Altitudinal seasonality as a potential driver of morphological diversification in rear-edge bird populations
2022-10-03JosLuisTellerIreneHernndezTellezIvdeHerJosIgnioAguirreAlejndroOnrui
José Luis Tellerí ,Irene Hernández-Tellez ,Iván de l Her ,José Ignio Aguirre ,Alejndro Onrui
a Department of Biodiversity,Ecology and Evolution,Universidad Complutense,28040,Madrid,Spain
b Department of Integrative Biology,Oklahoma State University,Stillwater,OK,USA
c Fundación Migres,Tarifa,11380,Cádiz,Spain
Keywords:Altitudinal movements Flight apparatus Population diversification Snow cover effects
ABSTRACT Populations at the low latitude limits of a species range (rear-edge populations) are often considered more vulnerable to climate change.However,their ability to track different environmental settings at a regional scale has been widely overlooked,although this may be relevant to accurately assess their adaptive capacity to cope with ongoing changes.Here we tested whether the endemic African Chaffinch (Fringilla coelebs africana) tracks environmental changes(e.g.decreasing temperatures,snow cover)by rearranging their numbers between seasons(spring vs.winter)along the altitude gradients of its northwestern African range.We additionally tested whether these seasonal changes in abundance were paralleled by morphological variation,suggesting a process of population diversification.We assessed African Chaffinch abundance in tree covered farmland and woodland sites distributed along an altitude gradient in spring and winter.In addition,we captured and measured chaffinches within the study gradient to explore the patterns of morphological variation.Our results showed that chaffinches shifted to lowlands from snow covered highlands during winter.In addition,highland individuals showed longer and more concave wings than their lowland counterparts.These morphological traits are usually related to flight efficiency in migratory birds,which suggest the presence of altitudinal movements aimed to track the environmental seasonality caused by orography.These results suggest a potential role of altitudinal seasonality as a driver of regional diversification within the African Chaffinch populations,which could be occurring in other North African avian species given their relatively high endemicity in the region.The evolutionary and conservation implications of these displacements have been often overlooked despite they can shape the adaptive capacity of rear-edge bird populations to face the ongoing environmental changes in this peripheral area of the Palearctic.
1.Introduction
In a context of global warming and poleward shift of species distributions,it has been suggested that populations living at the low latitude limits of a species range(rear-edge populations)would be more likely to become extinct(Hampe and Petit,2005).In this geographical setting,the deterioration of environmental conditions due to climate change will particularly undermine the habitat quality in these peripheral locations,leading to the extinction of the populations they harbor.However,this assumption often overlooks some particularities affecting the sensitivity to changes of these peripheral populations(Vilà-Cabrera et al.,2019).For instance,many rear edge populations are taxonomically different from the core populations,suggesting reproductive isolation and putative adaptations to regional conditions (Pironon et al.,2017;Angert et al.,2020).Thus,some intrinsic characteristics of rear-edge populations can be useful to predict their responses to environmental change,which raises the interest of exploring the ecological drivers promoting their regional diversification within their restricted geographical setting(Williams et al.,2008).In this context,investigating trait variation along environmental gradients could reveal mechanisms critical for understanding the adaptive capacity of rear-edge populations.
The ability to track the current spatial-temporal changes in habitat suitability might be an advantageous trait to overcome future environmental challenges (Robinson et al.,2009;Travis et al.,2013).In this context,birds display a broad range of dispersion strategies(Sheard et al.,2020),including migratory movements which are evident between species,but can also occur among populations of the same species,or between individuals of the same population (e.g.partial-migratory species;Dingle and Drake,2007;Chapman et al.,2011).In temperate regions of the northern hemisphere,a common pattern in widely distributed birds is the reduction of the migratory intensity and/or prevalence towards the lowest latitude of their range,which suggests a sedentary life style in the most southern populations(Newton and Dale,1996).However,in the presence of environmental heterogeneity caused by mountain ranges,these southern populations could perform seasonal altitudinal displacements,whose biological and conservation implications have often been overlooked(Boyle et al.,2016;Boyle,2017;Hsiung et al.,2018;Williamson and Witt,2021).For instance,because of the similar environmental seasonality of latitudinal and altitudinal gradients,it can be hypothesized that individuals performing altitudinal movements would differ from their sedentary counterparts in some phenotypic traits associated with flight economy(Tellería et al.,2001;Lundblad and Conway,2019).Thus,the spatial-temporal patterning of habitat suitability related to orography could be a main driver of population diversification at local scales,which could improve the potential of species to face environmental changes(Richardson et al.,2014).
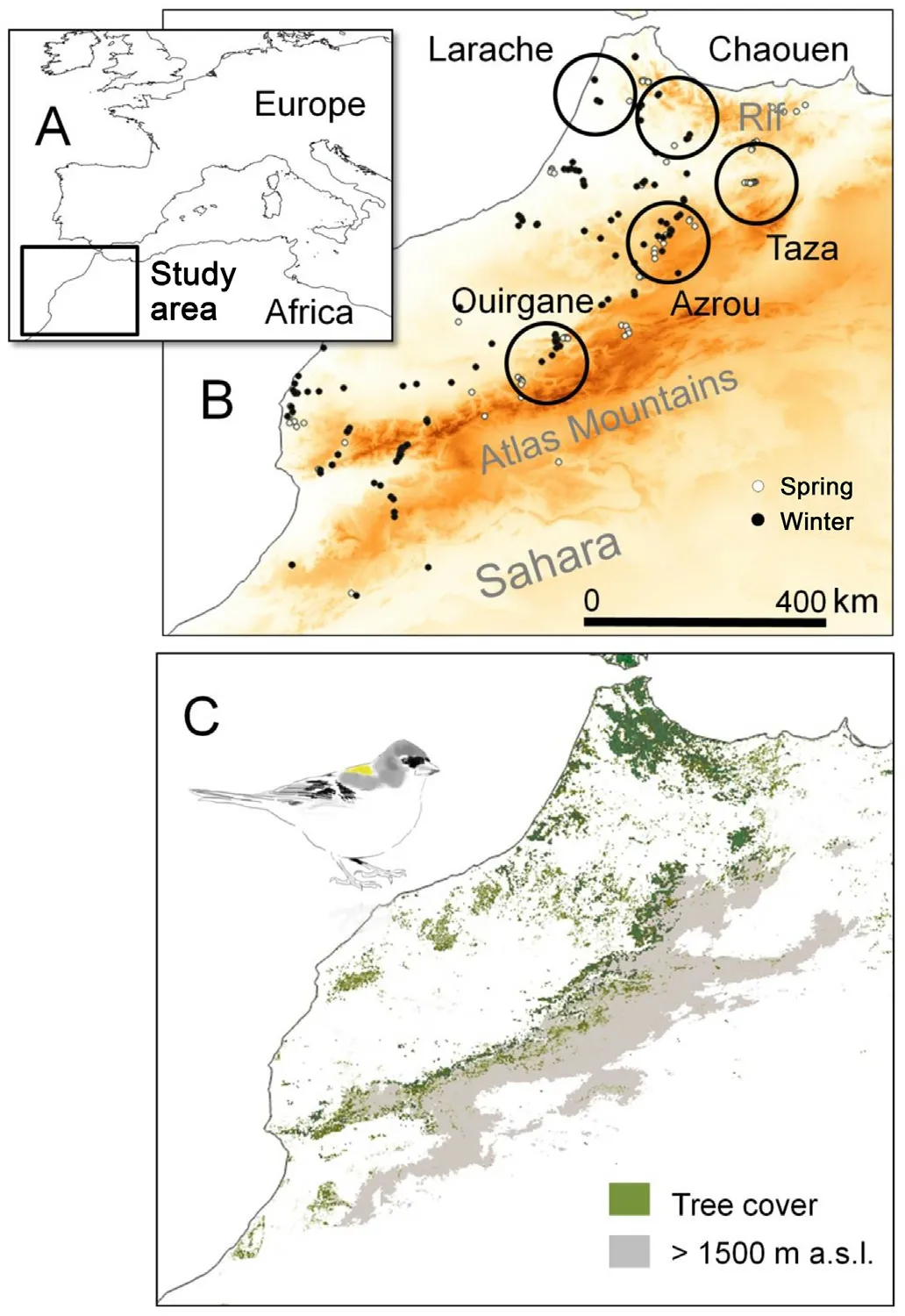
Fig.1.Situation of the study area within the western Palaearctic (A) and location of line transects carried out in spring and winter(white and black small dots respectively) and the areas (circles) where birds have been captured to explore wing morphology (large circles) (B).Tree cover distribution,the main habitat of the African Chaffinch,is represented in hues of green.The area over 1500 m a.s.l.,where most snowfalls occur in winter,is represented in grey (C).
Northwestern Africa is an area increasingly affected by drought as a consequence of raising temperatures (Bucchignani et al.,2018),which can jeopardize the long-term persistence of many species (Hampe and Petit,2005).The region is covered by high plateaus and mountains(Fig.1) occupied by rear-edge populations of Palearctic forest birds(Merker and Chandler,2020) whose taxonomic differentiation (Isenmann and Thévenot,2018)could result from reproductive isolation and putative adaptations to regional conditions (García-Ramos and Kirkpatrick,1997;Kawecki,2008).Here we explore the altitudinal movements of the endemic African Chaffinch (Fringilla coelebssubsp.africanaJ.Levaillant,1850;Griswold and Baker,2002;Perktas¸ et al.,2017) in Morocco,which is distributed along a wide environmental gradient in northwestern Africa (Thévenot et al.,2003;Tellería et al.,2021).This population is reputed as sedentary since it is observed within its breeding range all year round(Thévenot et al.,2003).However,this distribution does not preclude the existence of seasonal movements of chaffinches from the most elevated areas that,together with a morphological differentiation to improve flight efficiency,could support independent dynamics of diversification shaped by orography (Tellería et al.,2001).In this study,we tested this hypothesis by exploring the putative link of altitude to seasonal changes in abundance and morphology according to the following predictions:
(1) Seasonal changes in abundance.Birds can be compelled to leave highlands seasonally by the effect of some particular weather events,such as the direct or indirect effect of low temperatures.For instance,ground feeding birds will leave snow-covered areas when the access to ground food collapses(Carrascal et al.,1987,2012).Here we tested whether the abundance of the African Chaffinch reduces along an altitude gradient and whether this decrease is especially sharp over an altitude threshold in which snowfalls might restrict the access to ground food on which this species relies (Cramp and Perrins,1994).
(2) Morphological variation.We tested whether the external flight apparatus of the African Chaffinch varied with altitude.Since wing configuration can be related to flight efficiency,which affects migration and dispersal in birds(Winkler and Leisler,1992;Hedenström,2008;Vágási et al.,2016;Sheard et al.,2020;Claramunt,2021),we predict that highland chaffinches will show longer and more concave,or pointed,wings than lowland counterparts,which would support the existence of altitudinal movements within their range.
2.Material and methods
2.1.Study area
The study area covers the northwestern corner of Africa in Morocco(Fig.1A),which is crossed by the Rif and Atlas mountains(Tubkal mountain,4164 m a.s.l.,is the highest altitude).Since these mountains intercept the wet air masses arriving from the Atlantic Ocean,they strongly affect local rainfall,showing a more humid climate on the north-western than on the south-eastern slopes.Mountains also produce a sharp altitudinal gradient of environmental conditions,with lower temperatures and more frequent snowfalls athigher altitudes(Boudharetal.,2011;Marchane etal.,2015).As a consequence of these climatic factors,the area is occupied by humid forestsin the north(Quercus suber,Q.pyrenaica),Fir(Abies marocana)and Cedar (Cedrus atlantica) forests in some western slopes,drought-tolerant woodlands (Q.ilex,Juniperus thurifera,Tetraclinis articulata)in lowland areas,and Argan(Argania spinosa)and Acacia(Acaciaspp.)woodlands in the southernmost sectors(Fig.1B;Quézel,1983).
2.2.Altitudinal distribution of weather conditions
We studied the winter distribution of temperature and snow cover of the sites where chaffinch abundance was estimated(see below)to assess the harshness of winter conditions.For this purpose,we used the minimum mean temperatures(°C)of the coldest moth in CHELSA V 1.0 at a horizontal resolution of 30 arcsec (~1 km) for the period 1979-2013,which provides improved climatic estimates in landscapes with complex topography(http://chelsa-climate.org/;Karger et al.,2017).In addition,we recorded the monthly snow cover during the period November-March (2015-2017) provided by the Nasa Earth Observatory at a horizontal resolution of 0.1°(https://earthobservatory.nasa.gov/globa l-maps/).This databank provides the monthly snowfall cover (si,in %)by using an index that ranges from 0 to 100%.In this way we calculated the altitudinal distribution of the mean monthly snow cover ((∑si)/15 months,i=1-15) during the study period.The altitude pattern of this mean monthly cover of snow was used to divide the altitude gradient in sectors above and below a threshold of potential strong snow cover effects on birds.All these cartographic data were managed with QGIS 3.4.15(QGIS Association,2021).
2.3.Abundance
We assessed African Chaffinch abundance in 99 tree covered farmland and woodland sites distributed along an altitude gradient in spring(second half of April and May of 2015,2016 and 2017)and in other 123 different sites in winter (January 2013;see Tellería et al.,2014,2021).This sampling was aimed to work with independent samples covering the altitudinal distribution of wooded areas inhabited by African Chaffinches(Cramp and Perrins,1994).Within each study site,we recorded the individuals detected along a 500-m-long transect (Bibby et al.,2000).To account for the effects of habitat structure on species abundance,we assessed wood cover(vegetation >2 m height)in two 25-m-radius circles located at 150 and 350 m from the starting point of each transect.
2.4.Morphology
We mist-netted African Chaffinches from the end of April to the end of June (2007,2011,2018 and 2019).This period encompasses the breeding time of the species minimizing the probability of capturing vagrant individuals.The sampling sites were located around the villages of Larache(35.1745°N,6.1475°W;mean altitude of sampling sites 10 m a.s.l.),Chauen(35.1704°N,5.2690°W;450 m a.s.l.),Taza(34.1487°N,4.1265°W;1400 m a.s.l.),Azrou(33.4388°N,5.2222°W;1800 m a.s.l.)and Ouirgane (31.1757°N,8.080°W;970 m a.s.l.).Mist-netted birds were aged according to the EURING code classification (age 3: hatched during the year of capture;age 4: hatched before the year of captureexact year unknown,age 5: hatched during the previous calendar year;age 6:hatched at least two calendar years before the year of capture)and sexed (males and females) following Svensson (1992) and Jenni and Winkler (1994).In subsequent morphological analyses,we only considered fully-grown individuals (n=97) of known plumage type,which restricted our dataset to birds coded as Euring 5(juvenile plumage)and 6(adult plumage).From these individuals,we obtained the length of several morphological variables using the appropriate rulers or a digital caliper: right wing (maximum wing chord method),primary 8th,bill(from skull base to bill tip),right tarsus (minimum tarsus) and tail (see Svensson,1992 for more details).We also recorded body mass from each individual using a digital balance (resolution 0.1 g).These six morphological variables were used to carry out a principal component analyses(PCA) to detect some orthogonal variables to segregate individuals according to body size and flight related traits.
During 2018-2019,we studied the wing shape of 81 chaffinches in Chauen,Taza,Azrou and Ouirgane by measuring the primary distances of the 9 longest primaries (excluding the vestigial outermost primary P10).Primary distance was defined as the distance from the tip of each primary to the tip of the longest primary with the wing folded,assigning a value of zero to the primary(or primaries)constituting the wingtip.For analyzing wing shape configuration,we opted for transforming primary distances into distances from the carpal joint (cP1-cP9) by subtracting,for each primary,its primary distance from the wing length.These transformed distances (cP1-cP9) were then standardized according to the method suggested by Senar et al.(1994),which provides more reliable measurements(cP1*-cP9*)that correct for the among-individual variation in wing size.These standardized values were included in a PCA performed with the prcomp function in R,which gave rise to two principal components(wing concavity and wingtip pointedness)that reflected variation in wing shape among chaffinches.Captures and measurements were carried out by IHT,AO and JIA,experts of the Spanish Ringing Scheme who had their measurement protocols standardized and highly repeatable,and worked under the permission of the Moroccan authorities.
2.5.Analyses
We performed a multivariate approach to test for the effects of altitude on the seasonal distribution of Chaffinch abundance.We specifically tested whether the relationship between bird abundance and altitude varied between seasons(breeding vs.winter)according to the prediction that bird numbers should decrease at high altitude during winter,and increase in lowland areas.In order to test this hypothesis,we included the “season × altitude” interaction term in our model.In the case of altitude and according to our proposal in the Introduction section,we carried out two alternative analyses: one of them using the altitude gradient as a covariate (“altitude gradient”),and another one using altitude as a two-level factor,in which the study sites were classified as lowland or highland localities from a threshold of frequent snow covers(“altitude threshold”).After testing with ade-4 in R (Thioulouse et al.,2018) that abundance distribution of the African Chaffinch was not spatially autocorrelated(Mantel's test,r=0.035,p=0.160,n=222),we dealt with the large number of zeros in transects(25%of sampling points,Fig.2) by using the hurdle count model approach provided by pscl-R(Zeileis et al.,2008).This approach is composed of a hurdle component that models the zero counts and a truncated count model for positive counts.The hurdle model is a binomial logit regression and shows the features related to the occupancy(presence/absence)of the species.The count model is a truncated Poisson or negative binomial(with log link)model that explores the features affecting the abundance distribution.
We used linear models to explore the patterns of variation in bird morphological characteristics after testing with ade-4 in R the existence of spatial autocorrelation in all the study variables.These tests showed no evidence of spatial autocorrelation (all Mantel tests were nonsignificant).In this case,variation of morphological components resulting from principal component analysis(PCA;see Results)were analyzed in relation to altitude (independently for altitude gradient and threshold),sex (male vs.female) and age (Euring 5 and 6),that usually influence the observed patterns(e.g.de la Hera et al.,2014).We initially considered the two-way interactions between these three variables,but their effects were not included in the final models since they were non-significant.These morphological analyses were carried out with Rcmdr (Fox and Bouchet-Valat,2020) and all of them fulfilled the assumptions of general linear modelling.
3.Results
3.1.Environmental conditions and altitude
Tree cover increased progressively with altitude (Spearman correlation,rs:0.30,p<0.05,n=222),particularly over 1000 m a.s.l.However,mean temperatures of the coldest month decreased monotonically along the altitude gradient (Spearman correlation,rs: -0.91,p<0.001,n=222),so that altitude can be considered a good surrogate of this environmental trait in the study area.Snow cover was positively correlated with altitude(rs:0.69,p<0.001,n=222),but showed a sharp increase over the 1500 m a.s.l.(Fig.2).Thus,this altitude was selected as a putative threshold over which many African Chaffinches could be forced to leave the highlands during winter.
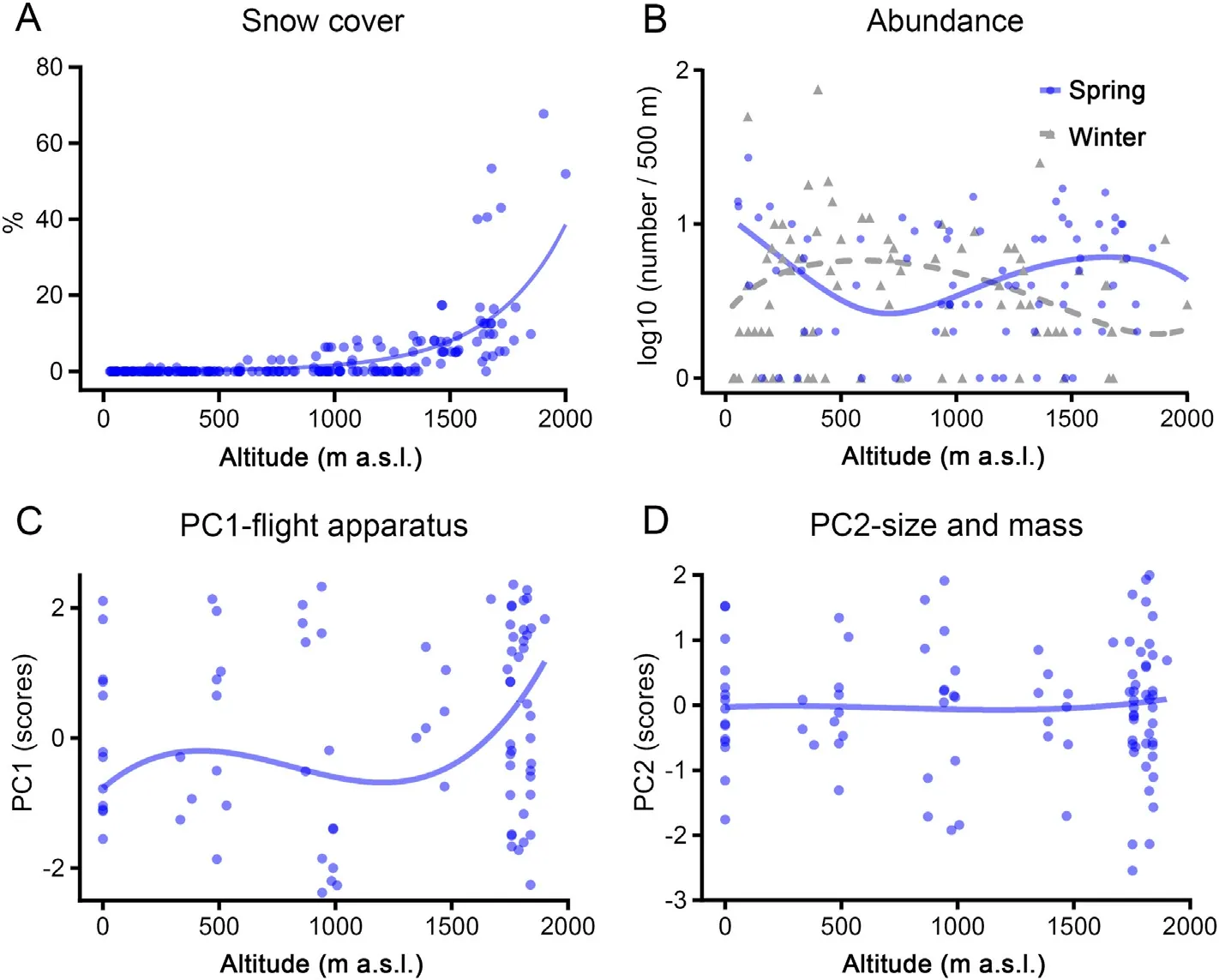
Fig.2.Altitudinal distribution of mean monthly snow cover(A)and spring and winter abundance distribution of the African Chaffinch(B)within the study area.The size of the external flight apparatus(C)and body size(D)of this species along the altitude gradient are also showed.In all the figures,distance weighted least squares lines have been adjusted to better visualize how the patterns change with elevation.
3.2.Bird abundance
The raw data of African Chaffinch abundance reflected a decrease in bird numbers during the winter in highlands over 1500 m(Table 1).After testing for the equi-dispersion assumption,the multivariate hurdle model suggested that both the occupancy and the abundance were strongly mediated by the distribution of tree cover and that the abundance was negatively related to altitude (Fig.2;Table 2).We did not find an interaction between season and altitude when this effect was included in the model as a continuous variable(i.e.altitude gradient;Fig.2;Table 2).However,this effect was stronger on bird abundance when the altitude was considered as a two-level factor(altitude threshold;Table 2).Further analyses in which the seasonal changes in abundance were compared reflected a significant decrease of bird numbers from summer to winterabove 1500 m(season:-0.61±0.72,p=0.010;tree cover:0.18±0.10,p=0.084,n=43),but the complementary increase in abundance from summer to winter under this altitudinal threshold was non-significant(season: 0.23 ± 0.19,p=0.240;tree cover: 0.49 ± 0.12,p<0.001,n=177).This supports a numerical rearrangement between seasons,whereby African Chaffinches experience a winter decrease in abundance over 1500 m(Tables 1 and 2).
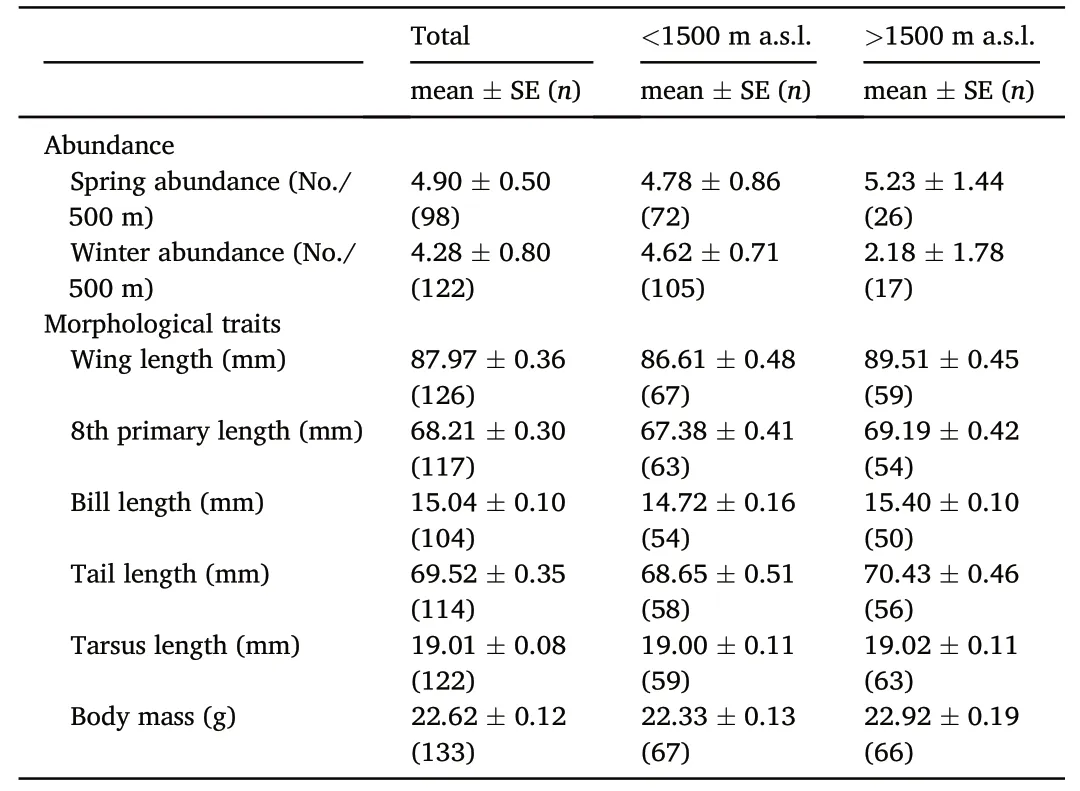
Table 1 Mean abundances and morphological traits of the African Chaffinch in the study area.These scores are also arranged according to the distribution of individuals above or below the altitude threshold of 1500 m a.s.l.
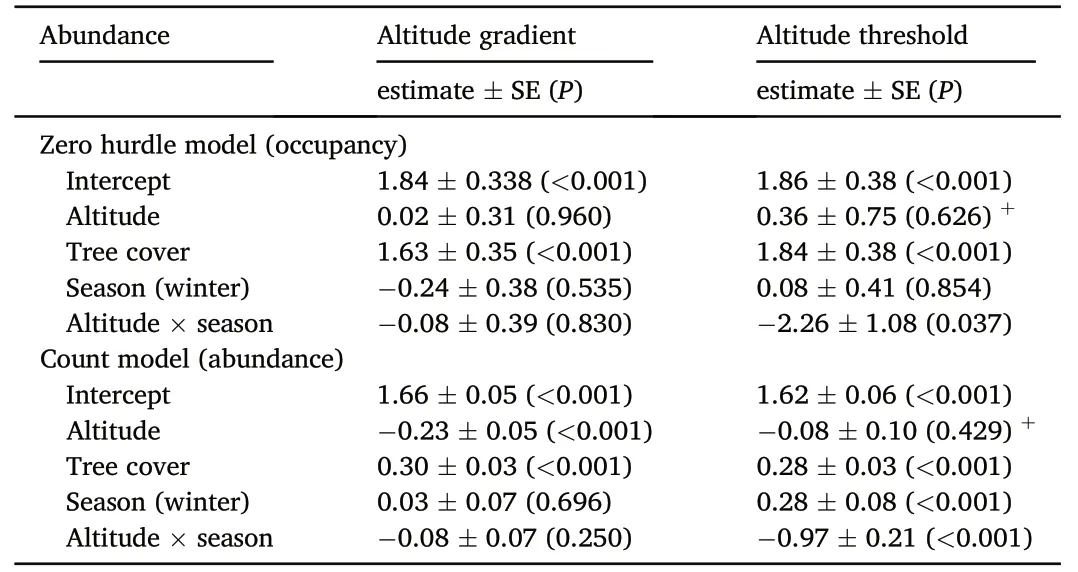
Table 2 Results of a hurdle count model approach to explain African Chaffinch abundance distribution in north-western Africa.Zero hurdle models show the features related to the occupancy of species and count models the features affecting abundance.The results show the coefficients (b ± SE),the results (P) of the associated t-tests and the best spatial correlation structure.+:under 1500.
3.3.Morphology
The raw data of morphological traits reflected longer wings,8th primaries and tails in highlands over 1500 m(Table 1).The results of a first Principal Components Analysis (PCA) that included the five lineal morphological measurements described above (Table 1) produced two relevant principal components.The first component reflected the degree of development of the flight apparatus(PC1,eigenvalue=3.10,variance=0.62,factor loadings:wing=0.55,p8=0.54,bill length=0.25,tarsus=0.25,tail=0.52),whereas the second component was related to tarsus length vs.bill length (PC2 eigenvalue=0.86,variance=0.17,factor loadings:wing:0.03,p8:0.03,bill length:-0.83,tarsus:0.56,tail:0.06).To test if those individuals with longer tarsus(a usual surrogate of body size;Senar and Pascual,1997)were also the heaviest ones and to explore their distribution along the altitudinal gradient,we included body mass in a new PCA.In this case,PC1 was positively correlated with the six variables included in the analysis but more strongly with those associated with flight-related morphology (eigenvalue=3.31;variance=0.55;factor loadings:wing: 0.52,p8:0.51,bill length:0.25,tarsus:0.26,tail:0.50,body mass: 0.29),so that we interpreted PC1 as an index of the development of the flight apparatus.Body mass and tarsus length were the variables more strongly associated with PC2 (eigenvalue=1.02;variance=0.17;factor loadings: wing: -0.23,p8: -0.30,bill length:0.29,tarsus: 0.57,tail: -0.26,body mass: 0.62).This pattern suggests that chaffinches with higher scores in PC2 were the largest and heaviest ones within the study population,reflecting chaffinch body size.As individual component scores resulting from the two PCA approaches were highly correlated,we finally opted for using the PCA that included body mass to explore the changes in the flight apparatus(hereafter PC1-flight apparatus) and body size and mass (hereafter PC2-size and mass) along the altitude gradients.
The PCA on the standardized primary feather distances provided two principal components.PC1 was interpreted as an index of wing concavity(eigenvalue=3.61;explained variance=0.40;factor loadings: P9=0.21,P8=0.27,P7=0.24,P6=-0.12,P5=-0.29,P4=-0.41,P3=-0.47,P2=-0.40,P1=-0.43),where individuals with higher PC1 scores showed proportionally shorter inner primaries,that is more concave wings.On the other hand,PC2 reflected variation in wingtip pointedness (eigenvalue=1.63;explained variance=0.18;factor loadings:P9=0.54,P8=0.48,P7=0.42,P6=-0.33,P5=0.14,P4=0.18,P3=0.22,P2=0.22,P1=0.17),whereby higher PC2 scores were associated with longer outer primaries for a given degree of wing concavity,or more pointed wingtips.
The models were carried out by considering all the two-way potential interactions between altitude,sex and age,but all these interactions were non-significant.Thus,we rerun the analyses without interactions to improve their ability to detect the effect of the involved variables.After accounting for the effects of sex (males larger than females) and age(individuals of age 6 were larger than those of age 5)in the models,PC1-flight apparatus clearly increased with altitude supporting a morphological differentiation of African Chaffinches over such gradient(Figs.2 and 3;Table 3).However,PC2-size and mass increased in males but did not vary along the altitude gradient and was not affected by age(Table 3).In the case of wing configuration,chaffinches in highlands reported more concave wings (PC1-wing concavity) than their lowland counterparts,but did not differ in wingtip pointedness (PC2-wingtip pointedness;Fig.3;Table 3).
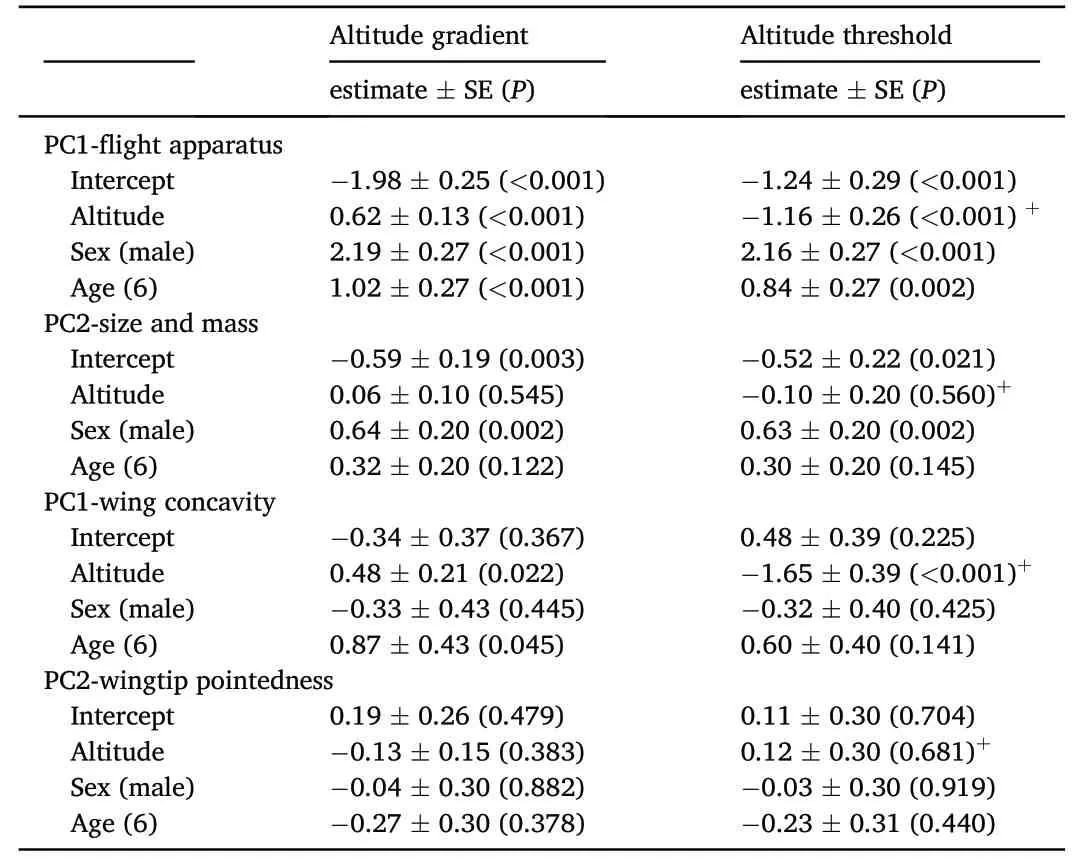
Table 3 Results of the General Lineal Models in which the components related to the development of the flight apparatus (PC1 of a PCA with the six morphological variables detailed in the Methods) and body size (PC2 of the same PCA that included with six morphological variables PCA),wing concavity and wingtip pointedness have been analyzed in relation to altitude (altitude gradient or altitude threshold)after controlling for the effect of sex and age.The results show estimates(±SE),and significance values(P)according to the associated t-tests.+:under 1500.
4.Discussion
Rear edge populations of birds occur in refuge areas in the southwestern Palaearctic (Merker and Chandler,2020) from where other populations of the same species progressively colonized more northern areas after the Quaternary ice retreat (Hewitt,1999;Husemann et al.,2014).In this historical context,migratory behaviour would have been a way to track the increasing availability of seasonal resources in the newly ice-free areas arising at higher latitudes(Bruderer and Salewki,2008).It can be suggested that the evolution of latitudinal migrations in the Eurasian Chaffinch populations were also accompanied by altitudinal displacements at a more local scale,and that both seasonal movements affected similarly several morphological and life history traits (Boyle et al.,2016).Results in this paper support this view and provide information on the way African Chaffinches could track the spatial-temporal environmental patterning within the south-western edge of the Palaearctic.
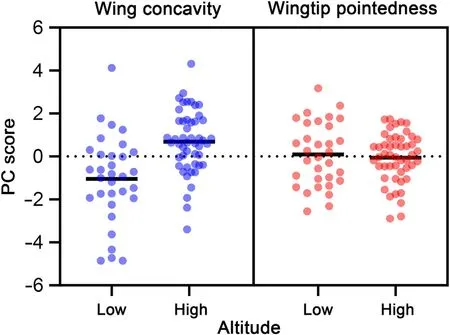
Fig.3.Distribution of wing concavity and wingtip pointedness of African Chaffinches according to their distribution in lowlands and highlands (under and above 1500 m a.s.l.) of the study area.Bars show mean scores.
4.1.Seasonal changes in abundance
Most rear-edge populations of forest birds inhabiting north-western Africa show a similar distribution,with their most suitable breeding habitats located in humid northern and western slopes of mountain ranges (Tellería et al.,2021).Results in this paper show that the distribution of the African Chaffinch tracks tree covered habitats within a broad range of altitudes where they occur all around the year (Fig.2B;Table 2).Despite this ubiquitous distribution along the altitude range,species abundance showed a significant decrease in winter over the 1500 m a.s.l.,supporting the presumed link between the intensity of snowfalls and the winter abandonment of highlands by many African Chaffinches(Tables 1 and 2;Fig.2).This reaction to seasonal changes supports the view that over a given altitude threshold,many chaffinches move to winter in lowlands,as reported in other areas of the south-western Palaearctic (Carrascal and Palomino,2012;Tellería et al.,2020).The true extent of these movements is far from being known,although most of them revolve within the breeding range of the species and may sometimes expand to nearby areas,such as the northern edge of the Sahara at the Atlas piedmont(Thévenot et al.,2003).
The presence of African Chaffinches at the highest sectors during winter and the poor effect of these numerical rearrangements on the abundance in lowlands could be explained by some idiosyncratic traits of both the species and the study area.Many chaffinches (particularly males) are reluctant to leave their breeding grounds in winter (Cramp and Perrins,1994).In this context,as snowfalls are uncommon in dry and sunny African high slopes in the shadow of oceanic rains (Fig.1A;Marchane et al.,2015),many highland chaffinches could overwinter nearby the breeding grounds by moving along slopes in the search of suitable habitat patches as it has been reported in other mountain ranges(Carrascal et al.,2012).The number of individuals involved in these altitudinal movements will probably be small if we have into account the limited area covered by woodlands (their breeding habitat) over the 1500 m threshold(Fig.1C).This trait could dilute the effect of displaced individuals on the detected abundance patterns in lowland areas(Table 1).It also remains obscured the actual effect of the Eurasian Chaffinches arrived to winter from Europe(Thévenot et al.,2003)on the observed seasonal patterns.As these birds are not easy to detect in field counts (e.g.immature and females are very similar to the African subspecies),they could distort the observed numerical arrangements of the African population.For instance,it could be speculated that an overall winter loss of individuals within the African range(e.g.mortality)could be compensated by the arrival of foreign individuals in lowlands areas,which are the best wintering grounds for birds in Morocco(Tellería et al.,2014).Thus,to cope with these potential difficulties,we used the spatial variation in bird morphology to interpret the extent to which African Chaffinch movements might occur within the geographical setting of Morocco(Tellería et al.,2013).
4.2.Morphological variation
Morphological change is a widespread response to environmental change and can occur through phenotypic plasticity and/or natural selection(Gienapp et al.,2008).In this way,the presence of morphological clines can be used to detect the response of populations to environmental gradients at different spatial scales(Richardson et al.,2014).
The altitudinal variation in the external flight apparatus of the African Chaffinch supports a different morphological configuration over and below 1500 m a.s.l.(Tables 1 and 3;Figs.2 and 3).More explicitly,our results show that birds above that threshold have more developed flight morphology (longer wing and tails) than their lowland counterparts(Tables 1 and 2;Fig.2).These results show an atypical pattern if we consider that a combination of longer wings and shorter tails have usually been related to flight efficiency in migratory flapping-flight birds,where long wings are designed to increase flight power and short tails to reduce drag (Leisler and Winkler,2003;Hedenström,2008).However,this pattern is not universal or can vary among species(de la Hera et al.,2012;Fudickar and Partecke,2012;Huber et al.,2017).For instance,the pattern observed in the African Chaffinch could be related to its bounding-flight,where longer wings would improve flapping in the powered phase and longer tails would increase lift and stability in the passive wing-folded phase (Sachs and Lenz,2011).Thus,longer wings and tails could enhance flight efficiency in highland African Chaffinches,an interpretation also supported by wing shape(Table 3;Fig.3).In this case,more concave wings were detected in highland chaffinches,a trait usually related to flight efficiency of birds involved in migratory movements(Rayner,1988;Lockwood et al.,1998;Leisler and Winkler,2003).
Although we believe that seasonal migration is the most likely interpretation for the morphological differences observed between high and low altitude chaffinch populations,we acknowledge that these morphological changes could also be alternatively,or complementary,explained by other factors.For instance,it has been shown that changes in forest could drive intraspecific wing morphology of forest birds(Desrochers,2010).In this context,it can be suggested that a denser tree cover could affect flight morphology as birds show more rounded wings to improve manoeuvrability (Savile,1957;Noreau and Desrochers,2018).But tree cover,a trait related to the physiognomic complexity of woodlands in the study area (Tellería et al.,2021),increased with altitude making difficult to link this pattern with the increased wing concavity of the highland chaffinches.Decreasing air density could also affect these patterns,as it has been suggested before for flying birds(e.g.altering lift and drag balance,increasing wing beats;Pennycuick,2001;Schmaljohann and Liechti,2009).But the putative effect of air density on the intraspecific morphological variation of birds has not been proved to date.In addition,we presume that this putative change would affect wing morphology over the whole altitude gradient and not over a given altitude threshold as reported in this study.In any case,our study encourages further research on the potential influence of these and other environmental factors in shaping avian wing morphology.
Thus,it can be summarized that(i)the external flight apparatus of the African Chaffinch varies along the altitudinal gradient,(ii) that the changes are detected between individuals breeding over and below an altitudinal threshold related to winter snow cover and (iii) that these changes could be related to an enhanced flight efficiency of the involved populations.In this context,it is interesting to highlight the lack of change in size and body mass of the African Chaffinch along the altitude gradient (Table 1;Fig.2D).This could be an idiosyncratic trait of this subspecies,reputed as one of the largest ones (Perktas¸ et al.,2017),which is not compelled to increase body mass to cope with the colder conditions of highlands(Sander and Chamberlain,2020).
5.Conclusions
Results in this study support several main conclusions related to the way a rear-edge bird population face the environmental changes in the south-western edge of the Palaearctic.Far from being the sedentary counterpart of a partial-migratory species,the African Chaffinch evolved independently within the seasonal environmental setting of its range.This process,which agrees with the morphological and genetic uniqueness of this population (Perktas¸ et al.,2017),could be applied to many other birds and taxa of northern Africa,given the high degree of endemicity in this region and the presumed adaptations to local conditions(Hampe and Jump,2011;Husemann et al.,2014;Isenmann and Thévenot,2018).This leads to several additional conclusions.First,these endemic populations cannot be viewed any more as the maladapted counterparts of the core populations(Angert et al.,2020 for discussion).Second,the morphological diversity associated with altitude within the African Chaffinch range underlines its putative adaptive ability to track the existing spatial-temporal environmental dynamics(Richardson et al.,2014).This is an interesting trait in a context of climate change where the less drought tolerant woodlands preferred by the species(Tellería et al.,2021) will shift upwards along the altitude gradient (Ruiz-Labourdette et al.,2012).It has been suggested that,in this context,the adaptive capacity of populations will be linked to their ability to track the spatial-temporal patterning of habitat suitability (Hampe and Jump,2011;Hannah et al.,2014),a process strongly linked to the idiosyncratic circumstance of the involved species (Angert et al.,2011;MacLean and Beissinger,2017).Finally,the meso and microclimatic conditions created by mountains in the southern edge of the Palaearctic highlight not only their role as refuge areas for northern organisms during glacial periods(Husemann et al.,2014),but also suggest a role as drivers of population diversification in highly mobile organisms (e.g.birds).This makes the protection of habitats within these altitude gradients (e.g.tree covered areas for the African Chaffinch)a main conservation strategy of rear-edge populations of birds(Tingley et al.,2014).
Author contributions
All authors designed the study;AO,IH-T,JIG,JLT participated in the field work;JLT and IdH carried out the statistical analyses and drafted earlier versions;all authors improved successive versions.All authors read and approved the final manuscript.
Declaration of competing interest
The authors declare that they have no competing interests.
Acknowledgements
We thank Cristina de Castro,Roberto Carbonell,Nadia Chekrouni,Marina Guerrero,Mohammed Hakkou,Mohammed Hilmi,María León,Maria Jesús de Lope,Carlos Martín,Álvaro Ramírez and Juan Ramirez for support in field work;and Abdeljebbar Qninba,Hamid Rguibi and the officers of the Haut Commissariat aux Eaux et Forêts et à la Lutte contre la Désertification for permissions to capture and measure birds in Morocco.Two anonymous reviewers considerably improved an early version of this manuscript.This paper is a contribution to the project CGL2017-85637-P (Life at the border: population differentiation of forest birds south of the Palearctic) granted by the Spanish Ministry of Science,Innovation and Universities.
杂志排行

Avian Research的其它文章
- Functional and phylogenetic structures of pheasants in China
- Thermoregulatory function and sexual dimorphism of the throat sack in Helmeted Guineafowl (Numida meleagris) across Africa
- Distribution pattern and driving factors of genetic diversity of passerine birds in the Mountains of Southwest China
- Multiple lines of evidence confirm that the critically endangered Blue-crowned Laughingthrush(Garrulax courtoisi)is an independent species
- Corrigendum to “Multiple lines of evidence confirm that the critically endangered Blue-crowned Laughingthrush (Garrulax courtoisi) is an independent species” [Avian Res.13 (2022) 100022]
- Shifts in phenology of autumn migration and wing length among reedbed passerines along the East Asian-Australasian Flyway