Interaction network between frugivorous birds and zoochorous plants in cloud forest riparian strips immersed in anthropic landscapes
2022-10-03OmrHernndezvilJvierLordeVinicioSosCeciliCstelzo
Omr A.Hernández-Dávil ,Jvier Lorde,* ,Vinicio J.Sos,** ,Cecili Díz-Cstelzo
a Instituto de Ecología,A.C.,Red de Ecología Functional,Carretera Antigua a Coatepec,No.351,El Haya,Xalapa,Veracruz C.P,91073,Mexico
b Instituto de Ecología,A.C.,Red de Interacciones Multitróficas,Carretera Antigua a Coatepec,No.351,El Haya,Xalapa,Ver.C.P,91073,Mexico
Keywords:Frugivory Keystone species Riparian corridors Seed dispersal Vegetation corridors
ABSTRACT Worldwide,tropical montane cloud forest is one of the most important and biodiverse ecosystems;however,it is also one of those most threatened by anthropic activities.These activities lead to a fragmented,deforested landscape with narrow riparian forest strips immersed in an agricultural matrix dominated by pastures.Here,we characterize the interaction network between frugivorous birds and zoochorous plants in riparian strips of cloud forest in deforested landscapes of Central Veracruz,Mexico.To characterize the network of this mutualistic interaction,we estimated network-and species-level metrics using the Bipartite R package.Nestedness,modularity and robustness were used to describe network structure.Centrality measures of degree,closeness,betweenness centrality and their relative contribution to nestedness were used to determine the importance of each bird/plant species to the network’s structure.This interaction network has 24 species of birds and 30 species of plants,with low connectance (0.11),low nestedness (11.53),and intermediate but not significant modularity(0.49).The bird species most important to network stability were Chlorospingus flavopectus,Myadestes occidentalis,and Catharus mexicanus.The most important plants were Conostegia xalapensis, C.arborea,and Rubus ulmifolius.Network robustness varied from 0.36 to 0.86 and its stability is compromised when species of birds or plants with the highest values of centrality are removed,with plant removal more detrimental.Riparian strips of cloud forest crossing deforested areas maintain a relatively rich set of birds that disperse the seeds of many forest plants,thus they are crucial to the preservation of this mutualistic network in anthropic landscapes.Network stability is severely undermined by the loss of any of the few species -whether birds or plants -with high centrality values.The most important plants for this stability are pioneer tree and shrub species that provide food for several bird species,and they are also crucial to cloud forest regeneration.A worrisome finding is that some of the bird species most important to network stability are also among the most sought-after as ornamental birds by illegal collectors in the region.
1.Introduction
Tropical montane cloud forests are one of the most threatened ecosystems worldwide,due mainly to their conversion into crop fields and pastures (Bruijnzeel et al.,2011;Hernández-Pérez et al.,2022).It is estimated that cloud forests cover an area of 250,000 km2worldwide,which represents 0.14% of the Earth’s terrestrial surface and 1.14% of the total area covered by tropical forests.Of the total area of cloud forest,41%is in the Americas,where it represents 6.3% of the area covered by tropical forests (Bruijnzeel et al.,2011).Approximately 3700 species have been recorded in cloud forests,including birds,mammals,amphibians,and tree ferns,half of which are endemic to this type of forest(Karger et al.,2021).Neotropical cloud forest reaches its northern limit in Mexico,where more than half of its original cover has been replaced by different types of land use,mainly crops and pastures(Muñoz-Villers and López-Blanco,2008;Ponce-Reyes et al.,2012;Hernández-Pérez et al.,2022).In this context,natural remnants of cloud forest within transformed landscapes play an important role in maintaining biodiversity (Toledo-Aceves et al.,2014).Riparian forest strips crossing agricultural or man-modified areas represent forest remnants that could be important for biodiversity conservation since they provide food,refuge,and habitat corridors for several species of flora and fauna in drastically simplified and impoverished sites (Lees and Peres,2008;Carrasco-Rueda and Loiselle,2019;Hernández-Dávila et al.,2020).
Habitat loss due to agricultural expansion leads to a decrease in the richness and abundance of native species,the local extirpation or global extinction of populations,and changes in species composition and their interactions,causing ecosystem degradation with dire consequences for biodiversity (Laurance et al.,2011;Hagen et al.,2012).Ecosystem function and community dynamics depend on biotic interactions between species(Hagen et al.,2012).In particular,mutualistic interactions between plants with fleshy fruit and vertebrate frugivores(mainly birds and bats)play a key role in the functioning of tropical ecosystems since 65-90% of their woody plant species depend on these vertebrates for seed dispersal(Howe and Smallwood,1982;García et al.,2010;Jordano et al.,2010).Mutualistic interactions can be analyzed using a network approach to understand some of the drivers that organize communities,as well as the role of these interactions in maintaining biodiversity and in the provision of environmental services (Bello et al.,2015;Peres et al.,2016;Morán-López et al.,2020);knowledge that can be applied to ecosystem management and restoration (Peters et al.,2016).An interaction network is depicted as a graph with nodes and edges,where nodes represent different interacting species and edges represent the interactions between these species (Dehling,2018;Delmas et al.,2019).Some parameters of network structure,such as connectance,nestedness,and modularity,have been used to describe the topology of the interacting community (Jordano et al.,2009;Dehling,2018).Similarly,centrality measures of the network like degree,closeness and betweenness centrality,as well as the contribution made to nestedness by each node,are useful for assessing the relative importance of each species to the whole structure of the interaction network (Martín-González et al.,2010;Dehling,2018;Morán-López et al.,2020).This information can be used to assess the relative importance of each of the interacting species and the weight or relevance of the different interactions to community dynamics and stability.For frugivory interactions between birds and plants,this approach can be used to weight the importance of interacting species with respect to seed dispersal and ultimately,forest regeneration(Mello et al.,2014;Escribano-Avila et al.,2018;Messeder et al.,2020).
In the cloud forest fragments that remain in the anthropic landscapes of Mexico,some aspects of the frugivory and seed dispersal interaction have been described for both bats (Saldaña-Vázquez et al.,2010) and birds (Hernández-Ladrón De Guevara et al.,2012;Baltazar,2014;Pérez-Cadavid et al.,2018).However,only one study in degraded cloud forest landscapes has used a network approach to assess the effects of anthropic disturbance on this mutualistic interaction.That study,carried out in a peri-urban park on the outskirts of Xalapa,in Veracruz,Mexico recorded 23 plant species interacting with 29 bird species(Salazar-Rivera et al.,2020),in spite of the highly modified landscape.
In our study site the deforested areas,mostly converted into extensive pastures to raise cattle,are crossed by long,narrow tracts of arboreal vegetation left uncut on both sides of permanent rivers in order to protect the riverbank.These riparian forest strips could be extremely important to the local flora and forest fauna within this highly degraded landscape,and might function as arboreal corridors that connect forest fragments and other arboreal patches(i.e.,living fences,isolated trees,etc.)that are currently separated (Hernández-Dávila,2013;Hernández-Dávila et al.,2020).In a region like this,we would expect that the number of birds and plants interacting in the riparian strips would be low,forming extremely poor,simple networks.The long-term maintenance of native biodiversity in any transformed landscape is in jeopardy if the bird-plant interaction network is impoverished and has a low degree of interaction.As such,the restoration of cloud forest in these landscapes would be impaired specifically due to seed dispersal failure.In this context,our goals were:1)to characterize the frugivory network between birds and plants in the riparian strips of cloud forest left along the rivers that cross man-made pastures;2) to identify the most important bird and plant species in terms of their contribution to network stability in these simplified ecosystems;and 3)to assess quantitatively the effect of potential changes in standard network-and species-level metrics of the interaction network under different simulated scenarios of species loss that could simplify further the current network.
2.Materials and methods
2.1.Study area
The study area is located in the upper basin of the La Antigua River in Central Veracruz,Mexico(19°32′32′′-19°22′09′′N,97°06′52′′-96°56′48′′W).The climate is humid temperate with a mean annual temperature of 18°C and 1500 mm of annual precipitation.The vegetation is composed of remnants of tropical montane cloud forest where tree species such asPlatanus mexicana(Platanaceae),Quercus lancifolia(Fagaceae),Clethra macrophylla(Clethraceae),Liquidambar styraciflua(Altingiaceae),Styrax glabrescens(Styracaceae),Carpinus tropicalis(Betulaceae) andTurpinia insignis(Staphyleaceae) dominate.For a detailed description of the vegetation structure and floristic composition of these remnants see Williams-Linera et al.(2013)and Hernández-Dávila et al.(2020).In this study,we use the term riparian strips to denote the cloud forest vegetation left along the riverbanks by local farmers to protect the river channel when converting their plots to crop-fields or pastures.These forest strips are usually 5-m wide on each side of the river and are dominated by 20-30 m tall canopy trees of Mexican sycamore (Platanus mexicana;Hernández-Dávila et al.,2020) and are rich in tree species native to the original cloud forest.It is important to emphasize that owing to agricultural management practices and extensive deforestation in the region,these riparian strips are immersed in an agricultural matrix with different types of land use,dominated by man-made pastures for cattle grazing,with some small crop fields and patches of secondary vegetation(Appendix Fig.S1).
2.2.Bird capture and seed collection
We selected six tracts of riparian strips for sampling in a region dominated by extensive cattle pastures.Within each of these strips,we erected nine mist nets(9×2.7 m)at ground level and suspended another six mist nets (6×2.7 m) from tall trees at 12 m above the ground.The latter were used to capture species that use the middle to upper forest strata and that usually do not visit the understory.The nets were opened twice a day from 06:00 to 11:00 and from 16:00 to 18:00,monthly during three consecutive days,avoiding rainy days.From August 2016 to July 2017,we sampled one of the six selected strips every month,so each one was sampled twice over a year.Thus,we sampled both the resident and migratory avifauna of the site to ensure acceptable sampling completeness (see below).Plastic sheets 1-m wide were placed on the ground beneath each mist net to collect any feces dropped by birds at the moment they were caught in the net(details in Hernández-Dávila et al.,2015).Additionally,to increase the chances of collecting defecated seeds,each bird captured was placed in a cloth bag for at least 20 min,and later released in situ.The birds were identified using the field guides of Sibley (2014) and Howell and Webb (1995),and following the nomenclature of the American Ornithological Society (Chesser et al,2021).With these field data we constructed a single bird-plant network to represent the frugivory network associated with forested riparian strips at the landscape level.
2.3.Recording frugivory interactions
For seed identification,we used reference seed collections curated by several researchers at the Instituto de Ecología,A.C.(INECOL) and the Instituto de Biotecnología y Ecología Aplicada(INBIOTECA)of the Universidad Veracruzana.Bibliographic references were also used (e.g.,Ibarra-Manríquez et al.,2015).Additionally,ripe fleshy fruit were collected from the sampling sites and their surroundings in order to extract their seeds and create a reference collection for the zoochorous plant species of the cloud forest.The botanical specimens collected were identified with the help of the personnel of the XAL Herbarium at INECOL.Plant species nomenclature follows Tropicos® (Missouri Botanical Garden,2020).
2.4.Data analysis
Our sampling effort for birds and plant species(seeds),as well as for the number of interactions was assessed using sample completeness curves in iNEXT (Hsieh et al.,2016).To analyze the frugivory interactions between plants and birds,a qualitative interaction matrix was built using all of the interactions recorded in the six riparian strips.In this matrix,bird species are the columns,plants are the rows,and each cell indicates whether the interaction was recorded(1)or not(0).
To analyze network structure,we calculated the following networklevel metrics.Connectance (C) is the proportion of recorded connections with respect to the total number of potential connections(Jordano,1987),based on formulaC=I/(A×B),whereI=the number of observed interactions,A=the number of bird species,andB=the number of plant species detected in the study.Nestedness (NODF) indicates the asymmetric structure of the interactions in the network,that is,whether there are species with a low degree of interaction with a subset of species that has a high degree of interaction(Bascompte et al.,2003).The estimation of NODF is based on overlap and decreasing fill(Almeida-Neto et al.,2008) and its value ranges from 0 to 100,where values close to 100 indicate highly nested networks.To determine the significance of nestedness we used null model II,in which the probability of one interaction is regarded as proportional to the total number of interactions between the bird and the plant species (Bascompte et al.,2003).Modularity(M)indicates whether or not the network is organized into distinguishable modules or compartments,that is,whether there is a group or subset of species that interact with each other more than would be expected by chance.Modularity was calculated using the Barber function and the simulated annealing (SA) algorithm to maximize the modularity function (Barber,2007).Significance was determined through a permutation test.Mranges from 0 to 1,where 1 indicates that the network is organized into modules and zero indicates an absence of modularity.Robustness(R)represents the stability of the network when a given set of species is eliminated from the recorded network(Bascompte and Jordano,2007).For this metric,we used two different methods based on species degree,eliminating species one by one from the highest to the lowest degree and vice versa.Simulated extinction scenarios(i.e.,removal from the network)for both plant and bird species based on these two methods were compared with a null model scenario based on the random deletion of species(Memmott et al.,2004).Rranges from 0 to 1,where values approaching 1 indicate greater network stability after species loss or removal(Burgos et al.,2007).Additionally,we estimated Horn’s niche overlap index,which indicates how much,if any,overlap there is among the bird species that feed on certain plant species or overlap among the plants with respect to the birds that disperse their seeds.Niche overlap ranges from 0 (no overlap) to 1 (complete overlap between species).
To analyze the relevance of each bird and plant species in the network structure,we calculated the following species-level metrics.Species degree (D) is the number of interactions per species (i.e.,the number of different plant species eaten by a bird species,and how many bird species defecated the seeds of a given plant).Closeness centrality(CC)measures the proximity of a node (bird or plant species) to all other nodes(Freeman,1978).High CC values imply that the bird or plant species interact with a high number of plant or bird species and,for this reason,can affect other nodes in the network (Martín-González et al.,2010;Delmas et al.,2019;Morán-López et al.,2020).Betweenness centrality(BC)measures the occurrence of a species in a connection between every pair of species,and thus describes the relevance of a node as a connector between different parts of the network(Freeman,1977).Species with a positive value (BC >0) are important for network cohesion (Martín--González et al.,2010;Delmas et al.,2019).Since our network was slightly nested,we also estimated the contribution to nestedness(CN)of each species(Saavedra et al.,2011),where species with positive values(CN >0)make a greater contribution than those with negative values.All network-and species-level metrics were estimated using the Bipartite package in R language(Dormann et al.,2008).
3.Results
Cumulative sampling effort was 1473 mist-net hours with a total capture of 273 birds belonging to 52 species(Table 1).Ninety-two of the captured birds,belonging to 24 species,defecated at least one undamaged seed,for a grand total of 6561 defecated seeds from 30 plant species.Sample completeness curves indicate that we recorded 92% of the estimated bird richness,87% of the estimated plant richness (i.e.,defecated seeds),and 46% of the potential interactions(Appendix Fig.S2).
3.1.Network-level metrics
The size of the interaction network was 24 bird species by 30 plant species (Fig.1),with a total of 81 different interactions observed.Network connectance was low,C=0.11,that is,11% of all potential interactions between the 24 birds and 30 plant species recorded.The interaction network was slightly but significantly nested(NODF=11.53;P<0.01) and had a moderate but not significant degree of modularity(M=0.49;P>0.05).Network robustness under the null random model eliminating birds wasR=0.66,while for plants it wasR=0.67.In the scenario in which species were eliminated from highest to lowest degree,robustness wasR=0.36 for birds andR=0.45 for plants.Eliminating species from the lowest to highest degree,robustness wasR=0.86 for birds andR=0.84 for plants.Extinction scenarios showed that,by eliminating 40% of the most connected species only 30-50% of the birds or plants species would remain in the impoverished network simulated this way.In contrast,by eliminating 40% of the less connected species,over 90% of interacting bird/plant species would remain in the network(Fig.2).Finally,the null model showed that when 40% of the bird or plant species were eliminated randomly from the network,approximately 80% of the bird and plant species would remain in the impoverished network.Niche overlap was weak among the species of each trophic group;0.10 for birds and 0.14 for plants.
3.2.Species level metrics
3.2.1.Birds
The bird species with the highest degree values wereChlorospingusflavopectus(D=10 different interactions),Myadestes occidentalis,Catharus mexicanus,Euphonia hirundinacea,andArremon brunneinucha(each withD=8).These birds also had the highest values of closeness centrality(CC;0.05 for each species).The species with the highest values for betweenness centrality (BC) wereTurdus grayi(0.30),Myiozetetes similis(0.17),C.mexicanus(0.12)andCh.flavopectus(0.11;Table 2).The bird species that made the greatest contribution to nestedness wereCh.flavopectus(CN=2.5),M.occidentalis(1.4),Melanotis caerulecens(1.2),E.hirundinacea(1.1),andC.mexicanus(1.1;Fig.3).
In the daytime she sat down once more beneath the windows of the castle, and began to card with her golden carding-comb; and then all happened as it had happened before
3.2.2.Plants
The plant species with the highest degree values wereRhamnuslongistyla(D=7 different interactions),Solanum aphyodendrum(D=6),Miconia glaberrima,Conostegia xalapensis,andTrema micrantha(each withD=5).Plant species with greatest CC values wereC.xalapensis,Rubus ulmifolius,andC.arborea(0.04 each).The highest BC values were obtained forR.ulmifolius(0.24),C.xalapensis(0.14),andC.arborea(0.10)(Table 2).Plant species that made the greatest contribution to nestedness wereC.xalapensis(CN=0.97),R.ulmifolius(CN=0.92),andAnthurium scandens(CN=0.90;Fig.3).
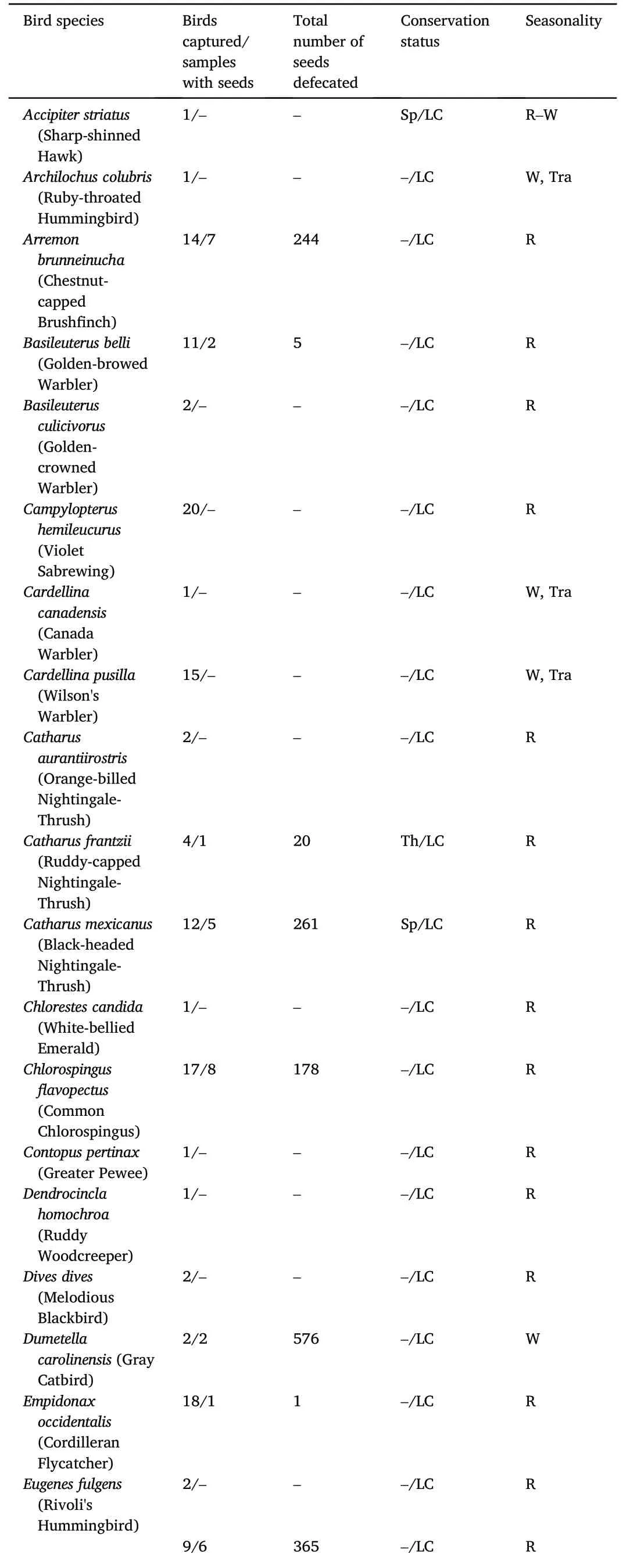
Table 1 Bird species in cloud forest riparian strips in Veracruz,Mexico,number of defecated seeds,bird conservation status and seasonality.
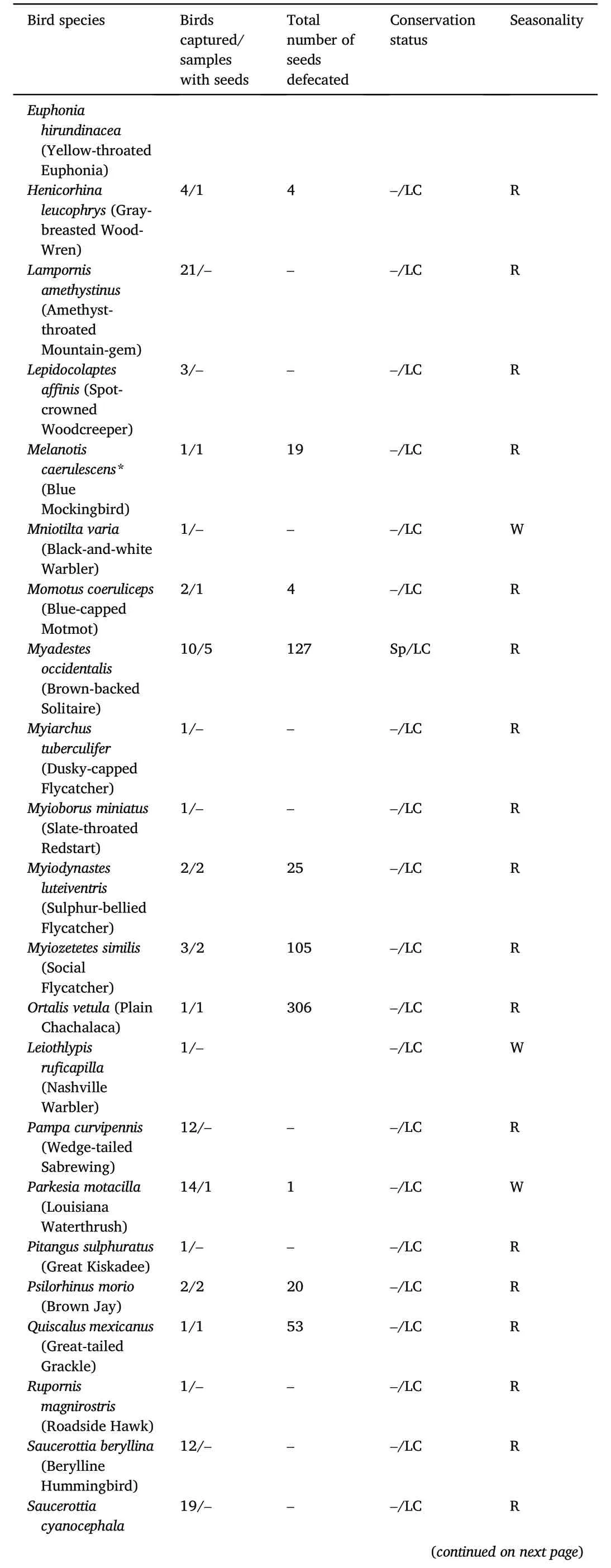
Table 1(continued)
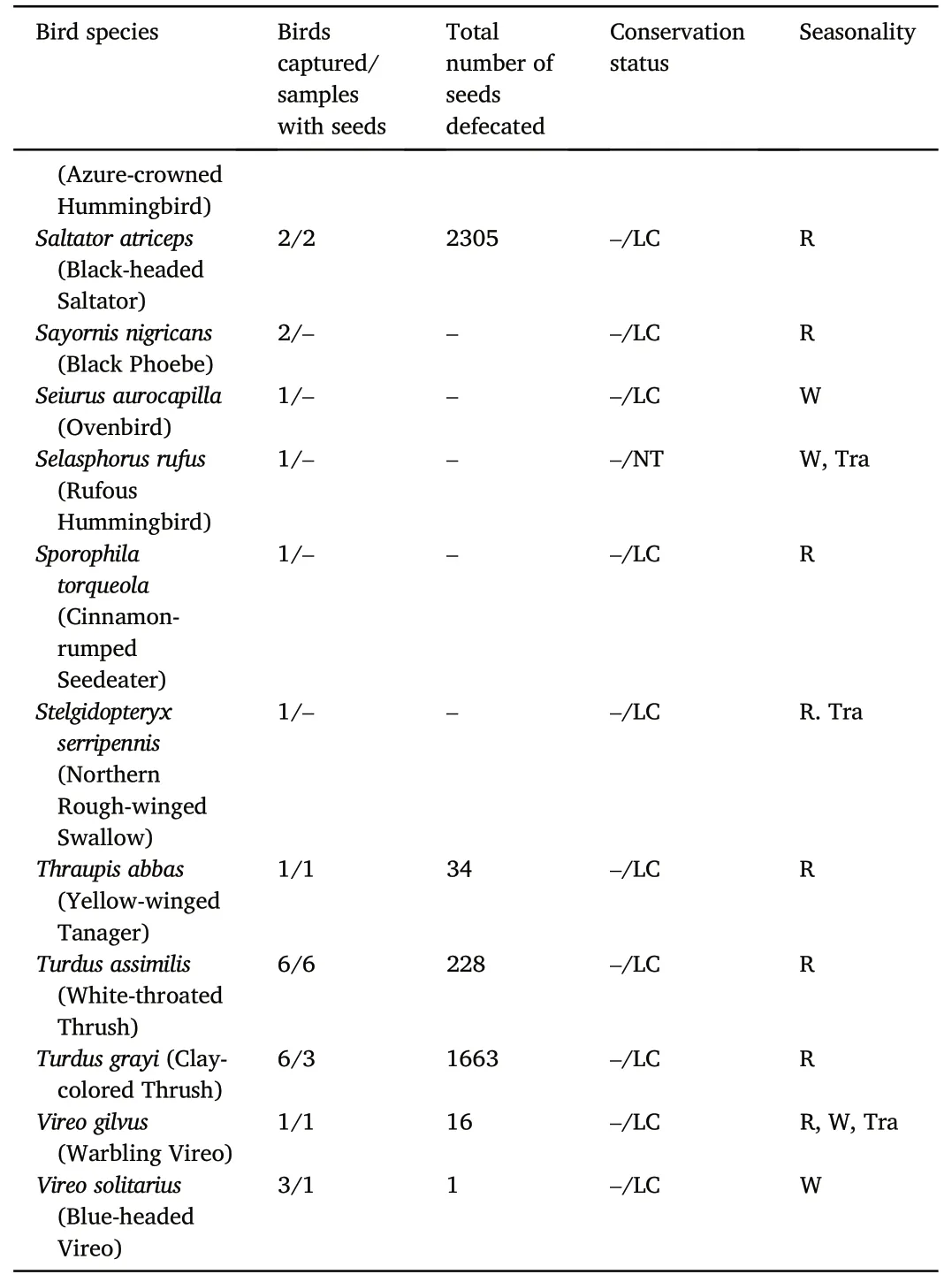
Table 1(continued)
4.Discussion
In highly fragmented tropical landscapes that are dominated by extensive agricultural areas,remnant arboreal elements composed of native forest species such as the riparian forest strips that cross agricultural areas play a vital role in maintaining biodiversity and ecosystem services.Our results show that,despite widespread deforestation,the interaction between frugivorous birds and zoochorous plants is still occurring in riparian strips of cloud forest crossing man-made pastures and involves many species of forest plants and birds.Our sampling was sufficient to account for the diversity of frugivorous birds and zoochorous plants present in the riparian strips,as shown by the completeness curves.
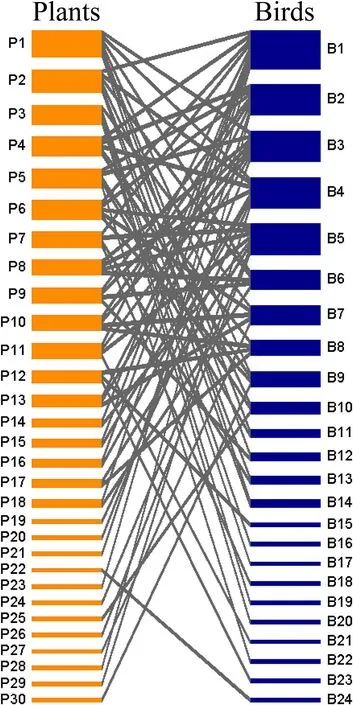
Fig.1.Qualitative interaction network between plants (30 spp.) and birds (24 spp.) in riparian strips of cloud forest in Veracruz,Mexico.The blue boxes represent bird species and the orange boxes,plant species.Box thickness indicates species degree.Species names for each group are listed in Table 2.(For interpretation of the references to color in this figure legend,the reader is referred to the Web version of this article.)
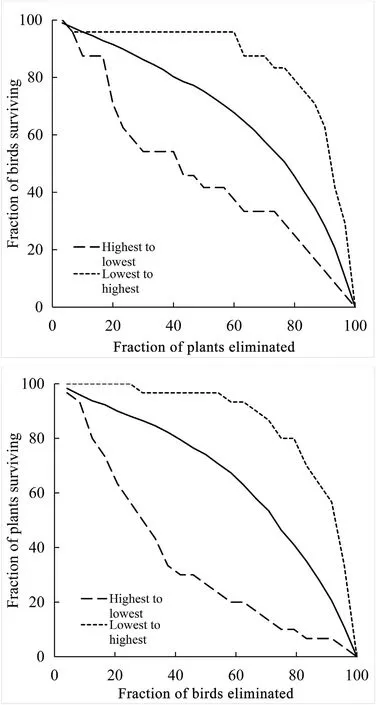
Fig.2.Scenarios of extinction/deletion of birds (up) or plants (down) in riparian strips in Veracruz,Mexico using two methods,from highest to lowest degree species (dashed line) and from lowest to highest degree species (dotted line).The null model is shown as a continuous line.The x axis represents the fraction of eliminated bird or plant species and the y axis represents the fraction of bird or plant species that continue to exist.
The number of bird and plant species recorded interacting in narrow riparian strips seems to be low when compared to similar studies carried out in less disturbed Neotropical forests,in which the number of interacting species was seven to 128 bird species and 100 to 178 plant species(Palacio et al.,2016;Emer et al.,2020;Messeder et al.,2020).However,our results were similar to those of other studies carried out in much larger fragments of old-growth protected cloud forest in anthropic landscapes in Central Veracruz,Mexico,where the interaction network had 23-33 bird species and 17-33 plant species (Hernández-Ladrón De Guevara et al.,2012;Baltazar,2014: Salazar-Rivera et al.,2020).Most interaction network studies are based on the direct observation of birds feeding on the fleshy fruit of different plants,which has a greater potential to detect bird/plant interactions than field methods based on seeds defecated by birds captured in mist-nets.In other words,our mist-netting method very likely underestimated the actual size of the network since the number of interactions recorded this way over a single year,is unlikely to include all actual interactions (García,2016).However,our method is more reliable for recording true seed dispersal interactions than direct observation,which only reports birds feeding on fleshy pulp,but not what happens to the seed.Recording the presence of seeds in the birds’ fecal matter guarantees that in the documented interaction,the bird ate the fruit,ingested the seeds and defecated the seed unharmed when captured,something that direct focal observations of frugivory cannot assert(Jordano,2000).Additionally,we would like to remark that,even though riparian forest strips crossing pastures represent a narrow arboreal element subjected to intense edge effects from adjacent pastures and have no forest interior habitat,the species richness of birds and plants that we found is relatively high and similar to that recorded in larger and less disturbed fragments of cloud forest.Even the network metrics of nestedness and modularity detected in our highly disturbed study sites (NODF=11.5;M=0.49) are relatively similar to those reported by Salazar-Rivera et al.(2020) in the peri-urban park of Xalapa(NODF=20.6;M=0.54)and fall within the ranges reported for other Neotropical forests (Vidal et al.,2014;Palacio et al.,2016;Emer et al.,2020).
Our analysis shows that of the 24 bird species recorded,the seven most important in the mutualistic network wereCh.flavopectus(Passerellidae),M.occidentalis,C.mexicanus,T.grayi(Turdidae),E.hirundinacea(Fringillidae),A.brunneinucha(Passerellidae),andMy.similis(Tyrannidae),based on the centrality metrics (D,CC and BC;Table 2) of the network and their contribution to nestedness(Fig.3).These same species are also among the most important seed dispersers of native woody plants in nearby,less disturbed cloud forest fragments(Hernández-Ladrón de Guevara et al.,2012;Baltazar,2014;Pérez-Cadavid et al.,2018).At least three other species recorded in our study (Melanotis caerulescens,Myiodynastes luteiventris,andThraupis abbas) have been reported as important seed dispersers in the studies mentioned above.Several other bird species recorded in our network have also been reported to be effective seed dispersers in the riparian strips of tropical rain forest that cross pastures in the lowlands of southern Veracruz,including:Momotus coeruliceps,My.similis,T.grayi,T.assimilis,D.carolinensis,E.hirundinacea,Th.abbasandSaltator atriceps(Graham et al.,2002).Therefore,our results together with those of similar studies carried out in deforested areas confirm that several species of birds have a high capacity for persisting in anthropic landscapes dominated by agricultural activities,while continuing their role as seed dispersers.All they need is access to arboreal elements on the landscape that provide food resources,perching sites and protection,and riparian cloud forest strips provide all of these.
We recorded four North American migratory birds (Vireo gilvus,V.solitarius,D.carolinensisandParkesia motacilla)as part of the network,revealing the high relevance of these arboreal elements to both resident and migratory bird species(Skagen et al.,1998),and the preservation of this interaction in anthropic landscapes.Other studies have recorded the same migratory species as participants in frugivory networks in Neotropical Forest (Ramos-Robles et al.,2018;Salazar-Rivera et al.,2020).It is noteworthy that in our study even species reported in the literature as preferentially insectivorous (e.g.,P.motacilla,Henicorhina leucophrysandBasileuterus belli)or omnivorous(e.g.,Quiscalus mexicanusandPsilorhinus morio;González-Salazar et al.,2014)were carrying intact seeds in their guts while moving along the riparian strips.Thus,facultative frugivores can also be effective seed dispersers(see also Pizo et al.,2021) that contribute to cloud forest regeneration in fragmented landscapes and could become redundant species in the network(see below).
Of the 30 plant species recorded in our study whose seeds were dispersed by birds,eight were highlighted as the most important in the mutualistic interaction,based on their contribution to the centrality metrics(D,CC,and BC)and network nestedness(Table 2;Fig.3).These eight species wereRhamnus longistyla(Rhamnaceae),M.glaberrima,C.xalapensis,C.arborea(Melastomataceae),Trema micrantha(Cannabaceae),Rubus ulmifolius(Rosaceae),andSolanum aphyodendron(Solanaceae),and all of them provide edible fruit for a large number of birds that would be greatly affected if these plants were not part of the arboreal riparian strips that cross the pastures.These plant species have also been reported amongst the most frequently consumed by birds in relatively large,undisturbed cloud forest fragments in the same region(Hernández-Ladrón De Guevara et al.,2012;Baltazar,2014).Species of the generaMiconia,Conostegia,Palicourea,TremaandSolanumstand out,since they commonly produce an abundance of accessible fleshy fruit that are small and usually available for several weeks of the year to birds of all sizes and thus represent a crucial part of their diets(Snow,1981;Gentry,1982;Pérez-Cadavid et al.,2018).The species ofMiconiaandConostegiaare considered keystone species in the community of Neotropical frugivores (Escribano-Avila et al.,2018;Messeder et al.,2020,2021).These zoochorous woody plants are also very important for the regenerationand restoration of cloud forest,since they are pioneer species able to establish in large canopy gaps,forest edges or abandoned old-fields,and can outcompete heliophytic grasses and weeds by shading them during the early phases of secondary succession (Gorchov et al.,1993;Galindo-González et al.,2000;Hernández-Ladrón De Guevara et al.,2012;Muñiz-Castro et al.,2012).The establishment of these pioneer species facilitates the arrival and establishment of other shrub and tree species of both the intermediate and late phases of secondary succession,whose long-term persistence in anthropic landscapes is in jeopardy due to current management practices.In addition,species of the generaHedyosmum,Saurauia,PerrottetiaandXylosmaare regarded as representative of old-growth cloud forest(Ruiz-Jiménez et al.,2012;Williams-Linera et al.,2013;see also Palacio,2014) and their seeds were being successfully dispersed by birds along the riparian strips we studied.
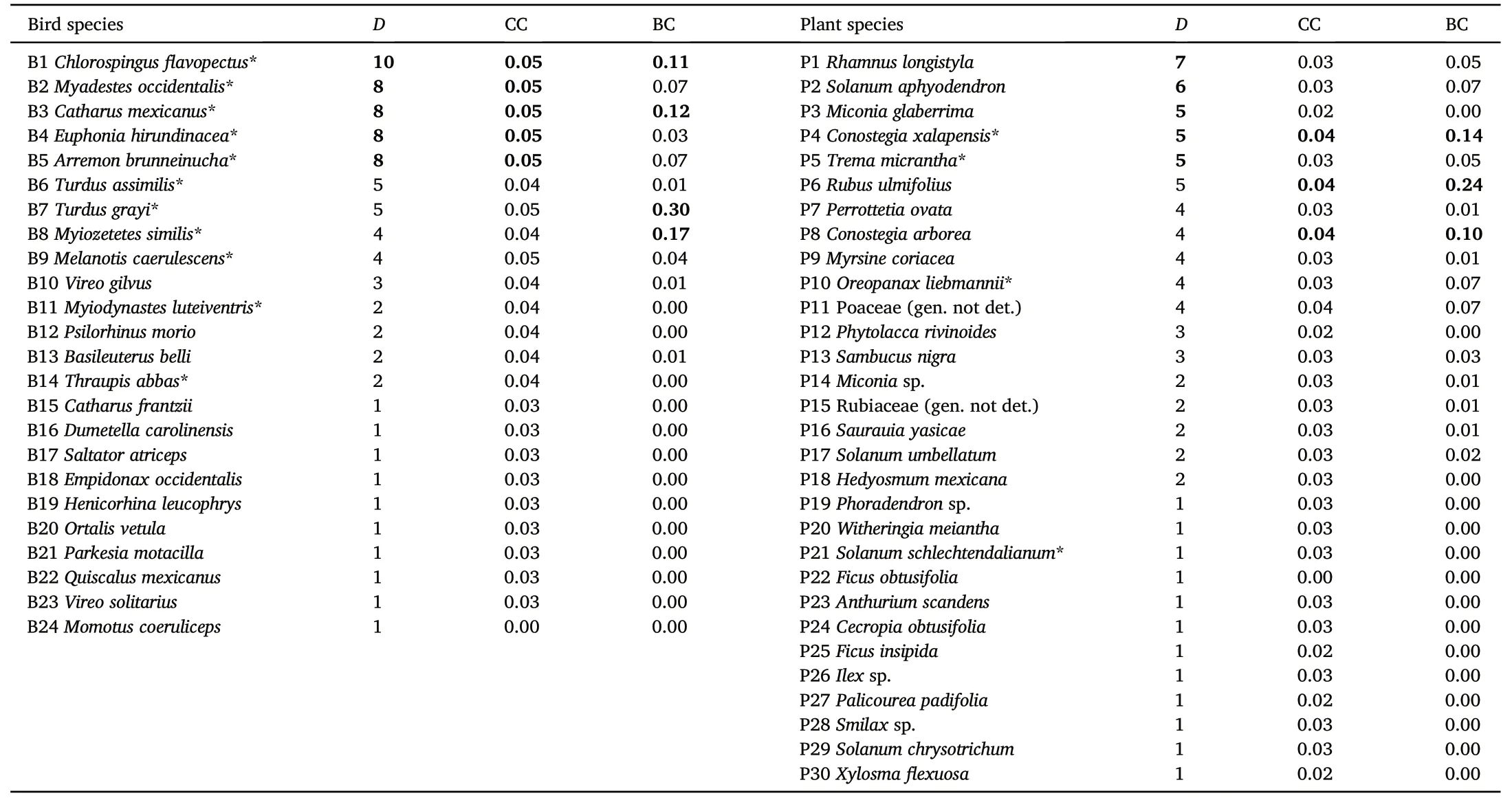
Table 2 Species-level metrics for the bird-plant frugivory interaction network in riparian strips of cloud forest in Veracruz,Mexico.
Some herbaceous and shrubby species that are known to produce large quantities of edible fruit,such asPhytolacca rivinoidesandRubus ulmifolius,or epiphytes such asAnthurium scandensandOreopanax liebmannii,(Snow,1981;Galindo-González et al.,2000;Cruz-Angón and Greenberg,2005;Palacio,2014),were also part of the network we studied,and represent an important resource for birds in Central Veracruz.Even the seeds of two vulnerable species (sensu The Red List of Mexican Cloud Forest Trees;González-Espinosa et al.,2011)-Rhamnus longistylaandO.liebmannii-were dispersed by birds along the riparian strips studied.It is interesting to note the presence of seeds ofCecropiaandFicus(F.obtusifoliaandF.insipida) in our sites.While known to produce copious amounts of fruit readily eaten by birds and bats,they are not common cloud forest trees,but rather are typical of tropical forest at lower elevations(González-Espinosa et al.,2011).Their seeds,therefore,had to have been brought upstream from afar by the birds captured in our study.This clearly indicates that riparian strips are relevant elements of connectivity among forest remnants since they maintain and enhance seed dispersal across the disturbed landscape.
Niche overlap among the species of birds and among the plants in the studied network was low.This is related to the low degree of similarity among bird species with respect to the plants they feed on,so these birds have complementary or contrasting functional roles (not redundant) in terms of seed dispersal(Blüthgen and Klein,2011;Escribano-Avila et al.,2018).Plants also shared few species of bird dispersers,suggesting that the loss of a given species at one trophic level from the landscape could have detrimental effects on the other level.This would increase the risk of subsequent coextinctions even if the network were to lose just a few of its species at first.Deforestation and fragmentation drastically affect seed dispersal and cause a sharp decline in the population size of forest birds and plants that are unable to persist in denuded areas (i.e.,devoid of trees).Our results show that it is imperative to implement pasture management practices aimed at maintaining riparian strips with a high degree of richness and density of native forest species.This would encourage the presence and movement of birds and plants,as well as their interactions in anthropic landscapes.
In the different scenarios of simulated selective extinctions,for both of the interacting groups (birds and plants) the greatest decrease in network stability occurred after deleting the most connected species,with plants the most affected(but see below).The bird species that make the greatest contribution to the network structure (based on the robustness analysis),unfortunately are prone to disappear from the study region because several of them are valued as ornamental species (e.g.,Myadestes occidentalis,Turdus grayi,Euphonia hirundinaceaand some species of the genusCatharus).They are frequently caught by local inhabitants to bring in extra cash (Aguilar-Rodríguez,1992;SEMARNAT,2010;Roldán-Clarà et al.,2014,2017) and their local populations are therefore in great danger.Even though some of these species(C.mexicanus,C.frantziiandM.occidentalis) are protected by Mexican law (Norma Oficial Mexicana-NOM-059),this law is rarely enforced(SEMARNAT,2010).
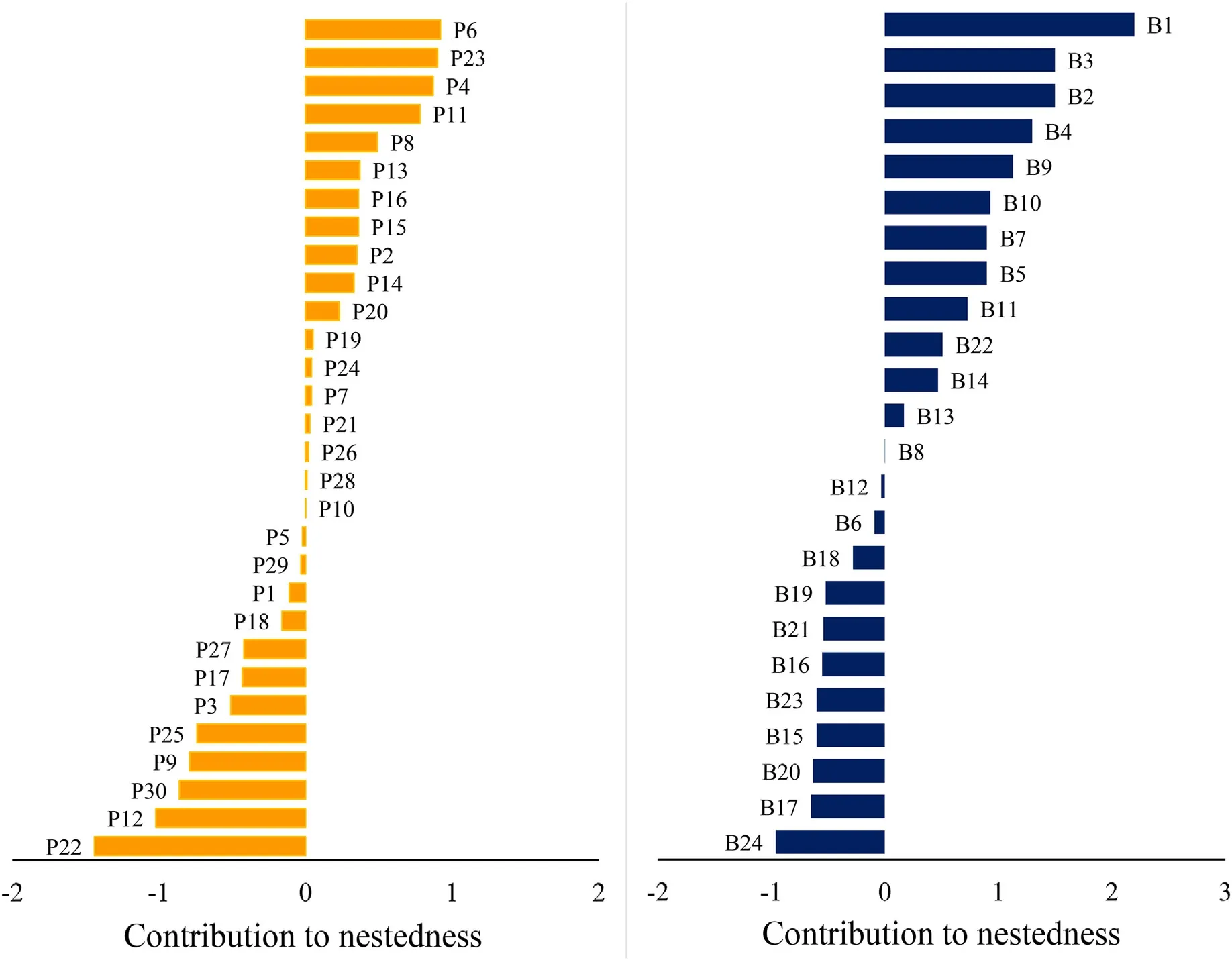
Fig.3.Contribution to nestedness by plants(left panel)and birds(right panel)to the frugivory/dispersal interaction network recorded in cloud forest riparian strips in central Veracruz,Mexico.Species are ordered from highest to lowest contribution to network nestedness in each panel.Species names for each group are in Table 2.
The simulated extinction scenarios should not be accepted at face value since our analysis does not contemplate the possible rewiring of interactions after the removal of a species from the network,nor does it contemplate the role of other species or interactions that were not recorded in our study but may be part of this network.For example,several genera(Piper,Solanum,Conostegia,andFicus)of woody plants in our network are also dispersed by different mammals,particularly bats(Saldaña-Vázquez et al.,2010;Hernández-Montero et al.,2015;Castaño et al.,2018)and non-flying,medium-sized mammals in the cloud forest of Central Veracruz (Gallina et al.,2008) that can disperse their seeds even if bird dispersers disappear from the site.Additionally,several plant species of the generaConostegiaandMiconiaas well as others from the Solanaceae and Rubiaceae families that we know are abundant in riparian strips of our study area and provide fruit for birds (Hernández-Dávila et al.,2020) were not recorded in the droppings of the birds we captured.Therefore,we should not underestimate the role of other taxonomic groups,also present in the region but undetected in our study,which could be important seed dispersers or fruit resources.They could be part of this mutualistic network and make a crucial contribution in terms of redundancy (sensu Memmott et al.,2004).While our completeness curves show that the number of birds and plants recorded is close to the true number of possible interacting species in the network,the completeness curve for the number of interactions (less than 50%)suggests that a greater sampling effort would result in a better representation of the actual interaction network.In spite of this,our analysis allowed us to identify key species in this mutualistic network,some of the main structural characteristics of the network,and the relevance of riparian strips in not only maintaining this network in the current landscape but also in the regeneration of cloud forest.Regardless of the main driver of species loss in a landscape,the importance of riparian strips and of the species that make up the frugivory interaction network in deforested landscapes cannot be overstated.In fact,in the landscape we studied,riparian strips are often the only remaining relicts of forest in an extensive system of pastures and crop-fields.They thus represent key elements for maintaining connectivity between the few,widely spread fragments of well-preserved cloud forest and the numerous patches of secondary forest that are still present in this highly modified landscape.
5.Conclusions
The riparian strips we studied maintain a relatively rich set of birds that disperse the seeds of many species of zoochorous plants,which are notably absent from open areas that are completely devoid of trees.The plant species most important to network structure represent a key source of food for several bird species,in addition to being pioneer species that facilitate the regeneration process of cloud forest in disturbed areas.The robustness analysis shows that network stability may be compromised by the elimination of key species,with the plants most affected by the removal of key bird species.However,the most threatened species in our network are several of the most highly connected bird species,which are illegally captured in the region.In order to ensure the continued existence of these birds,plants and their interactions in intensively deforested and fragmented landscapes,the protection of riparian strips should be a priority.Based on our results,we deem riparian forest strips invaluable arboreal elements of the landscape,not only as biodiversity reservoirs and biological corridors for forest organisms,such as seed dispersing birds,within the agricultural landscape matrix,but also as the custodians of the regeneration potential of the cloud forest.Ideally,longterm conservation and management plans to preserve riparian strips within agricultural landscapes should also include the legal protection of key plant species that provide edible fleshy fruit and key bird species that disperse the seeds of native cloud forest plants,and that made a significant contribution to network stability in our study.
Authors’ contribution
OHD designed the study and wrote the draft manuscript.OHD&VJS carried out field work.All authors ran the statistical tests or interpreted results and contributed to writing and revising the text.All authors read and approved the final manuscript.
Declaration of competing interests
The authors declare that they have no competing interests.
Acknowledgments
We are grateful to María de los Ángeles García and Diana Vázquez for their valuable help in the field.Theejidatariosof the different communities generously allowed us to work on their land.Bianca Delfosse revised the English.Very special thanks to The Rufford Foundation(Our ref: 20471-1),and to the Consejo Nacional de Ciencia y Tecnología(grants CB-2008-01 No.101542,CB-2016-01 No.285962) for funding.Idea Wild donated the equipment that made the field work possible.
Appendix A.Supplementary data
Supplementary data to this article can be found online at https://do i.org/10.1016/j.avrs.2022.100046.
杂志排行

Avian Research的其它文章
- Functional and phylogenetic structures of pheasants in China
- Thermoregulatory function and sexual dimorphism of the throat sack in Helmeted Guineafowl (Numida meleagris) across Africa
- Distribution pattern and driving factors of genetic diversity of passerine birds in the Mountains of Southwest China
- Multiple lines of evidence confirm that the critically endangered Blue-crowned Laughingthrush(Garrulax courtoisi)is an independent species
- Corrigendum to “Multiple lines of evidence confirm that the critically endangered Blue-crowned Laughingthrush (Garrulax courtoisi) is an independent species” [Avian Res.13 (2022) 100022]
- Altitudinal seasonality as a potential driver of morphological diversification in rear-edge bird populations