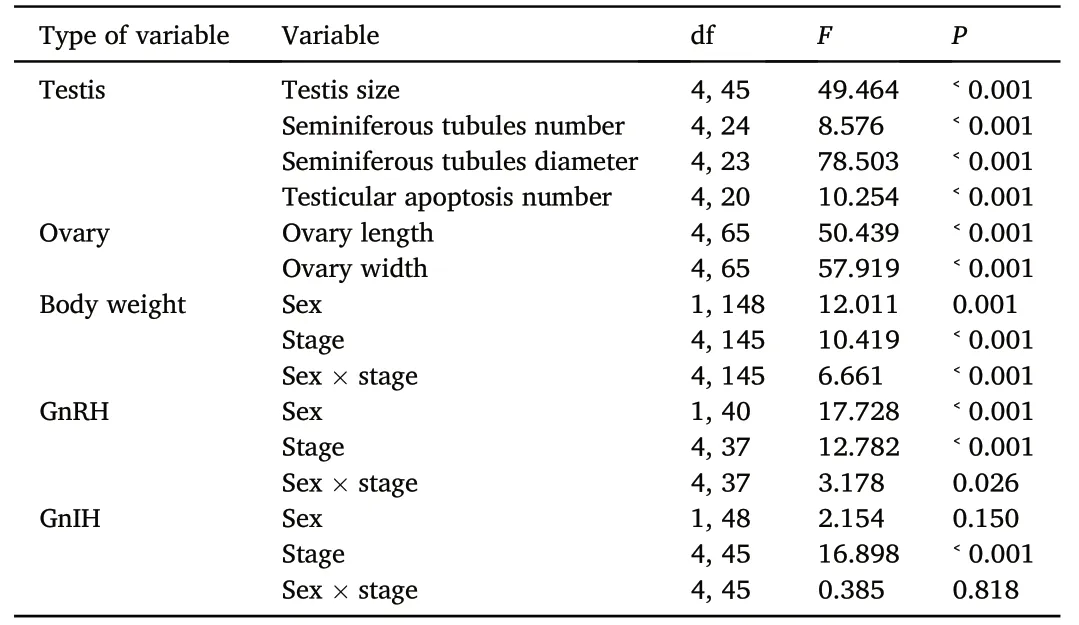
Table 1 The statistical results of testis and ovary morphology across different life-history stages in a one-way ANOVA,and body weight,GnRH,and GnIH in a two-way ANOVA by considering sex,life-history stage (stage),and interaction of species and life history stage as fixed factors.
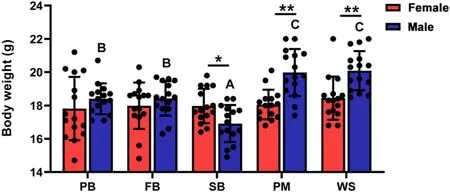
Fig.1.Life-history stage-dependent variations in body weights of male and female Eurasian Tree Sparrows (Passer montanus).PB,pre-breeding stage;FB,first breeding stage;SB,second breeding stage;PM,pre-basic molt stage;WS,wintering stage.Data are shown as mean ± SEM.Groups with different letters show significant differences from other groups (P<0.05),and asterisk (*) indicates significant differences between sexes at a certain life-history stage (P<0.05).
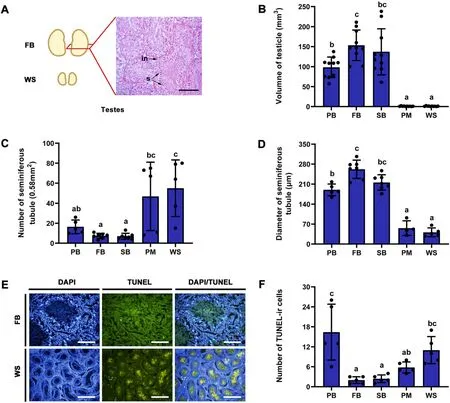
Fig.2.Testicular structure and apoptosis across different life-history stages of male Eurasian Tree Sparrows (Passer montanus).Testes in FB and WS,and transverse section of testis in FB stage showing the interstitial tissue (in) and spermatogonia (s) (A);changes in testicular volume (B),seminiferous tubule number (C),and seminiferous tubule diameter(D)across different life-history stages;TUNEL-ir cells present in the lumen and interstitial space,and there are significantly more cells in the WS relative to the FB stages(E);changes of TUNEL-ir cell number across different life-history stages(F).PB,pre-breeding stage;FB,first breeding stage;SB,second breeding stage;PM,pre-basic molt stage;WS,wintering stage.Bar=200 μm.Data are shown as mean ± SEM.Groups with different letters show significant differences from other groups (P <0.05).
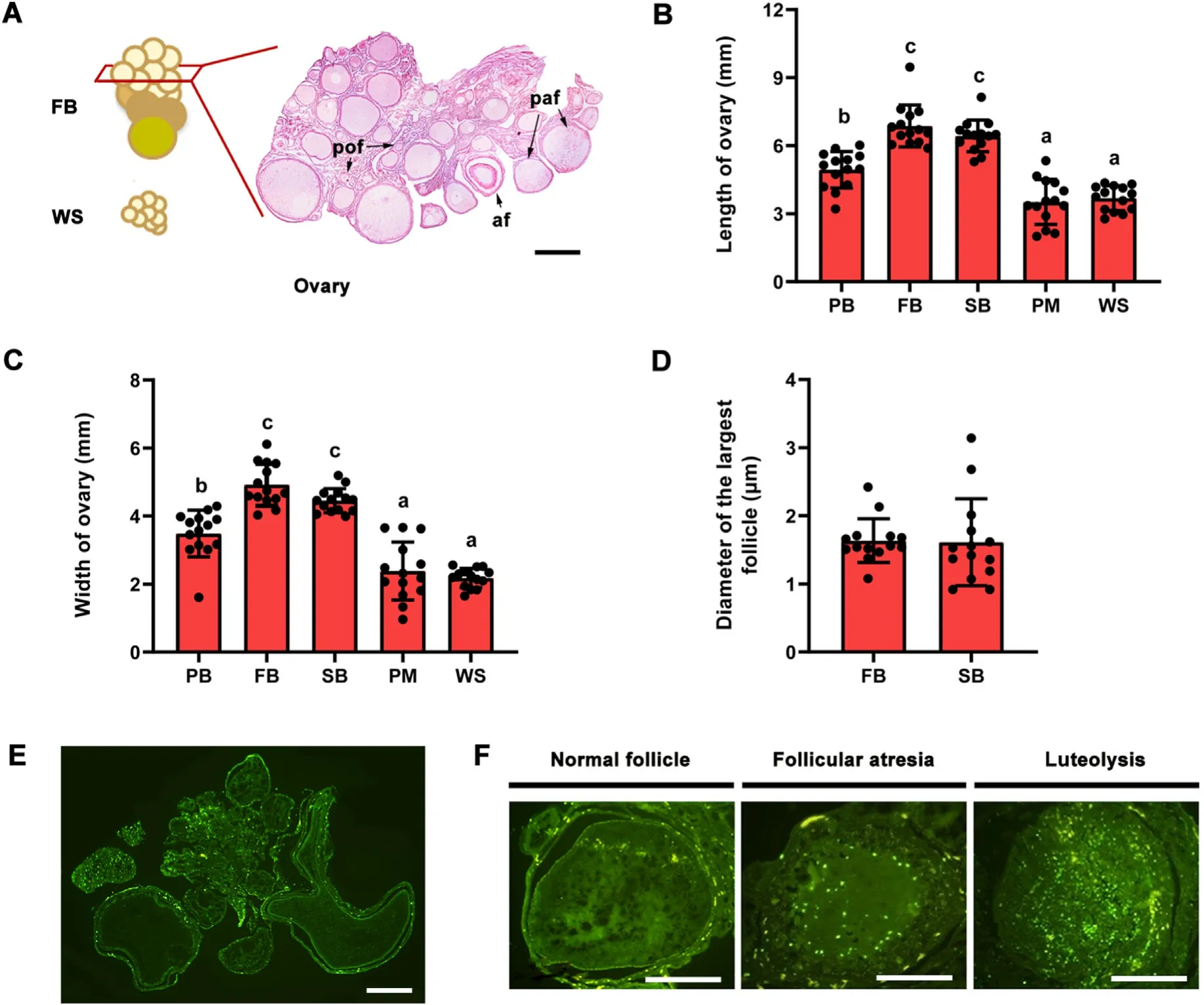
Fig.3.The ovarian structure across different life-history stages of female Eurasian Tree Sparrows(Passer montanus).Ovary in FB and WS,and transverse section of the ovary in FB showing primordial follicles(pof),primary follicles(paf),and atretic follicles(af)(A);changes of ovary length(B)and ovary width(C)across different lifehistory stages;comparison of largest follicle diameter between FB and SB stages (D);follicle apoptosis during FB (E);apoptotic cells in the normal follicle,follicular atresia,and luteolysis (F).PB,pre-breeding stage;FB,first breeding stage;SB,second breeding stage;PM,pre-basic molt stage;WS,wintering stage.Bar=200 μm.Data are shown as mean ± SEM.Groups with different letters are significantly different from one another stage (P<0.05).
In the ovary,the TUNEL-positive apoptotic cells were mainly present in the follicular granulosa cells (Fig.3E).Follicular atresia of the ovary occurred in all life-history stages,and the apoptotic cells in follicular atresia and luteolysis were significantly more obvious than those in normal follicles.(Fig.3F).
3.4.Expression of GnRH-I in the POA and GnIH in the PVN across the lifehistory stages
There were significant differences in number of GnRH-I-ir cells in the POA between the sexes and among life-history stages;a significant interaction existed between sex and life-history stage (Table 1;Fig.4A,C).Post hocanalysis revealed that both male and female sparrows had significantly higher number of GnRH-I-ir cells in the POA during the breeding (FB and SB) stages than the PM and WS stages (Fig.4C;Appendix Tables S1 and S2).Furthermore,neither sex exhibited a significant difference between PB and other stages.Although there were no significant differences between male and female sparrows in the nonbreeding stages (PB,P=0.065;PM,P=1.000;SW,P=0.807),male sparrows had significantly higher number of GnRH-I-ir cells in the POA than females in the breeding (FB,P=0.003;SB,P<0.001) stages(Fig.4C).
There were significant differences in number of GnIH-ir cells in the PVN between the sexes and among life-history stages,and no significant interaction between sex and life-history stage(Table 1;Fig.4B,D).Post hocanalysis revealed that males had significantly lower number of GnIHir cells in the PVN during the FB and SB stages relative to the PB stage,and lower number during the SB relative to the non-breeding (PM,WS,and PB) stages (Fig.4D;Appendix Table S1),although there was no significant difference between the FB and SB stages.Females had significantly lower number of GnIH-ir cells in the PVN during the breeding(FB and SB)stages than in the non-breeding(PM,WS,and PB)stages (Fig.4D;Appendix Table S2).There were no significant differences between male and female sparrows in both breeding (FB,P=0.443;SB,P=0.054)and non-breeding(PB,P=0.295;PM,P=0.896;WS,P=0.956)stages(Fig.4D).
4.Discussion
4.1.Seasonal variations of body condition and gonadal morphology
Seasonally breeding birds exhibit well-defined seasonality in their reproductive functions (Dawson,2007).In the multi-brooded Eurasian Tree Sparrows,we found that body condition,gonadal structure,and size varied significantly with the life-history stage.The testes and ovaries were significantly larger during the breeding (first and second) stages than during other life stages,which is in agreement with previous results(Li et al.,2012;Dixit and Singh,2013;Nabi et al.,2020).In the male sparrows,our results are consistent with previous findings showing that testicular size,spermatogenesis,and seminiferous tubule diameter are positively correlated in birds(Dixit and Singh,2013;Akhtar et al.,2020).In addition,the increase in testicular volume during the breeding season is an outcome of increasing the diameter but decreasing the number of seminiferous tubules.This result conforms with the observations in other birds,such as the White-eyed Parakeet (Aratinga leucophthalma;Peixoto et al.,2012) and the Jungle Babbler (Turdoides striatus;Bhavna and Geeta,2010).In addition,we found that the male Eurasian Tree Sparrows exhibited significantly lower body weight during the breeding than other non-breeding stages;this differs from the females that had no variations across life-history stages.Testes in birds can develop up to 480-fold during the breeding season due to increased diameter of the seminiferous tubules,epithelial height,and lumen diameter:this developmenthelp to cope with the high copulatory and spermatogenic demands (Vézina and Salvante,2010;Akhtar et al.,2020).A similar pattern of development in testicular size during the breeding stage accompanied by higher metabolic costs were reported in Eurasian Tree Sparrows (Nabi et al.,2020),Jungle Babbler(Bhavna and Geeta,2010),White-eyed Parakeets(Peixoto et al.,2012),Japanese Quail (Coturnix coturnix japonica;Shil et al.,2015),and Yangzhou Goose(Anser domesticus;Akhtar et al.,2020).In Eurasian Tree Sparrows,testicular physiology,spermatogenesis,mating,and other reproductive behaviors are highly associated with increased metabolism induced by plasma triiodothyronine (T3) during the breeding stage: body weight,relative to other stages,decreases significantly(Li et al.,2020a;Nabi et al.,2020).
Our results indicate that testicular apoptotic cells increase significantly outside the breeding stages.In the majority of avian species studied to date,photoperiod is an important environmental factor regulating seasonal reproduction.During their annual gonadal cycle,birds sensitive to either short-or long-day effects develop refractoriness to the same day length and gonads develop or regress accordingly(Banerjee and Chaturvedi,2017).To achieve precise testicular function,the renewal,proliferation,and apoptosis of each germ cell type in the active testis must be finely balanced(Print and Loveland,2000;Jenkins et al.,2007;Otsuka et al.,2010).These highly regulated sequential steps of spermatogenesis occur at the cost of substantial germ cell loss.It also appears that apoptosis might be responsible for testis regression in seasonally breeding species (Young and Nelson,2001;Heninger et al.,2004).During the non-breeding stages,apoptotic bodies form when cell membranes disintegrate and are then phagocytosed by macrophages,leading to testicular regression.This regression may be an adaptation for reducing the costs of flight during the non-breeding season (Dawson et al.,2001;Young et al.,2001;Banerjee and Chaturvedi,2017).During the non-breeding period when estradiol (E2/Erα) levels are lower,extracellular signal-regulated kinases(ERK1/2)and the serine/threonine kinase (Akt-1) pathways are inactivated;these inactivation increase the levels of Bax and cytochrome C,and thus favors testicular apoptotic events(Falvo et al.,2021).We failed to detect any significant differences in testicular size and apoptotic cells between the first and second broods.Previous studies have shown that Eurasian Tree Sparrows have comparable testicular volume but lower plasma testesterone levels during the second broods relative to their first(Li et al.,2012).Our results indicate that plasma testesterone levels may not maintain testicular structure and function.
Ovarian size increases significantly during the breeding stage and regresses during the post-breeding stage (Hurley et al.,2020).In the female Eurasian Tree Sparrows,we found that the size and structure of the ovary varied with life-history stage.Both primary and secondary follicles were found in the breeding stages(FB and SB),and only primary follicles were found in the non-breeding stages.Moreover,the primary follicular atresia of the Eurasian Tree Sparrow occurs throughout the entire life cycle,while secondary follicular atresia only occurs during the entire breeding period.Similar to our results,other passerine birds also exhibit well-developed ovaries,with the largest follicle increasing by up to 60-fold during the breeding season (Dixit and Singh,2014).These results are consistent with those of other studies in the Northern hemisphere and temperate-zone birds(Jacobs and Wingfield,2000;Williams and Ames,2004;Dawson,2008;Hurley et al.,2008).
Egg production is costly and has several energy constraints: it compromises flight,depletes protein,and causes immunosuppression (Williams,2005).Birds can adjust the energy demands of the reproductive organs during different breeding sub-stages via numerous energy trade-offs between reproduction and emergent life-history stages(Vézina and Williams,2003;Wingfield,2008;Follett,2015).Therefore,an increase in ovarian size during the breeding stages is essential for meeting the physiological and energy requirements of egg formation (Hurley et al.,2020).When birds are at the onset of other life-history stages such as pre-basic molt and wintering periods,they switch off reproduction and allocate energy to other events(Wingfield,2008;Follett,2015).
It is widely accepted that the circannual recrudescence and regression cycle observed in seasonally breeding species reflect the energy and physiological costs of egg production and the maintenance of reproductive tract tissues.The idea that such costs influence reproductive function is further supported by seasonally breeding birds that only engage in one energy-intensive life stage at a time (Wingfield,2008;Follett,2015).Furthermore,a number of seasonally breeding species shifts their metabolic rates to their reproductive state,for example,Great Tit (Parus major) (Nilsson and Råberg,2001) and European Starlings (Sturnus vulgaris)(Vézina and Williams,2002).
Apart from the breeding stages,both the PM and WS are also energyintensive life-history stages(Buttemer et al.,2020;Li et al.,2020b;Nabi et al.,2021).Feathers represent approximately 25% of a bird's protein content and impose huge nutrient and energy demands during self-maintenance to carry out diverse critical physiological functions,including feather replacement (molt),thermoregulation,communication,protection,and enhancement of aerodynamic performance (Buttemer et al.,2020;Pageau et al.,2021).When food availability is limited during the wintering stage,small-sized passerines must enhance their metabolic capacity for powered flight and sustained aerobic thermogenesis(Li et al.,2020a,b;Nabi et al.,2021).The reduced gonadal sizes we detected in both sexes of the Eurasian Tree Sparrows during PM and WS stages reflect a strategy for tightly scheduling these life-history stages,linking trade-offs among breeding,self-maintenance,and survival(Wolfe et al.,2021).
4.2.Seasonal variations of hypothalamic GnRH-I and GnIH
We found that both male and female Eurasian Tree Sparrows showed significantly higher POA GnRH-I-ir but lower PVN GnIH-ir cells during the two breeding sub-stages relative to other life-history stages,with no significant differences between the first and second breeding stages.GnIH expression is negatively correlated with circulating sex steroids,growth of seminiferous tubules,and proliferation of quail germ cells,suggesting that it is the main regulator of testicular development and function (Ubuka et al.,2006).At the peripheral level,in mature male birds,extensive studies have demonstrated that chronic GnIH treatment decreases plasma testosterone concentration,induces apoptosis of testicular cells,and decreases the diameter of seminiferous tubules(Ubuka et al.,2006;Bentley et al.,2008;McGuire and Bentley,2010;Rizwan et al.,2012).In immature male birds,GnIH treatment suppresses testicular growth of the testis (Ubuka et al.,2006).These findings indicate that as a key hypothalamic neurohormone,GnIH suppresses the development and maintenance of gonads by decreasing the synthesis and release of gonadotropin in birds(Ubuka et al.,2006;Tsutsui and Ukena,2006;Tsutsui,2009,2016;Tsutsui et al.,2010a,2010b,2015).
In seasonally breeding birds,GnRH-ir cells reach a peak during the breeding season,and reach a minimum after the onset of photorefractoriness,while day lengths are also becoming shorter in many species(Stevenson et al.,2009;Ubuka and Bentley,2009).Similar to our results,larger gonads in Eurasian tree sparrows during the mating season are stimulated by higher hypothalamic GnRH-I and lower GnIH levels(Stevenson et al.,2012).Increased GnRH-I expression,during the breeding season,promote sex steroid secretion,gonadal maturation,and ultimately,sexual behavior when food availability,and other environmental conditions,are favorable(Dawson,2008;Sibly et al.,2012;Nabi et al.,2020).Higher GnIH acts on GnRH-I neurons suppressing the synthesis and secretion of GnRH-I,gonadotropins,and gonadal steroids,and ultimately terminates reproductive behavior(Ubuka et al.,2006;Tsutsui et al.,2012;Surbhi et al.,2015;Bauer et al.,2018)to allow reallocation of energy for maintenance and survival.
Furthermore,we found that during the non-breeding stages(PM and PS),the number of POA-GnRH-I decreased while PVN-GnIH cells increased along with regressed gonads.These changes signify that the reproductive mode is switched off as the activity of GnRH-I decreases while hypothalamic GnIH expression increases post-breeding (Ubuka et al.,2006;Stevenson et al.,2012;Tsutsui et al.,2012).Therefore,increased GnIH during post-breeding has gonado-inhibitory effects -leading to gonadal regression and suppression of reproductive behaviors,both directly and indirectly (Stevenson et al.,2012;Bauer et al.,2018).
During the pre-breeding stage,both gonadal size and POA-GnRH-I and PVN-GnIH expressions were comparable to those in the other stages.At the end of the photorefractory period,the number of GnRH-I cells increased significantly in the POA and then in the ME (Dawson and Goldsmith,1997),and ultimately,a rapid increase in gonadotropin levels in gonadectomized birds reconnects the brain and reproductive endocrine systems (Dawson and Goldsmith,1984).However,it was an unexpected discovery that both sexes had a comparable PVN-GnIH expression throughout the annual cycle,but males had significantly higher POA-GnRH-I expression than females during the breeding stages.To the best of our knowledge,this is the first report to identify a sex-specific difference in GnRH-I,but not in GnIH expression,controlling the activities of the downstream HPG axis in free-living animals during the breeding season.
Previous studies have shown that the number of brain GnRH-I-ir cells in songbirds was positively correlated with circulating testesterone levels during breeding stage (Amorin and Calisi,2015),and was highly associated with sex-dependent abilities in learning and memory,and stress responsiveness(McGuire et al.,2013).Furthermore,a recent study found that exogenous GnRH treatment in male Tree Swallows (Tachycineta bicolor) resulted in more frequent provisioning during parental care compared to untreated males,suggesting that increased GnRH male birds during breeding is essential for paternal effort (George et al.,2021).Considering that little information is available to explain why breeding male sparrows remarkably increased the expression of GnRH-I-ir cells,the underlying regulatory mechanisms of such sex-specific differences need to be further investigated.
5.Conclusions
In the present study,we identified dynamic changes in body weight,gonadal structure,size,and the hypothalamic GnRH-I-ir and GnIH-ir neurons in a typical multi-brooded passerine,the Eurasian Tree Sparrow.First,our results confirm that temperate-zone species exhibit season-dependent changes of increased gonadal volume and GnRH-I neurons but decreased GnIH neurons.These results identify no significant differences in the measured reproduction-related parameters between the first and second broods.Second,our results provide the changes in gonadal histomorphology across breeding substages in both sexes of this multi-brooded species.Finally,it is notable that the male sparrows had higher POA-GnRH-I expression than females during the breeding stages,but both sexes had a comparable PVN-GnIH expression at a specific stage throughout the annual cycle.Our results contribute to a better understanding of how the GnIH-GnRH-I neural circuitry is regulated in a multi-brooded passerine;and suggest that a poorly understood sex-dependent regulatory mechanism of the HPG axis might occur in the breeding season.Whether such increased expression of POA-GnRH-I-ir in breeding male sparrows is critical for male-dependent parental care,and how exactly this neural circuitry is regulated in both sexes in response to changing photoperiod and other environmental cues,emergent life stages,and other energy-intensive events,merit further investigation.
Ethics statement
This study was conducted in accordance with the Institutional Committee for Animal Care and Use of Hebei Normal University,China,and was carried out under the auspices of scientific collecting permits issued by the Departments of Wildlife Conservation(Forestry Bureau)of Hebei Province,China.
Author contributions
LW:Formal analysis,Writing original draft,Writing review&editing,Funding acquisition.
GN:Formal analysis,Writing original draft,Writing review&editing.
LZ: Investigation,Methodology.
D.Liu:Investigation,Methodology.
ML:Investigation,Methodology.
JL: Investigation,Methodology.
KS: Investigation,Methodology.
IMA:Writing original draft,Writing review&editing.
YW:Conceptualization,Supervision.
JCW:Writing original draft,Writing review&editing.
D.Li: Conceptualization,Supervision,Funding acquisition,Writing review&editing.
Declaration of competing interest
The authors declare that they have no competing interest.
Acknowledgments
We thank Shuo Cui,Yinchao Hao,Xuelu Liu,Yaqing Li,Chenyang Hao for their assistance with sample collection in the field.This work was supported by the National Natural Science Foundation of China (NSFC,31971413) to DL and NSFC (31770445) to YW,the Natural Science Foundation of Hebei Province,China(NSFHB,C2020205038)to DL,and NSFHB (C2020205005),and the Postdoctoral Research Foundation of China(PRFC,2020M670685)to LW.
Abbreviations
HPG Hypothalamic-pituitary-gonadal
GnRH-I gonadotropin-releasing hormone-I
GnIH gonadotropin inhibiting hormone
PB pre-breeding
FB first breeding
SB second breeding
PM pre-basic molt
WS wintering stages
POA preoptic area
PVN paraventricular nucleus
ME median eminence
CBG corticosteroid-binding globulin
T testosterone
PBS phosphate buffered solution
PFA paraformaldehyde
TUNEL terminal deoxynucleotidyl transferase dUTP nick end labeling
DAPI 4',6-diamidine-2′-phenylindole dihydrochloride;
BSA bovine serum albumin
DAB diaminobenzidine solution
Appendix A.Supplementary data
Supplementary data to this article can be found online at https://do i.org/10.1016/j.avrs.2022.100037.
杂志排行

Avian Research的其它文章
- Functional and phylogenetic structures of pheasants in China
- Thermoregulatory function and sexual dimorphism of the throat sack in Helmeted Guineafowl (Numida meleagris) across Africa
- Distribution pattern and driving factors of genetic diversity of passerine birds in the Mountains of Southwest China
- Multiple lines of evidence confirm that the critically endangered Blue-crowned Laughingthrush(Garrulax courtoisi)is an independent species
- Corrigendum to “Multiple lines of evidence confirm that the critically endangered Blue-crowned Laughingthrush (Garrulax courtoisi) is an independent species” [Avian Res.13 (2022) 100022]
- Altitudinal seasonality as a potential driver of morphological diversification in rear-edge bird populations