Long-term monitoring data reveal effects of age,population density,and environmental aspects on hatching success of Common Cranes (Grus grus)
2022-10-03IselBrwischWolfgngMewesAngelSchmitzOrn
Isel Brwisch ,Wolfgng Mewes ,Angel Schmitz Ornés,*
a AG Vogelwarte,Zoological Institute and Museum,University of Greifswald,Soldmannstr.23,Greifswald,Germany
b Grüner Weg 3,19395,Plau Am See,OT Karow,Germany
Keywords:Adaptation Age Cranes Grus grus Habitat Nest survival Population density Reproduction
ABSTRACT Facing climate and land use change,a species’ability to successfully adapt to changing environments is crucial for its survival.Extensive drainage and intensification of agriculture and forestry set wetlands and associated species at risk of population declines.The population of Common Cranes (Grus grus) has experienced considerable fluctuations over the last century.Despite increasing population numbers,hatching success seemed to have decreased over the last years.The aim of this study was to identify factors influencing hatching success and nest survival of Common Cranes based on analyses of long-term individual-based monitoring data from northeastern Germany and evaluate the species ability to adapt to changing environments.Hatching success decreased over the course of the study period from 0.75 to 0.55.Surprisingly,nest survival and hatching success did not vary across different nesting habitats,whereas factors such as female age,timing of nest initiation and breeding pair density were found to have significant effects on hatching success.Older females showed higher hatching success,even though the proportion of unhatched eggs was highest in females aged 20 years or older.Early nest initiation had a positive effect on hatching success.Water levels are more favorable early in the nesting season,whereas increasing evaporation with time causes water levels to decrease,granting easier access for predators.Independently of female age,hatching success decreased with increasing numbers of breeding pairs within a 2-km radius around a nesting site.High population densities intensify competition for resources and promote intraspecific interactions,affecting reproductive outcome negatively.This study gives first insights into mechanisms behind population regulation in Common Cranes,highlighting the importance of population dynamics and individual features.We suggest to further investigate density dependent effects including landscape and habitat features as well as reproductive success in terms of chick survival,since successfully raising juveniles is crucial for a species survival.
1.Introduction
Effective monitoring is a valuable tool for determining population developments and changes over time,e.g.,in association with changing landscapes due to human impact or climate change (Heldbjerg et al.,2018).In addition to climate change,land use and habitat fragmentation are considered to mainly be responsible for the loss of biodiversity across all animal taxa.Intensification of agriculture and forestry as well as rapidly developing urbanization,including infrastructure,are major threats to wildlife on a global scale (Chamberlain et al.,2001;Donal et al.,2001;Laurance,2001;Ehrlich,2004;Vale et al.,2008).Possibilities and opportunities for plastic responses including behavioural adaptation,genetic adaptation or range shifts as reactions of species to the loss of suitable habitats are often rather limited causing populations to decline or even disappear (Parmesan,2006;Thomas,2010).To effectively counteract the biodiversity crisis,it is crucial to increase conservation efforts especially protecting natural areas and renaturation.Wetlands are particularly prone to be negatively affected by land use change,mainly due to drainage or over-exploitation(Hu et al.,2017)and global warming,e.g.rising temperatures along with increasing evaporation (Briffa et al.,2009;Teuling et al.,2019).Wetland degradation is assumed to be responsible for the decline of more than half of the vertebrate populations linked to wetlands (Dudgeon et al.,2006).Additionally,increasing human interference has been directly linked to decreases in population densities and reproductive success of many wetland bird species (Kentie et al.,2013;Fraixedas et al.,2020;Moreno-Opo,2020;Sica et al.,2020).
Survival rates and reproductive success have a determining influence on a species’ ability to successfully adapt(Nuijten et al.,2020) and this adaptation to drastically changing environments has been proven only for a few species,so far(Trautmann,2018).Moreover,we need to gain a better understanding about the complex mechanisms behind successful adaptation.
Extensive drainage and intensification of agriculture during the 19thand 20thcentury caused many wetland bird species to disappear from certain regions within Europe,one example being the Common Crane(Grus grus,Linnaeus,1758;Meine and Archibald,1996).Due to land use change and human agricultural activities the population of Common Cranes almost disappeared during the mid-20thcentury in several European countries.However,conservation efforts enabled the Common Crane to repopulate most of its former distribution area and recover to approximately 220,000-250,000 individuals during the mid-1990's(Meine and Archibald,1996;Lehrmann et al.,2016).The Common Crane is a long-living migratory bird species of the family Gruidae distributed across northern Europe and Asia.The species was known to primarily populate forested areas nesting within swamp forests and big mire complexes(Prange et al.,2016).On average,cranes start reproduction at the age of four or five years,usually lay two eggs,and can lay replacement clutches when losing the first one (Prange et al.,2016).Cranes'nesting season in northern Germany usually starts in March and lasts throughout June or even July,depending on individual traits and weather conditions(Mewes,2019a).
The species is currently listed as species of ‘least concern’ in the International Union for Conservation of Nature (IUCN) Red List of Threatened Species (Bird Life International,2016) and showed continuous positive population trends over the last decades,despite intensive agricultural and silvicultural developments (Leito et al.,2003;Boldt,2015).Nowadays,the global population is estimated to exceed 700,000 individuals (International Crane Foundation,2021).However,recent studies suggest a decreased reproductive success within several areas of high nesting population densities within Germany,like in the federal states Mecklenburg-Western Pomerania and Brandenburg (Mewes,2014a,b),whereby causes of differences in reproductive success between regions remain unknown.
Some researchers suggest that the species has shown changes in nesting habitat use over the last decades due to increasing population numbers and limited availability of preferred nesting habitats.Whereas only 8%of the crane population in Mecklenburg-Western Pomerania and Brandenburg nested within agricultural landscapes in 1996,this proportion increased to more than 30%in 2010(Mewes,2010).Therefore,there might be a correlation between the increasing proportion of crane pairs breeding within intensively used agricultural landscapes,referred to as suboptimal breeding grounds,and a decreased reproductive success(Mewes,personal communication).
High population densities intensify competition for territories,good nesting sites and food sources(Fernandez et al.,1998),and might affect reproductive success negatively.Density-dependent decreases of reproductive success have been documented for other bird species,like Mute Swans (Cygnus olor),Bewick's Swans (C.columbianus bewickii) and Barnacle Geese (Branta leucopsis),already (Larsson and Forslund,1994;Nummi and Saari,2003;Solovyeva et al.,2019).
The overall loss of appropriate wetlands for nesting,due to drainage and intensive agricultural land use additionally intensifies competition for resources in cranes (Nowald et al.,2010).An impact of habitat selection on reproductive success has been proven for other bird species(Kentie et al.,2013;Léandri-Breton and Bêty,2020),but at the same time researchers also found numerous indications that habitat selection plays a rather minor role concerning hatching success in several species (Ballantyne and Nol,2011;Sakseide,2020).An individual's experience seems to have a stronger impact on reproductive success instead (Noreikien˙e et al.,2021).Effects of age and reproductive history have not been studied for cranes so far but are expected to be positively correlated with reproductive outcome as it is also the case in other long-living bird species,e.g.Eiders(Somateria mollissima),Houbara Bustards(Chlamydotis macqueenii) or Mediterranean Storm Petrels (Hydrobates pelagicus;Van Heezik et al.,2002;Hernández et al.,2017;Noreikien˙e et al.,2021).However,Leito et al.(2003)found age to be linked to habitat selection,as well,which demonstrates the complexity and species specificity behind mechanisms determining reproductive success.
Since little is known about crucial factors having an impact on the reproductive performance of Common Cranes we focused our study on the effects of different habitats,weather conditions,timing,female age and population density on hatching success and nest survival.We expected hatching success to increase with female age and assumed hatching success to be higher at traditional nesting sites,i.e.forest wetlands and mire complexes,compared to rather newly populated habitat types located within intensively used agricultural landscapes.Considering the breeding biology of Common Cranes,we expected hatching success to be positively correlated with the amount of precipitation prior to the breeding season,since higher water levels are associated with better protection against predators.The objectives of our research are to gain a greater knowledge of the reproduction dynamics of the species and its breeding biology in general.Understanding the mechanisms behind reproductive performances and adaptation to changing environments is crucial for the protection of a species (Zhu et al.,2012).Information about the causes of reduced reproductive success will help to establish suitable management and conservation measures concerning long-term species conservation.
2.Methods
2.1.Study area
The study was conducted in the German state of Mecklenburg-Western Pomerania(34,100 ha).The study site,hereafter referred to as Goldberg,extends from the Plauer See in the east to the city of Parchim in the west and is part of the northern Lowland.The area is characterized by scattered woodland areas (13.8%),large portions of agricultural landscape (66.3%),grassland(11.0%) and a variety of water bodies such as rivers,lakes,swamps or different kinds of pothole wetlands (Mewes,2014a,b).Annual precipitation in the region ranges from 585 to 600 mm and annual average temperature is 8.4°C (German Meteorological Service,2018).
Overall breeding pair density in Mecklenburg-Western Pomerania increased from 17 breeding pairs per 100 km2to 21 breeding pairs in 2020 (Lehrmann,2020).However,population density varies considerably on smaller spatial scale.Goldberg represents a region of high population density of reproductive pairs of Common Cranes.Mewes(2019a,b)estimates 42 breeding pairs per 100 km2making it an area with one of the highest densities of nesting Common Cranes across Germany (Lehrmann,2020).
2.2.Data collection and preparation
2.2.1.Hatching success
Monitoring of crane nesting sites in this area started in 1988.In 2002 data collection became standardized and was continued by WM throughout the years until today.For this reason,we restricted our analyses to the data collected between 2002 and 2020.
Field work usually started in March,although there was variation depending on the weather and cranes’ return from their wintering grounds.We systematically searched the study area for crane nests.For each nest we collected coordinates,as well as information on the habitat location within the landscape.Habitat locations of monitored nesting sites included woodland (WL;entirely surrounded by at least one continuous treeline),open landscape (OH;not surrounded by forest structures,single trees or bushy structures possible) and intermediate habitats (WOH),which are located on the edges of forests and open landscapes.Additionally,the size (length × width) and weight of each egg were measured.We used these parameters to calculate approximate age of the clutches (Mewes,2019b).Knowing the approximate laying date,hereafter referred to as day of nest initiation,the day of hatching could be roughly estimated due to the fact that Common Cranes incubate their eggs on average for 30 days(Prange et al.,2016).We checked the nests again around the assumed day of hatching of the first clutch(FC).If at least one chick had hatched,we considered the nest successful.In cases of nest failure,we checked nesting sites for replacement clutches(RC)at least once again later that season.The occurrence of foul eggs,e.g.unfertilized eggs,was noted as well.We did not include nests with unknown or uncertain fate in data analyses.
Apart from parameters collected during field work,additional ones were added to the data set.Weather parameters were provided by the German Meteorological Service(DWD,Deutscher Wetterdienst):amount of precipitation prior to the nesting season(November-April),cold sum of the preceding winter-a measurement to account for the harshness of a winter,average temperature during the nesting season (March-May),and total sunshine duration(March-May).We used the software ArcGIS(Version 3.4.15)to determine the number of crane pairs nesting within a 2-km radius around each active nest.Numbers of breeding pairs were calculated for each year separately,based on nest coordinates.We included these values in statistical analyses as a measure of population density.Before 2005 there was no collection of coordinates of crane nests,therefore nesting density could not be estimated for the period 2002 to 2004.Due to insufficient data on 2ndreplacement clutches we did not differentiate between 1stand 2ndreplacement clutches during statistical analyses.
Using an already established method to identify female cranes through their eggs based on morphological features(Mewes and Rauch,2010;Schmitz Ornés et al.,2014;Höltje et al.,2016;Mewes,2020),we were able to assign clutches to specific females,enabling the recording of life and reproductive history of single female cranes for up to 30 years.Since this identification method can only be applied to breeding females,their ages cannot be known exactly.However,due to high nesting site fidelity in cranes (Mewes,2017),constant comprehensive breeding site monitoring during the study years,and the assumption that Common Cranes start to reproduce at the age of 4 years old,on average (von Blotzheim et al.,1994;Prange et al.,2016),females appearing for the first time were assumed to be 4 years old,conducting their first breeding attempt.
2.2.2.Nest survival
Hatching success is a good measure for reproductive success in general.Unfortunately,more detailed information on changes in the probability of the clutch surviving the incubation period over the course of the breeding season are missing.Therefore,we used a complementary study set up to investigate daily survival rates (DSR) of nests.From 2016 to 2021,a total of 157 nesting sites of Common Cranes have been equipped with camera traps to monitor nesting periods in greater detail and gather information on cranes’ behaviour and external factors affecting reproductive outcome.We made use of collected photo material to generate encounter histories for analysing nests DSR(Rotella,2018).
Following Rotella (2018) encounter histories included: (1) day the nest was found;(2)last day the nest was active;(3)last day the nest was checked;and(4)fate of the nest(i.e.,hatched or failed).For this purpose and a standardization of nesting seasons among years,we converted ordinal dates into numeric values starting with the 1stof March as first day of the nesting season.The latest day of nest completion was day 104(12thJune)and the latest day of checking a nest was day 128(6thJuly),resulting in a 128-day nesting season comprising 127 daily intervals.We added additional variables,as described before,to the data set.Since batteries and storage cards of cameras needed to be checked more or less frequently during the observation period,depending on surrounding vegetation and placement of the cameras,we included an additional variable accounting for the number of nest checks in terms of human disturbance at the nesting site(nest checks).Data from 13 nests had to be excluded from nest survival analyses due to a lack of information or uncertainties about reproductive outcome.
2.3.Data analyses
We conducted all statistical analyses using the statistic software R(R Core Development Team,2020,Version 4.0.2) and R Studio (2021,Version 1.4.1106).Packages used included ‘lme4’ (Bates et al.,2015,2021),‘car’ (Fox et al.,2021),and ‘MuMIn’ (Bartoń,2020),as well as RMark(Laake,2013).
2.3.1.Hatching success
We defined hatching rate,also referred to as hatching success or fate,respectively,as the proportion of successful nests out of all nests monitored in the corresponding breeding season.Differences in hatching success along the years was analysed applying a linear regression (lm).For comparison of overall hatching success between first and replacement clutches we applied a Wilcoxon rank-sum test.We used a Pearson's Chi-squared test to test the dependency of the occurrence of foul eggs on female age.For this we classified female age into six groups (Table 1).Generalized linear mixed models(GLMM)and Generalized linear models(GLM)were used to analyze the effects of biotic and abiotic parameters on hatching success.We applied a GLMM to analyze the effects of female age,day of nest initiation and density of breeding pairs on hatching success of first clutches including the variable‘female’as random factor.Replacement clutches were excluded from this analysis because of allegeable differences between first and replacement clutches concerning time of nest initiation.For the analyses of general effects of habitat and weather parameters on overall hatching success we used a GLM.We built models step by step,eliminating non-significant variables until the best model was found.Model selection was based on the Akaike’Information Criterion(AIC;Burnham and Anderson,2002).
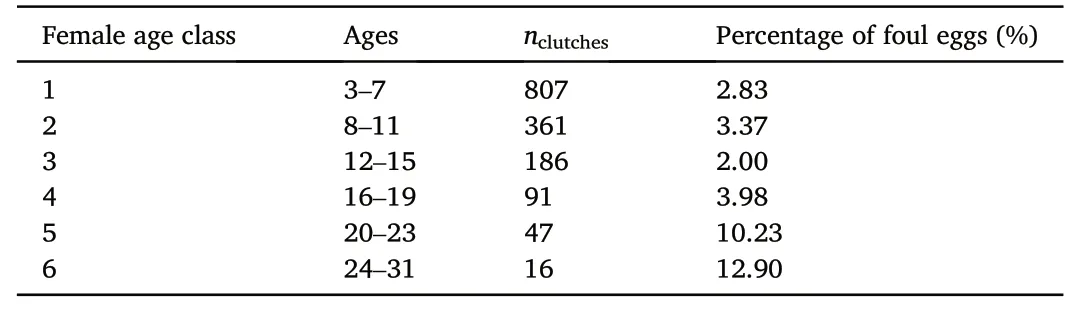
Table 1 Classification of female age and the occurrence of foul eggs along the age classes.
2.3.2.Nest survival
We conducted a complementary analysis of DSR of nests using RMark to investigate whether survival probabilities changed over the course of the nesting season depending on considered parameters.Applying the RMark package we modeled DSR of crane nests in R incorporating the following predictor variables: (1) year;(2) nest initiation;(3) habitat location;(4) clutch (i.e.,FC or RC);(5) mean temperature;(6) total sunshine hours during nesting season;and (7) nest checks.Dummy variables (i.e.,0 and 1 for each factor) were established for variables habitat location and clutch to simplify model building.We used logit link functions in all models and ranked nest survival models built in RMark based on Akaike's Information Criterion adjusted for small sample size(AICc),whereas we considered models with the smallest AICc,preferentially with ΔAICc <2 according to Burnham and Anderson(2002),to explain DSR best.
3.Results
3.1.Hatching success
We used data from 1316 clutches of Common Cranes (nFC=1124,nRC=192) to analyse hatching success.In general,hatching rate,referred to as hatching success,decreased over the course of the study period from~0.75±0.003(2002-2004)to~0.55±0.003(2018-2020;Fig.1;lm;R2=0.005,p=0.0124).
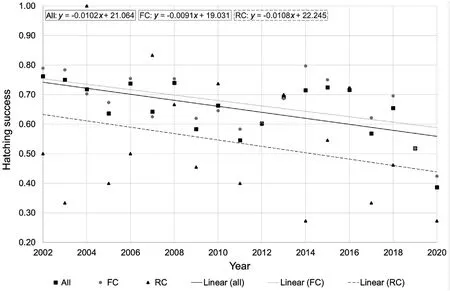
Fig.1.Hatching success of Common Cranes from 2002 to 2020 (nFC=1124, nRC=192).
Hatching success was significantly higher in FC compared to RC(Fig.1).Whereas 68.1 ± 1.4% of FC hatched successfully,a highly significantly lower percentage of only 52.6 ± 3.6% of RC hatched successfully(Wilcoxon rank-sum test,W=130027,p<0.001).
Hatching success showed an increasing trend with female age(Fig.2A;GLM;R2m=0.028,R2c=0.039,EST=0.034,SE=0.017,z=1.990,p=0.0466).However,the occurrence of foul eggs increased significantly with increasing female age as well.Probability of occurrence of foul eggs was highest in females aged 20 years and older(Pearson's Chi-squared test,χ2=41.384,df=10,p<0.001;Table 1),whereby the probability of occurrence of foul eggs did not differ between FC and RC(Pearson's Chi-squared test,χ2=0.4031,df=2,p=0.8175).Between 2002 and 2020 we recorded a total of 58 foul eggs including 23 clutches with foul eggs only and 35 clutches where one of two eggs was considered foul.
Earliest nest initiation was observed in 2016 as one crane pair started nesting on the 29thof February,whereas the latest initiation of a FC was documented in 2018,starting incubation on the 25thof May.On average,incubation started around the 6thof April (±11 days) and did not vary across different habitats nor did it shift over the course of the years,considerably.Day of nest initiation had a highly significant effect on hatching success of FC(Fig.2B;GLM;R2m=0.028,R2c=0.039,EST=-0.023,SE=0.007,z=-3.488,p<0.001).Hatching success decreased with delayed nest initiation (Fig.2B).Furthermore,increasing numbers of breeding pairs within a 2-km radius reduced hatching success significantly (Fig.2C;GLM;R2m=0.028,R2c=0.039,EST=-0.051,SE=0.023,z=-2.219,p=0.0265).Breeding pair density increased slightly,but statistically not significantly,over the course of the study period(lm;p=0.09),from an average of 4.8 pairs nesting within a 2-km radius around each nesting site in 2005 to an average of 6.1 pairs in 2018.Neither of the three factors were found to be linked to each other.
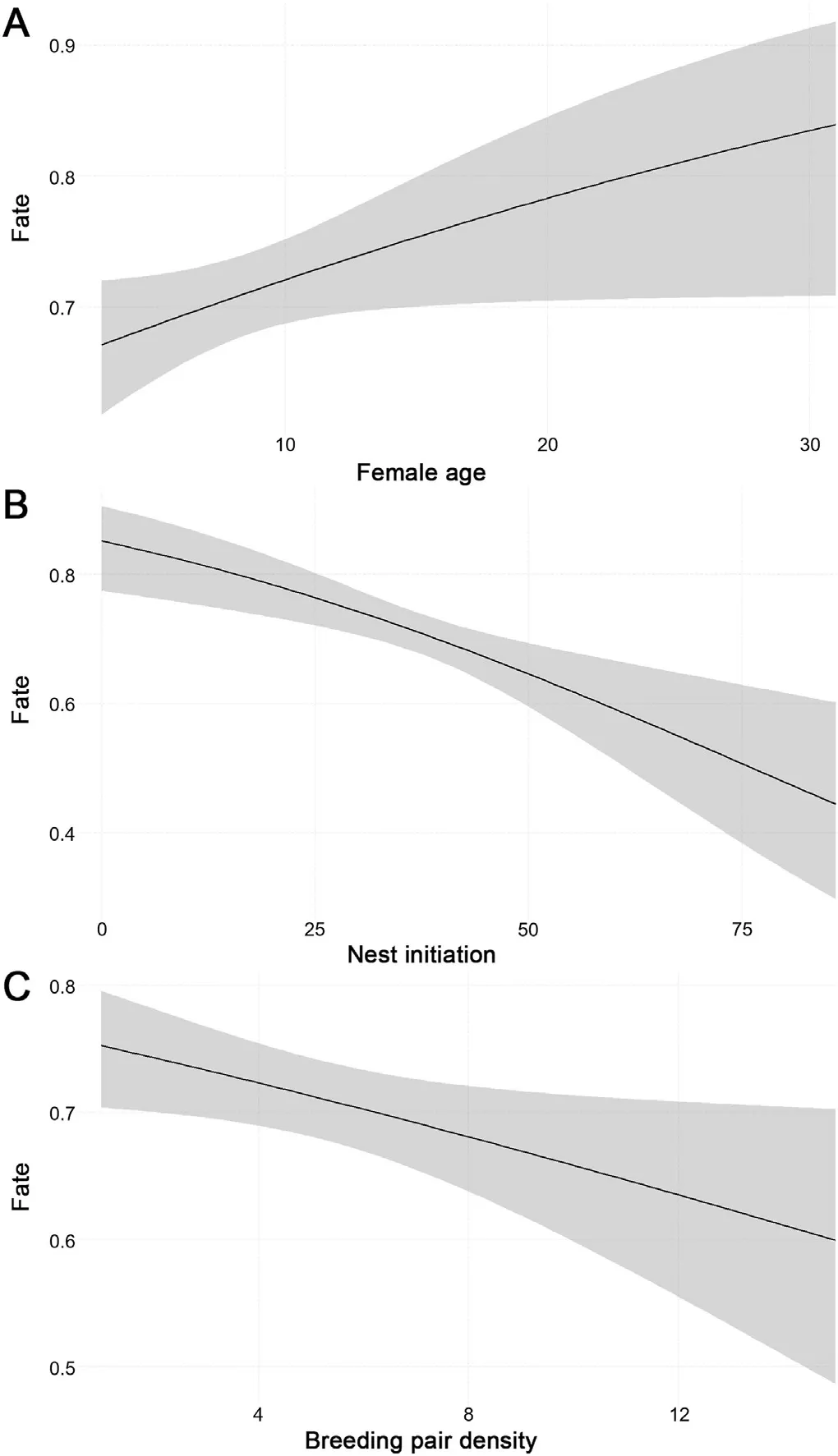
Fig.2.Effects of age (A),time of nest initiation (B) and breeding pair density(C) on hatching success.
Analyses of Pearson's correlation between abiotic parameters showed weak positive but insignificant correlations between years and mean temperature during breeding period(r=0.130),total sunshine duration(r=0.074) and amount of precipitation during preceding winter (r=0.379).Additionally,harshness of winter was weakly negatively correlated with years (r=-0.228).Correlation analysis was conducted to determine which variables might be responsible for the slight decrease in hatching success over the years,even though no direct effect of parameters on hatching success could be detected during modelling analyses.
Confirming the statement made by Mewes (2010) for the area Mecklenburg-Western Pomerania and Brandenburg in general,proportions of the three different nesting habitat locations changed significantly over the course of the study period within Goldberg (Fig.3).Whereas the proportion of open landscape nesting habitats increased(GLM;R2m=0.033,R2c=0.033,EST=0.086,SE=0.015,z=5.885,p<0.001),proportions of woodland(GLM;R2m=0.010,R2c=0.010,EST=-0.042,SE=0.013,z=-3.304,p<0.001)and intermediate(GLM;R2m=0.006,R2c=0.006,EST=-0.038,SE=0.015,z=-2.544,p=0.011) nesting habitats decreased.However,habitat location had no effect on hatching success and weather parameters did not affect hatching success,either(Table 2).
3.2.Nest survival
In total,we used 144 data sets from camera traps installed at cranes’nesting sites to model DSR,of which 80 hatched successfully (55.56%)and 64 failed due to predation (44.44%).The best model explaining variation of DSR included the variables day of nest initiation and number of nest checks during incubation period (Table 3).Weather parameters and habitat location as well as year did not have an impact on DSR.
Overall DSR as calculated in the best model was 0.978±0.003.Time of nest initiation(βNInit=-0.017±0.007;95%CI=-0.031 to-0.002)and number of nest checks (βNInit=0.697 ± 0.200;95% CI=0.305 to 1.090) both affected DSR (Table 4).Predicted DSR was negatively affected by delayed nest initiation as DSR decreased from 0.989±0.003 to 0.951±0.012 with delayed nest initiation,whereas DSR of nests that were checked more often(0.995±0.003)was slightly higher compared to DSR of nests checked only once (0.967 ± 0.007).However,survival estimates based on nest checks were not considered representative,since the data was assumed to be biased.Successful nests were observed for a longer period of time and therefore required multiple checks,whereas nests failing during early incubation stages were checked only once.
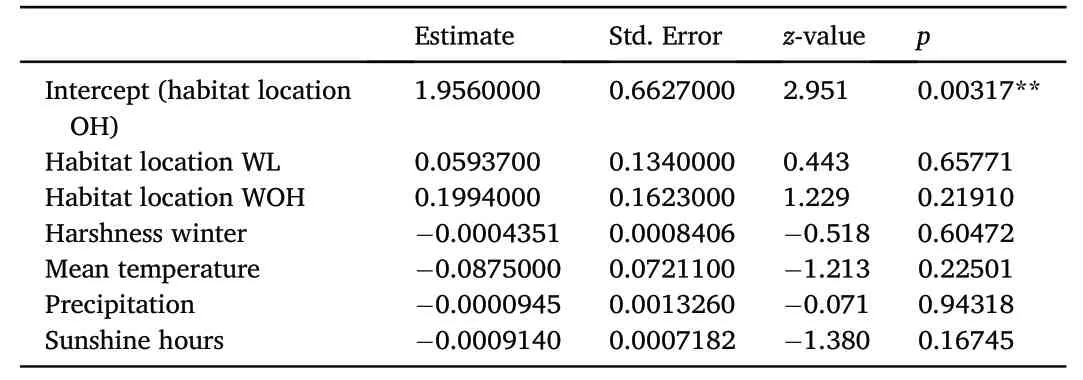
Table 2 Effects of environmental aspects on hatching success.
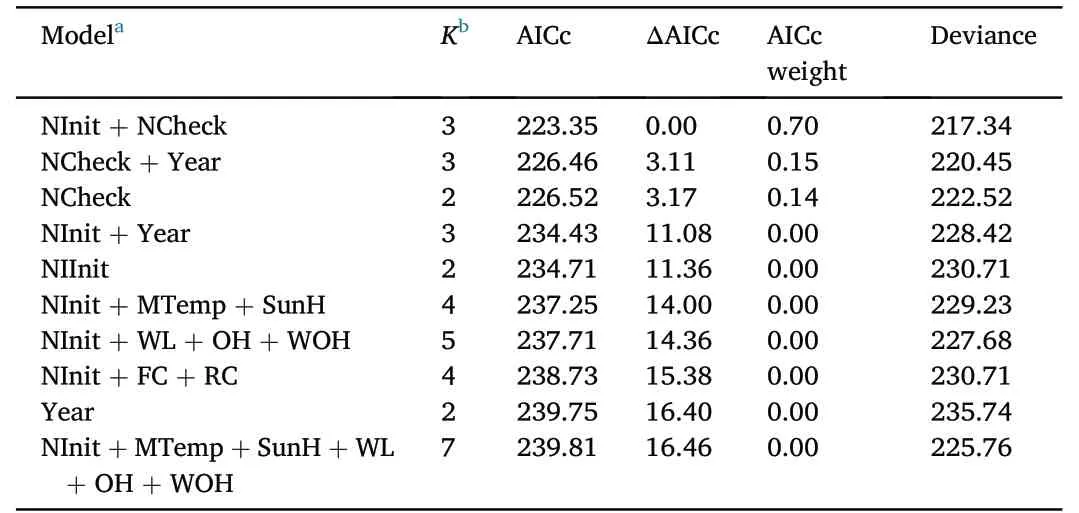
Table 3 Top ten daily nest survival models for Common Cranes,ranked by AIC.
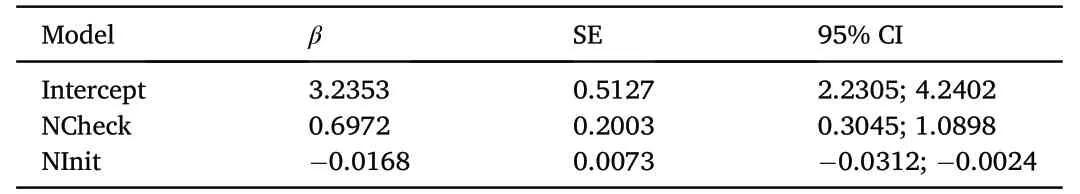
Table 4 Estimated beta coefficients (β) from the top-ranked DSR model for Common Cranes: S (NCheck+NInit)a.
4.Discussion
Our study confirmed observations by Mewes (2014a) that hatching success of Common Cranes decreased over the last decades and revealed the importance of age,day of nest initiation,and population density for the reproduction of the species,which had not been investigated before.
Our results,that hatching success increases with female age,corresponds to previous studies of other long-living bird species(Hernández et al.,2017;Nisbet et al.,2020;Wlodarczyk and Minias,2020).Higher age is associated with greater experience,i.e.longer reproductive history including a higher number of past reproductive experiences,resulting in higher reproductive success(Van Heezik et al.,2002;Forschler and Kalko,2006;Wlodarczyk and Minias,2020).Sensitivity towards predators might decrease with experience,resulting in higher aggressiveness and therefore successful defence of clutches(Møller and Nielsen,2014;Solovyeva et al.,2019).However,there has been a lack of studies on cranes’ behaviour,especially during the incubation stage,as well as age dependent changes in behaviour and physiology over time.Whether the population or age structure,which could belinked to changes in reproductive success,has changed over the years is unknown.We believe the more frequent occurrence of foul eggs in females aged 20 years and older to be a senescence effect.So far,senescence effects in cranes are only known in terms of decreases in egg size with increasing female age (Mewes,2020),but senescence effects are largely understudied in female birds (Nussey et al.,2013;Lemaître and Gaillard,2017;Assersohn et al.,2021),and true reasons for hatching failure,i.e.infertility or embryo mortality,in this study remain unknown.However,this did not affect overall hatching success,suggesting continuously good breeding performance in terms of incubation and protection of the clutch at higher ages compensating for potentially decreased female fecundity.Additionally,we have to consider that our data only reflect effects of females on hatching success,whereas data for males is missing.Possible negative effects of decreased sperm fecundity on reproductive outcome in older males (Lemaître and Gaillard,2017)can not be ruled out in this study.In birds,males often play a major role in the protection of clutches and offspring,contributing essentially to successful reproduction (Piper et al.,2017).In Common Loons (Gavia immer) males were shown to contribute to increases in hatching success with age over the entire life-span(Piper et al.,2017)and therefore effects of males on reproductive outcome,e.g.age and reproductive history,should be included in future investigations to expand knowledge on the breeding biology of cranes.

Fig.3.Changes in the proportions of nesting habitats in northern Germany from 2002 to 2020.
We also found delayed nesting initiation to be associated with lower hatching success,whereas age was not found to have an effect on the timing of nest initiation.Results of nest survival analysis confirm findings,that early nest initiation favor nest survival and therefore hatching success.Positive effects of early nest initiation on reproductive success have been observed for many bird species,especially long-living species(Nisbet et al.,2002;Vergara et al.,2007;Clark and DiMatteo,2018),accompanied by an age dependency of timing of nest initiation(Devries et al.,2008;Claassen et al.,2014).An early start of reproduction offers several advantages,such as the occupancy of the best nesting habitats,sufficient food availability or an extended time window for raising young or producing replacement clutches (Verhulst and Nilsson,2008).Additionally,increasing evaporation due to increasing temperatures over the years,which has already been predicted and proven for Europe (Briffa et al.,2009;Teuling et al.,2019;Mozny et al.,2020),might have a considerable impact on water availability.Even though we found no direct evidence to state that weather parameters affected hatching success,a correlation can not be ruled out.With rising temperatures over the nesting season,evaporation increases as well,causing a reduction of water levels.Decreasing water levels grant easier access to cranes'nests for predators,increasing the probability of nest failure.Considering this,negative effects of delayed nest initiation also explained reduced hatching success in RC's compared to FC's.
Our data do not support the hypothesis of a correlation between age and timing of nest initiation in Common Cranes,resembling findings by Clark and DiMatteo (2018) for American White Pelicans (Pelecanus erythrorhynchos).Behavioural traits could be linked to timing of nest initiation,instead,like it was the case for Goshawks(Accipiter gentilis),in which individuals initiating incubation early in the breeding season were found to be more aggressive or bolder than others,which positively affected reproductive success (Møller and Nielsen,2014).Since cranes show a similar territoriality,this might also be applicable to cranes,but evidence is missing due to a lack of behavioural studies.
Cranes are highly territorial species reacting sensitively to intraspecific competition (Johnsgard,1983) and our results support the hypothesis of density dependent declines of reproductive success.Increasing numbers of crane breeding pairs within a 2-km radius around a nesting site caused the reproductive success to decrease significantly.High population densities intensify competition for territories,good nesting sites and food sources (Fernandez et al.,1998).Marking and defending territories is energetically costly and time consuming,preventing birds from taking proper care of the clutches and offspring(Bretagnolle et al.,2008).Additionally,high breeding densities increase the risk of intraspecific predation,which has been documented for Common Cranes,recently(Barwisch et al.,2019).Optimal territory size mainly depends on the habitat and resource availability (Johnsgard,1983) and ranges from 0.2 km2to 17 km2within the family Gruidae(McCann and Benn,2009;Månsson et al.,2013;Miller and Barzen,2016).However,we suggest intraspecific interactions associated with competition for good nesting sites to negatively affect hatching success rather than limited food availability,since adult cranes show high mobility during the incubation stage.Home ranges of Common Cranes in northern Germany were found to range between 0.2 and 1.3 km2(Nowald,2003),but we have to take into account that this resembles the home range of families including less mobile pre-fledged chicks.Home ranges tend to be larger during incubation stages,decreasing competition for food sources(Maxon,1974).For that reason and referring to territory sizes found in other crane species,we assumed a larger territory size for the species at the upper end of approximately 12.5 km2(i.e.2-km radius around the nesting site).So far,sufficient data,i.e.information on population density complemented by detailed data on reproductive success,from other areas for comparisons are missing.To substantiate the negative effects of high population densities on reproductive success comprehensive breeding site monitoring programs should be established in different areas across the geographic range of Common Cranes.
Our analyses did not account for interactions between population density and habitat parameters,which should be taken into account in future studies to investigate the effect of nesting habitat on territory size and habitat capacities.Regardless of this,hatching success did not vary across different nesting habitats in our study area.Different than expected,hatching success of cranes nesting within intensively used agricultural landscapes was the same as for those using wooded areas.Prior investigations between 2003 and 2012 in the same study area by Mewes and Rauch (2012) stated a significantly lower hatching success for pothole wetlands located within agricultural fields compared to other habitats.Since the proportion of breeding pairs using agricultural areas for nesting further increased after 2012 cranes might have further adapted to the conditions as well,accounting for an approximation of hatching success.Converse effects were observed for White-naped Cranes(Grus vipio)and Florida Sandhill Cranes(G.canadensis pratensis).Cranes using nesting sites located within managed pastures or suburban areas showed lower reproductive success compared to cranes nesting within undisturbed habitats (Toland,1999;Bradter et al.,2005).A possible explanation for the results found in our study could be the cranes’ability to adapt quickly to disturbances caused by agricultural activities.Another closely related species,the Sarus Crane(G.antigone)was found to successfully adapt to agricultural activities within rice paddies (Sundar,2009).
Based on the high nesting site fidelity of Common Cranes (Mewes,2017),there is no indication that cranes change their nesting site after unsuccessful reproduction.However,single females could not be detected in one or more years but showed up one or several years later.Information on the whereabouts and reproduction of these individuals,when their presence could not be proven,remain unknown.To complement our results,long-term data from GPS-tracked individuals should be taken into account to analyze the impact of non-breeding periods and current reproductive success on future habitat selection and reproductive performance of an individual.Such information is valuable for the estimation of fitness aspects,life history traits and the understanding of a specie's population dynamics (Kennamer et al.,2016).
Additionally,differences in predator abundance across the study area might be accountable,but details on predation rates and major predators within the area are missing.In general,population numbers of common predator species,e.g.Wild Boars (Sus scrofa;Gethöffer et al.,2007),North American Raccoons (Procyon lotor;Michler,2016) or Raccoon Dogs (Nyctereutes procyonoides;Drygala et al.,2000),increased over the last decades within Germany,which might contribute to decreases in the hatching success of Common Cranes,but we lack details on spatial abundances within our study area.Despite-or perhaps precisely because of -the cranes’adaptability,results of our analyses do not fully explain decreases in hatching success over the last years,which is also confirmed by model statistics.Analysed variables could only explain a small proportion of variance in hatching success.However,our results clearly indicate the importance of population dynamics and individuals features of crane breeding pairs.
The ability to successfully raise juveniles is another crucial factor determining reproductive success(Pakanen et al.,2020).We hypothesize that habitat quality concerning food availability varies markedly between natural and managed or intensively used areas,respectively.Protein-rich food is essential for juvenile growth (Park et al.,2001).Comparative studies of reproductive success between natural and agriculturally used habitats showed lower chick survival and reduced growth rates of juveniles in agricultural landscapes of Europe (Britschgi et al.,2006;Kentie et al.,2013).Insect abundance is very limited in intensively used farmland(Hendrickx et al.,2007)and therefore adults might not be able to provide sufficient food for successfully rearing the young.Further investigations are necessary to be able to objectively evaluate habitat quality in terms of raising young and overall reproductive success of Common Cranes.
5.Conclusion
Cranes have shown to be relative flexible in the use of habitats within disturbed agricultural areas.However,our study area in Germany represents a unique region within the distribution range of Common Cranes showing such an increase in nesting populations,despite the great proportion of intensive agriculture.Additionally,being long distance migrants,crane populations are exposed to other challenges along migratory routes and wintering areas.For these reasons,it is necessary to be aware of the effects of high densities on reproduction success,without relaxing their protection status in other regions,where human disturbances are higher and the crane populations are more negatively affected and still in low numbers.
Our study was the first to investigate effects of age and population density on the reproductive outcome of Common Cranes on a larger scale using long-term monitoring data.The possibility to follow the reproductive history of single females for up to 30 years is very unique for studies on wild birds.Results reveal the importance of information on the age structure of Common Cranes for the understanding of mechanisms behind successful reproduction.Based on our results we see an urgent need to further investigate the reproductive success and breeding biology of Common Cranes nesting within anthropogenically shaped landscapes.Increasing population densities within northeastern Germany suggest a good capability of cranes to adapt to changing landscapes at first,but decreasing hatching success may lead to a reduction of the population in the long term.Sufficient data on reproductive success,i.e.fledging chicks,and recruitment rates are missing,making predictions for future developments of the population as well as the establishment of adequate conservation measures difficult.Additionally,behavioural studies are needed to evaluate the adaptability of cranes to disturbances and changing landscapes.
Author contributions
WM and ASO conceived the idea of the study.WM collected data on hatching success in the field alongside others (see Acknowledgements).IB contributed to the data collection between 2016 and 2021,collected data for analysing nest survival,and processed raw data.IB and ASO analysed and interpreted the data regarding hatching success of Common Cranes and nest survival of clutches including statistical analyses.IB led the writing of the manuscript.ASO and WM contributed significantly to reviewing and editing the manuscript.All authors read and approved the final manuscript.
Ethics statement
This study was purely observational,disturbances were kept as low as possible and no harm was done to individual birds or clutches.Permissions to visit crane nesting sites were present throughout the study period.
Declaration of competing interest
All authors declare that they have no conflicts of interest.We confirm that we have no affiliations with or involvement in any organization or entity with any financial interest,or non-financial interest in the subject matter discussed in this manuscript.
Acknowledgements
We are grateful to Moriz Rauch and Volker Mewes for supporting data collection in the field over many years and additionally,we thank Prof.Dr.Martin Haase and Dr.Silke Fregin for open and constructive debates throughout the project.Acquisition of camera traps used for collecting data on nest survival was partly funded by the German Ornithologists'Society(DO-G).
杂志排行

Avian Research的其它文章
- Functional and phylogenetic structures of pheasants in China
- Thermoregulatory function and sexual dimorphism of the throat sack in Helmeted Guineafowl (Numida meleagris) across Africa
- Distribution pattern and driving factors of genetic diversity of passerine birds in the Mountains of Southwest China
- Multiple lines of evidence confirm that the critically endangered Blue-crowned Laughingthrush(Garrulax courtoisi)is an independent species
- Corrigendum to “Multiple lines of evidence confirm that the critically endangered Blue-crowned Laughingthrush (Garrulax courtoisi) is an independent species” [Avian Res.13 (2022) 100022]
- Altitudinal seasonality as a potential driver of morphological diversification in rear-edge bird populations