Shifts in phenology of autumn migration and wing length among reedbed passerines along the East Asian-Australasian Flyway
2022-10-03JohnAllcockTimothyBonerkeYikHeiSungCrolineDingle
John A.Allcock ,Timothy C.Bonerke ,Yik Hei Sung ,Croline Dingle,*
a School of Biological Sciences,Kadoorie Biological Sciences Building,The University of Hong Kong,Pok Fu Lam,Hong Kong SAR,China
b Science Unit,Lingnan University,Hong Kong SAR,China
Keywords:Autumn migration Climate change East Asian-Australasian flyway Passerines Phenology Reedbed
ABSTRACT Climate change impacts bird migration phenology,causing changes in departure and arrival dates,leading to potential mismatches between migration and other key seasonal constraints.While the impacts of climate change on arrival at breeding grounds have been relatively well documented,little is known about the impacts of climate change on post-breeding migration,especially at stopover sites.Here we use long-term (11 years) banding data(11,118 captures) from 7 species at Mai Po Marshes Nature Reserve in Hong Kong,a key stopover site for migratory birds along the East Asian-Australasian Flyway,to describe long-term changes in migration phenology and to compare observed changes to annual weather variation.We also examine changes in wing length over a longer time period(1985-2020)as wing length often correlates positively with migration distance.We found that observed changes in migratory phenology vary by species;three species had later estimated arrival (by 1.8 days per year),peak(by 2.6 days per year)or departure(by 2.5 days per year),one showed an earlier peak date(by 1.8 days per year)and two showed longer duration of passage (2.7 days longer and 3.2 days longer per year).Three species exhibited no long-term change in migration phenology.For two of the four species with shifting phenology,temperature was an important predictor of changing peak date,departure dates and duration of passage.Wing length was shorter in three species and longer in two species,but these changes did not correlate with observed phenological changes.The complex changes observed here are indicative of the challenges concerning the detection of climate change in migratory stopover sites.Continued monitoring and a better understanding of the dynamics of all sites in the migratory pathway will aid conservation of these species under global change.
1.Introduction
Timing of migration exerts a strong selection pressure on migratory birds as individual reproductive output and survival probability are influenced by decisions about the time of arrival and departure at breeding grounds,wintering grounds and stopover sites (Alerstam,2011).Climate change has disrupted the optimal migration strategy for many species,as evidenced by changes in the timing of pre-breeding(spring) migration in birds (McDermott and DeGroote,2017;Horton et al.,2020),with many species arriving at their summer breeding grounds earlier in the year in response to increasing spring temperatures in recent decades(e.g.,Lehikoinen et al.,2004;van Buskirk et al.,2009;Covino et al.,2020).The influence of global climate change on post-breeding (autumn) migration has been relatively less well-studied(Gallinat et al.,2015),although there has been an increase in studies in recent years(Harris et al.,2013;Ellwood et al.,2015;McKinnon et al.,2016).
The timing of spring migration is primarily constrained by the need to arrive on breeding grounds early to establish an optimal territory but without arriving so early that food is unavailable in a highly seasonal environment (Newton,2008).In contrast,multiple ecological and life-history traits may place constraints on the timing of autumn migration,such that the selection pressure to adjust migration timing to weather conditions can vary according to species,sex and age(Newton,2011;Wobker et al.,2021).Factors constraining migration timing potentially include the timing and duration of breeding (Sokolov et al.,1999;Chmura et al.,2020;Imlay et al.,2021),ability to raise multiple broods(Jenni and Kery,2003),timing of moult(Stutchbury et al.,2011;Wobker et al.,2021),body size and morphology (La Sorte et al.,2013;Heim et al.,2018),dietary guild (Jenni and Kery,2003;La Sorte et al.,2013;Heim et al.,2018),migration distance (Jenni and Kery,2003;La Sorte et al.,2015;Barton and Sandercock,2017),latitude of breeding and wintering sites (Bozó et al.,2021),conditions at stopover sites(Hasselquist et al.,2017;Bozó et al.,2018),and niche competition among related species(Heim et al.,2018).
The multiple ecological and life-history constraints on autumn migration may explain the variability observed in long-term trends in the timing of autumn migration.According to the species and location,birds have been found to depart breeding grounds earlier or later(Bitterlin and van Buskirk,2014;Dorzhieva et al.,2020),extend the duration of migration (Zimova et al.,2021),and arrive at wintering sites earlier(Horton et al.,2020)or later(Harris et al.,2013;Brisson-Curadeau et al.,2020).Leaving breeding sites early may be beneficial if it allows individuals to establish high-quality territories on their wintering grounds(Smallwood,1988;Wernham et al.,2002) or if migration can be completed while weather conditions are still relatively stable (Newton,2008).However,species staying longer on the breeding grounds may be able to take advantage of rich food supplies under a longer growing season and possibly raising a second brood,increasing breeding success(Newton,2008;Imlay et al.,2021).In a review of published studies of autumn migration,Bitterlin and van Buskirk (2014) found trends towards later departures in larger species,species that moult prior to migration,and species with granivorous,insectivorous or frugivorous diets,but earlier departures in smaller species and species with a carnivorous diet.Age may also influence phenological responses to climate change,especially in long-lived species (Martín et al.,2016).Perhaps due to the multiple ways in which autumn migration may be impacted by climate change,or because the fitness and reproductive consequences for arriving late at the wintering grounds are less severe,population-level changes to autumn migration phenology are often less pronounced than observed on migration to breeding grounds (Covino et al.,2020;Horton et al.,2020).
Changes in climate may also have an impact on migration distances/routes or choice of wintering sites (Newton,2008;Visser et al.,2009;Rotics et al.,2017).As northern sites become warmer,birds may benefit from overwintering at sites closer to the breeding grounds as this reduces energy expenditure during both migration seasons and allows an earlier return to breeding sites the following year (Newton,2008;O˙zarowska et al.,2018).Changes in the wintering range in response to changing climate have been reported for species in Europe and North America(Bearhop et al.,2005;La Sorte and Thompson,2007;Maclean et al.,2008;Ambrosini et al.,2011).
One of the consequences of a change in migration distance in response to climate change may be a change in wing length or structure.Longer,more pointed wings are considered to be more efficient for power generation during flight than shorter,rounder wings(Wang and Moore,1994) such that species that migrate shorter distances are generally shorter-winged than those with longer migrations (Nowakowski et al.,2014).Shorter wings could also be correlated with an overall decrease in body size,which might be predicted as a response to a warmer climate in accordance with Bergmann's rule(Bergmann,1847).Long-term declines in body size or wing length attributed to warming climate have previously been reported around the world (Yom-Tov,2001;Yom-Tov et al.,2006;Gardner et al.,2009;Salewski et al.,2010;van Buskirk et al.,2010;Jirinec et al.,2021).
Much of the published research on the impact of climate change on migration phenology has been conducted in Europe and North America(Bitterlin and van Buskirk,2014).The East Asian-Australasian Flyway(EAAF) is estimated to support a higher abundance and diversity of migratory birds than any of the other major flyways around the world(Yong et al.,2015),including many species considered threatened by IUCN(BirdLife International,2021).Migrant birds in this region are not faced with barriers as significant as the Sahara Desert,which is known to influence migration patterns among European birds (Jenni and Kery,2003;Newton,2008),and may therefore not be subject to such strong selective pressures on migration.However,the migration of landbirds along the EAAF is relatively understudied compared to other parts of the world,and while there has been an increase in studies in recent years to increase the understanding of migration in Asia(e.g.,Harris et al.,2013;Yamaura et al.,2016;Heim et al.,2018;Bozó et al.,2020,2021;Dorzhieva et al.,2020;Wobker et al.,2021;Yong et al.,2021),still little remains known about the impact of climate change on the timing of migratory events along this flyway,especially in tropical and subtropical latitudes.Further studies are required to confirm whether the trends in migration phenology observed elsewhere also apply to the Asian flyways.
One of the benefits in Europe and North America for conducting this research is the availability of long-term datasets from bird observatories and other migration watch points in these regions,often spanning periods of several decades (e.g.,Sokolov et al.,1999;Hubálek,2003;Hüppop and Hüppop,2003;Jenni and Kery,2003;van Buskirk et al.,2010;Haest et al.,2019;Møller et al.,2021).Such data allows studies to examine long-term changes in observed migration dates which can be compared to trends in climate data (Barton and Sandercock,2017).Compared to Europe and North America,there are few sites in Asia where data has been collected systematically for such a long time period.One such site is Mai Po Marshes Nature Reserve in Hong Kong,an important stopover and wintering site for migratory birds.In this study,we used banding data from the reserve,one of the longest-running banding efforts in Asia,to 1) determine whether autumn migration phenology has changed over the last decade,and 2) assess whether changes in timing and duration of stopovers are correlated with temperature along the flyway or with changes in wing length.
2.Methods
2.1.Trapping
Data used in this study were collected during regular bird banding sessions by the Hong Kong Ringing Group in reedbeds at the Mai Po Marshes Nature Reserve (22.49°N,114.04°E),located along the northern boundary of Hong Kong.The reserve is located at the core of the Mai Po Inner Deep Bay Ramsar Site,an internationally important wetland providing important passage and wintering habitat for migrants along the EAAF.Reedbeds at the reserve are used as a stopover site by migratory songbirds,especially during southward migration,and support populations of globally threatened species such as Manchurian Reed Warbler (Acrocephalus tangorum),Pleske's Grasshopper Warbler (Helopsaltes pleskei) and Yellow-breasted Bunting (Emberiza aureola) (Allcock et al.,2018).This study includes data collected between 2008 and 2018,when trapping effort was consistent on each visit.During the autumn migration season(September-December)over this 11-year period,birds are trapped once or twice a week,for a minimum of 3 h a day.
Birds were trapped in two 96 m lines of mist nets in reedbed habitat,consisting of 8 nets of 12 m each.Nets were opened 30 min before sunrise and trapping continued for 3 h,during which time,the nets were checked regularly.All birds were ringed before being released at the same location.We measured wing length of all newly captured birds.Wing length was measured as the distance from the carpal joint to the tip of the longest primary using a maximum chord method (Svensson,1992).Trapping was conducted under permits issued by the Agriculture,Fisheries and Conservation Department of Hong Kong SAR government.
In this study,we included data on the seven most abundant longdistance migrant species at the site: Black-browed Reed Warbler (Acrocephalus bistrigiceps),Oriental Reed Warbler (A.orientalis),Pallas's Grasshopper Warbler (Helopsaltes certhiola),Lanceolated Warbler(Locustella lanceolata),Dusky Warbler (Phylloscopus fuscatus),Siberian Rubythroat (Calliope calliope),and Amur Stonechat (Saxicola stejnegeri).Most individuals occur at our study site on passage in autumn before continuing south,but all species have also been recorded in the overwintering period.
2.2.Estimation of parameters relating to migration timing
Migration studies often use the dates of the first or last record of a species to define the beginning and end of the migration period and/or the median date of occurrence as a measure of the peak migration period.These measures are most useful when data are collected every day during the migration season,when researchers can be confident that the first,last and median dates were sampled.As our data were not collected daily,these measures of migration timing are potentially influenced by the dates of data collection.Furthermore,two of the studied species(Pallas's Grasshopper Warbler and Oriental Reed Warbler) sometimes start migration through Hong Kong in late August,before the start of our trapping season,and the first arrival date for these species may therefore be missed in our data.
To address this problem,we estimated the migration phenology for each year using parametric season distribution curves to estimate the number of individuals present on each day throughout the season(Knudsen et al.,2007).Gaussian functions follow an assumption that migration date is a normally distributed trait (Linden et al.,2016) and have been found to be an effective way to model the estimated number of birds throughout the season,even accounting for missing data and truncated monitoring seasons,and often outperform General Additive Models(Linden et al.,2016).In order to facilitate comparisons between species and years,we used a single Gaussian function for every species/year combination,with each of the estimated parameters describing the shape of the curve varying each year.Although more complex models(especially a combination of two Gaussian functions)may better describe the occurrence of birds in some years,the use of a different model each year would not allow us to compare between years.
While symmetrical Gaussian functions have been found to fit data well at many observatory sites (Jenni and Kery,2003),we found that right-skewed curves fit better for most species in our study,probably because a high proportion of individuals of most species stop at the site for a period of several days or overwinter at the site.Our models therefore incorporated an estimate for the degree of skewness;a low estimate for skewness suggests a more symmetrical curve(a symmetrical curve has a skewness of 0) while higher estimates of skewness suggest a rapid arrival period followed by a long‘tail’of birds on stopover.
From the number of birds caught on each trapping morning,we created models to estimate the number of birds that would be expected to be trapped on each morning throughout the season,with a separate model generated for each year of the study period.Recaptures of birds that had been caught on a previous date were included in the data set to reflect the total number of individuals present on each day,but multiple recaptures of an individual on the same date were not included.We modelled optimal Gaussian functions to describe the migration phenology for each species -year combination in our dataset using Markhov Chain Monte Carlo (MCMC) techniques with the package R2WinBUGS (Sturtz et al.,2005).The models estimated the mean,variance,skewness and height of a Gaussian curve describing the number of individuals trapped on each of the capture dates over the course of the autumn.We used initial values for these parameters that varied according to species and ran 6000 iterations of the MCMC model to obtain stability in the estimated parameter values.Models covered the period from 1st August to 31st January,which includes periods before and after the peak passage period for all species,to account for both early arrivals at the site before the start of the trapping period on 1st September and the presence of overwintering birds remaining at the site into January.Birds trapped later into the winter and in spring were not used to estimate the Gaussian curve to exclude the possibility of birds on northward migration influencing the shape of the curve.
From the optimized models,we extracted the mean,variance and skewness from the final 2000 iterations of the MCMC model and used these values to estimate the number of birds expected to be present on each day over the migration season.From this,we calculated the expected date of peak occurrence (the highest number of individuals predicted on a single date) and the dates on which the number of birddays exceeded 5%,25%,75% and 95% of the total number of birddays for the entire season (the 0.05,0.25,0.75 and 0.95 quantiles).We used quantiles because these are less sensitive to changes in total population size than the first or last dates of occurrence(Miller-Rushing et al.,2008;Koleček et al.,2020) and do not assume parallel changes throughout the migration season if earlier and later arriving birds respond differently to changes in climate (Dorian et al.,2020).We also estimated the duration of passage as the number of days between the 0.05 and 0.95 quantiles (total passage duration) and between the 0.25 and 0.75 quantiles (peak passage duration).We calculated these dates separately for each of the 2000 iterations and used the mean value as our estimate for the expected dates for each species-year combination.Data from 2008 for Lanceolated Warbler,Siberian Rubythroat,and Amur Stonechat were excluded from statistical analyses because the low number of individuals trapped in that year would limit the accuracy of the estimated phenology curves.Data for Pallas's Grasshopper Warbler from 2008 and 2013 were also excluded from statistical analyses because the estimated dates of peak occurrence in these two years were in late August,prior to the first trapping date,and therefore the estimated shape of the curve was unreliable.
2.3.Weather data
We included both local and regional weather variables in our analysis.Local weather variation is likely to impact the stopover duration at a site,while regional weather may reflect cues birds use to initiate migration.We collected data for temperature,rainfall and wind speed for Hong Kong from the Hong Kong Observatory website (http://www.hko.gov.hk/en/cis/climat.htm).For the analysis,we calculated the daily anomaly from long-term average data(for the period 1981-2010)by subtracting the weather parameter for that day from the reported long-term average for the same date.We then calculated the mean of these anomalies for the period from September to December in each year,to provide an overall annual anomaly for each of the weather parameters(whether the autumn was warmer,wetter and/or windier than average).
Migration has also been linked to large-scale climatic variation,including the North Atlantic Oscillation and El Niño-Southern Oscillation(Forchhammer et al.,2002;Hubálek,2003;Paxton et al.,2014).To investigate whether these larger-scale climatic variations influence migration through Hong Kong we collected temperature anomalies for the Niño 3.4 region of the equatorial Pacific during the years of the study.Temperatures in this region are known to influence migratory patterns of some species in China(Paxton et al.,2014).Data were downloaded from the NOAA website (https://www.cpc.ncep.noaa.gov/data/indices/ssto i.indices).El Niño is considered to occur when the anomaly in this region is more than 0.5 above normal for a three-month period,while La Niña occurs when temperatures are 0.5 below normal for a three-month period.We calculated mean temperature anomalies for this region during two periods in each year-the period from February to August(referred to hereafter as ‘summer ENSO’;this may affect northward migration,breeding phenology or reproductive success prior to migration) and the two-month period of peak migration for each species (referred to hereafter as ‘autumn ENSO’,covering the period September-October for Oriental Reed Warbler,Pallas's Grasshopper Warbler and Amur Stonechat,and October-November for Black-browed Reed Warbler,Lanceolated Warbler,Dusky Warbler and Siberian Rubythroat).
2.4.Statistical analyses
To investigate the long-term changes to migration phenology,we conducted generalized linear model (GLM) regressions of each of the phenology parameters (0.05 quantile,0.25 quantile,peak date,0.75 quantile,0.95 quantile,total passage duration and peak passage duration)using the year as the predictor variable.Because the day number of the phenology parameter is a discrete variable,these GLM models used a Poisson distribution.
For those combinations of species and weather parameters that showed long-term trends,we investigated which weather parameters might account for the observed trend.We studied this using GLM Poisson regression models for each of the phenology parameters,with the predictor variables of temperature,rainfall,wind,autumn ENSO and summer ENSO;we also retained year number as a predictor variable in these models to investigate any long-term trend independent of local weather conditions and to assess whether these explained variation better than the weather parameters.To establish which (if any) of these weather parameters were useful for predicting the phenology metrics,we refined the GLM model using a dredge function in the package MuMIn(Burnham and Anderson,2002) to select the combination of weather parameters providing the lowest AICc.
2.5.Wing length data
To investigate long-term variation in wing length we used data collected from birds ringed in Hong Kong since 1985.This predates the establishment of the trapping site in the reedbeds at Mai Po Marshes Nature Reserve,although this includes data collected from other locations within the reserve.To prevent pseudo-replication,we only used data from the first capture of an individual and excluded data from recaptured individuals.We used generalized additive models to determine variation in body size across years to investigate both linear and non-linear temporal changes.We built models separately for each study species for wing length,body weight,and body condition as the response variable and year as the predictor variable.We estimated the body condition by calculating the residuals from the linear regression of body weight on wing length.We included age and month as random variables.For species where sexes differ in plumage(Siberian Rubythroat and Amur Stonechat),we also included sex as a random variable.Although Dusky Warblers are sexually dimorphic for wing length(Svensson,1992;Bozó and Heim,2016),we found considerable overlap in the wing lengths made it impossible to confidently assign sex to individuals,and so pooled both sexes for this analysis.We fitted the models using the package‘gamm4’(Wood and Scheipl,2013)in software R(R Core Development Team,2014).
3.Results
3.1.Captures during trapping from 2008 to 2018
We trapped at the site on a total of 290 dates between 2008 and 2018(Appendix Table S1).Of the 7 species included in our study,the most numerous species trapped at the site between 2008 and 2018 was Dusky Warbler,with 3727 captures(including original captures and recaptures)over the 11-year period and up to 552 in a single year.The least numerous overall was Amur Stonechat (266 captures),although Lanceolated Warbler had the lowest number of captures during any particular year assessed,with only three birds trapped in 2008.
3.2.Changes to migration phenology
Most species showed later peak,arrival and departure dates,and longer duration of passage over the 11-year study period(Table 1;Fig.1).However,these results were only significant in 4 of the 7 species,each of which showed a different pattern of change over the study period.The end of the Black-browed Reed Warbler passage(0.95 quantile)was later towards the end of our 11-year study period(later by 2.5 days per year,p=0.02)and the duration of the overall passage period and peak passage period were longer(total passage duration 2.7 days longer per year,p=0.001;peak passage duration 1.2 days longer per year,p=0.02).Oriental Reed Warbler showed a later start to the passage period over the course of the study(0.05 quantile later by 1.8 days per year,p=0.001).Dusky Warbler showed a later peak date over the course of the study(2.6 days later per year,p=0.003).Amur Stonechat,however,showed an earlier peak date (1.8 days earlier per year,p=0.04) but a longer duration of passage(total passage duration 3.2 days longer per year,p=0.008;peak passage duration 1.8 days longer per year,p=0.02).We did not detect any changes in migration phenology among Pallas's Grasshopper Warbler,Lanceolated Warbler,or Siberian Rubythroat(Table 1).
3.3.Influence of weather on migration phenology
We examined the influence of weather on phenology parameters that showed long-term change over the course of the study (Table 2).For Black-browed Reed Warblers,an increase in the average autumn temperature by 1°C,caused the 0.95 quantile to be later by 19.3 days,total duration longer by 20.4 days and peak duration longer by 8.6 days.The changes to the phenology of Amur Stonechat were also best explained by temperature trends;for every 1°C increase in temperature,the peak date occurred 13.8 days earlier,total duration was 29.7 days longer and peak duration 14.6 days longer.The trends seen in Oriental Reed Warbler(later 0.05 quantile) and Dusky Warbler (later peak date) were best explained in models containing only the year and without any of the weather parameters.
3.4.Changes in wing length across years
For five of the seven species included in this study,we found a significant change in wing length between 1985 and 2020(Fig.2;Table 3).For three species (Black-browed Reed Warbler,Dusky Warbler,and Siberian Rubythroat),wing length of birds trapped in Hong Kong became shorter over time,while for two species (Oriental Reed Warbler and Lanceolated Warbler) wing length increased over this time period.The wing length of Black-browed Reed Warbler and Siberian Rubythroat declined by 0.025 and 0.031 mm per year whereas the wing length of Lanceolated Warbler increased by 0.104 mm per year(Table 3).Changes in wing length for Oriental Reed Warbler and Dusky Warbler were not linear and varied during the course of the study period.There was no significant change in wing length for Pallas's Grasshopper Warbler or Amur Stonechat.
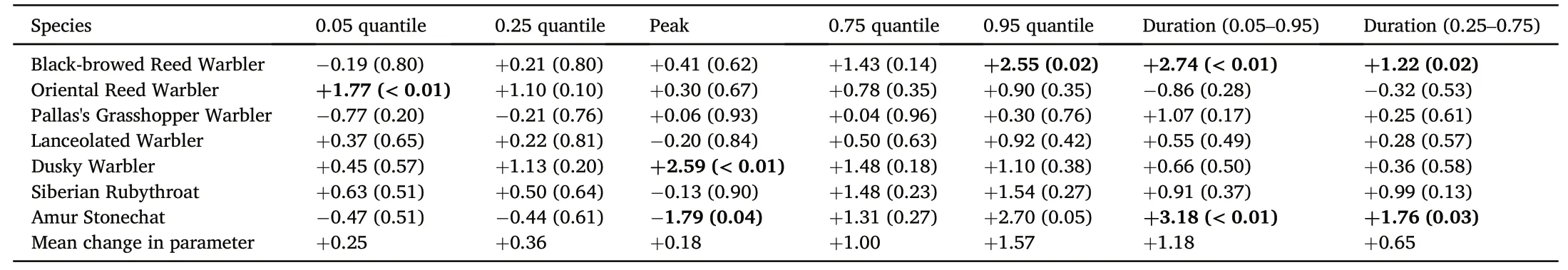
Table 1 Summary of changes in migration phenology for all seven species included in this study.
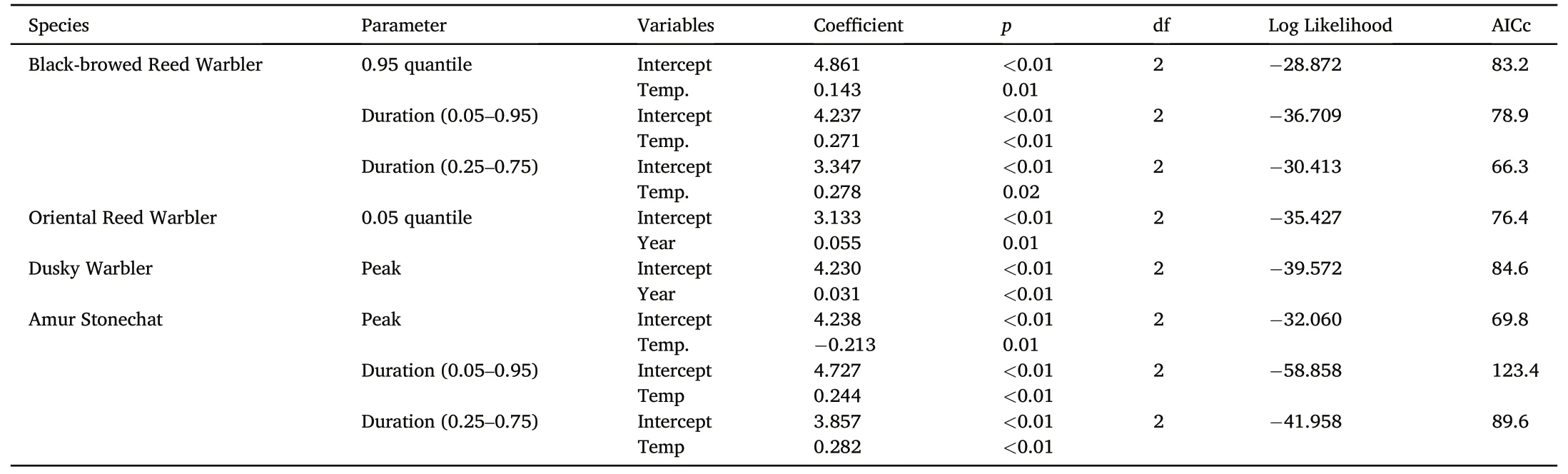
Table 2 Summary of results of GLM dredge of weather parameters to explain observed long-term trends in migration timing parameters.
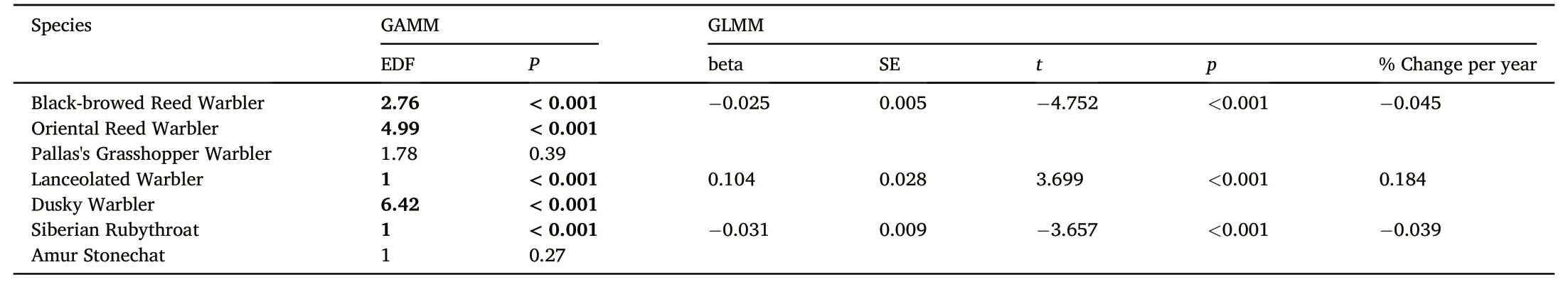
Table 3 Temporal change in the wing length of seven species of songbird from 1985 to 2020 in Hong Kong.
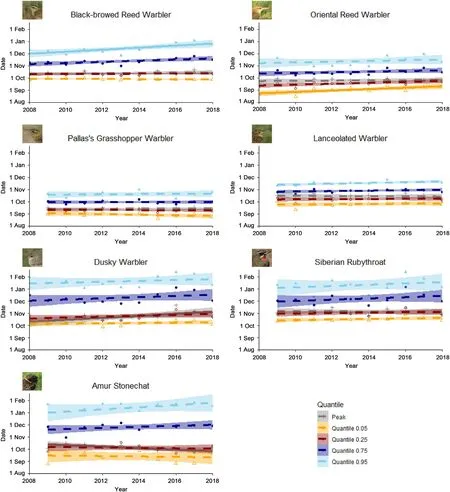
Fig.1.Predicted changes in peak date and dates of 0.05,025,0.75 and 0.95 quantiles between 2008 and 2018.Solid lines represent significant changes(p
4.Discussion
Our results reveal species-specific differences in changes to migration phenology among seven songbird species using reedbeds at Mai Po Nature Reserve as a stopover site during their annual southward migration to their wintering grounds.Only four species showed directional changes in their migration phenology over our 11-year study period:later arrival dates among Oriental Reed Warblers,later end to the migration period and longer duration of passage in Black-browed Reed Warblers,later peak date in Dusky Warblers,and earlier peak and longer duration in Amur Stonechat.The other three species(Pallas's Grasshopper Warbler,Lanceolated Warbler and Siberian Rubythroat) did not show any significant long-term trends in any of the migration parameters studied.While changes are not entirely consistent across species,we did find that the overall trend across all species was towards later migration dates and longer passage periods.We also found that wing length had changed for five out of the seven species,with three showing a shorter wing length and two showing an increase in wing length.Altogether these results support the hypothesis that warming temperatures are impacting morphometrics and migration phenology of landbird species on the East Asian-Australasian Flyway.It is important to note,however,that these findings represent changes observed at a single site,and these changes may be influenced by other factors that have not been included in our study,such as changes to breeding or wintering ranges,shifts in migration routes,and changes to population sizes.More research is needed at other sites along the flyway to investigate whether similar changes are observed elsewhere and to more fully interrogate the causes for the changes recorded here.
4.1.Influence of weather on migration phenology
Previous research has found inter-annual variation in autumn migration timing to be influenced by temperature(Ellwood et al.,2015;Haest et al.,2019;Chmura et al.,2020),precipitation,wind speed anddirection,and barometric pressure (Haest et al.,2019;Chmura et al.,2020).Species vary in their response to these climatic factors,but many species have been found to delay migration in warm and wet conditions(Ellwood et al.,2015;Barton and Sandercock,2017;Chmura et al.,2020),with species-specific differences in responses to these variables.Our study also found that annual changes in migration phenology in response to weather conditions varied according to species.It is important to emphasize that our study only included weather variables in Hong Kong,and variation at other more distant locations along the migration route or on breeding grounds could also impact migration phenology.
Black-browed Reed Warblers and Amur Stonechats had extended passage periods during warmer years in Hong Kong,suggesting that these species may respond to prevailing weather along the migration route by adjusting stopover duration.Under this strategy species may winter further north in warmer years rather than migrating to more southerly locations.Towards the end of the study,we observed Black-browed Reed Warblers regularly wintering at our site where they were previously rare in winter,perhaps in response to the increasing temperatures experienced in later years.This increase does not seem to be accounted for by an increase in overall abundance,as peak counts and the total number of bird-days over the course of the season did not change.This is consistent with observations in Europe and North America,where certain species of birds are recorded wintering in increasing numbers further north as a result of a warming climate (La Sorte and Thompson,2007;Maclean et al.,2008;Curley et al.,2020;Møller et al.,2021) and may alter wintering locations according to prevailing temperature(Ambrosis et al.,2016).Migration strategies may favour birds that migrate shorter distances,simultaneously saving energy and allowing an earlier return to breeding sites in the following season.Changes of the duration of the migratory season,as observed in Black-browed Reed Warblers and Amur Stonechats,are caused by changes to the timing of the start and end of the season occurring at differing rates(Barton and Sandercock,2017;Dorian et al.,2020).Although first and last dates may be affected by changes in abundance,our method using quantiles should be less sensitive to changes in population size,but changes to distributions,migratory routes or seasonal weather may have an influence.Previous research has suggested that these non-parallel changes in phenology affecting migration duration are more likely to affect autumn migration than spring migration(Dorian et al.,2020;Zimova et al.,2021).
The other changes to migration phenology observed in our study were in a delayed start to migration(0.05 quantile)in Oriental Reed Warbler and delayed peak date in Dusky Warbler passing through Hong Kong.Neither of these changes were found to be related to any of the weather parameters studied in Hong Kong.As both are affected by the start of the migration period,it is possible that these are influenced more by weather conditions to the north of Hong Kong,affecting birds before they arrive at the site.For example,warming conditions on the breeding grounds may encourage birds to start migration later and thus arrive at stopover sites later.
Some of the observed changes to migration phenology may result from changes on the breeding grounds or migratory routes not documented in our study.Several bird species are known to have shown northward expansion of breeding ranges in response to warming climate(Thomas and Lennon,1999;Hitch and Leberg,2007;Yong et al.,2021),which could result in increased population sizes and changes to migration routes and stopover sites.Longer migratory journeys from northward expansion of breeding grounds could be a cause of later arrival at stopover sites,as observed with Oriental Reed Warbler and Dusky Warbler.Small changes to migratory routes could have a large impact on the number of birds present at a single stopover site.
Comparing migration phenology to climate variables at the breeding grounds would be informative,but very little is known about the origin of individuals passing through Hong Kong.The seven species included in our study have ranges covering a wide area of Asia.Oriental Reed Warbler,Black-browed Reed Warbler and Amur Stonechat have breeding ranges covering central and northeastern China,eastern Mongolia,southeastern Russia,Korea and Japan,while the other four species also include large areas further north in Russia,with Siberian Rubythroat reach as far northeast as Chukotka,and Lanceolated Warbler as far west as Finland.However,there are relatively few banding studies throughout these breeding ranges and the only confirmed movements between Hong Kong and breeding grounds among these seven species involve Blackbrowed and Oriental Reed Warblers migrating between Hong Kong and Japan(Carey et al.,2001;Allcock et al.,2021).Morphological variation among Pallas's Grasshopper Warbler trapped during the study suggests that birds present locally originate from multiple breeding populations.Further research to investigate the breeding distribution of individuals involved and whether the timing of migration through Hong Kong is affected by weather conditions at breeding grounds would be informative.
4.2.Interaction between changes in migration phenology and wing length
Three of the seven species included in this study showed a decrease in wing length over the period from 1985 to 2020,while two showed an increase in wing length and two showed no change.Temperatures in Hong Kong have increased over this period(increasing at a rate of 0.24°C per decade from 1991 to 2020;https://www.hko.gov.hk/en/clima te_change/obs_hk_temp.htm).Long-term declines in wing length have also been reported in birds from Israel,Australia,and the Eastern US(Yom-Tov,2001;Gardner et al.,2009;van Buskirk et al.,2010),while Amazonian birds are reported to have declining body mass but increasing wing length (Jirinec et al.,2021).Wing length is correlated with migration distance (Nowakowski et al.,2014) and timing of spring migration but not with the timing of autumn migration (Zimova et al.,2021).Among the three species showing a decrease in wing length over the study period,Dusky Warbler and Siberian Rubythroat both overwinter in Hong Kong,while Black-browed Reed Warbler previously occurred mostly as a passage migrant with wintering range to the south but has also been overwintering in increasing numbers.The observed decline in wing length may be an adaptation towards shorter migrations in these species in response to climate change.In the case of Dusky Warbler,males are longer-winged than females (Svensson,1992;Bozó and Heim,2016)and we cannot rule out the possibility that the observed decrease in wing length in this species could be partly due to a change in ratio between males and females locally.The two species showing an increase in wing length (Oriental Reed Warbler and Lanceolated Warbler)occur almost entirely as passage migrants in the study site,with few individuals overwintering locally.
Climate change would also be predicted to select for smaller body size under Bergmann's Rule (Bergmann,1847) and wing length is often positively correlated with body size(Gosler et al.,1998).However,wing length may not necessarily scale in proportion to changes in body size.For example,increasing wing length despite decreasing body size has been reported in North American migratory species(Weeks et al.,2019)and in the Amazon rainforest(Jirinec et al.,2021).Salewski et al.(2014)found that wing length and tarsus length of European species varied independently and did not correlate to changes in climate.Remacha et al.(2020)demonstrated that wing length in Common Nightingale(Luscinia megarhynchos) declined over time despite greater survival of longer-winged birds during migration,perhaps because shorter wings enhance breeding success more than survival.
Wing lengths of Oriental Reed Warbler and Lanceolated Warbler increased over the 35-year period included in our study.Longer wings allow for more cost-efficient flight,so that birds can cover longer distances with given fat stores,thus reducing the need for stopovers during migration(Wang and Moore,1994;Alerstam,2011;Nilsson et al.,2013).Longer wings are often associated with a greater total migration distance(Nowakowski et al.,2014)and may reflect a northward expansion of the breeding range of these species,as has been documented for other species(Thomas and Lennon,1999;Hitch and Leberg,2007;Yong et al.,2021).Longer-winged birds have been found to migrate earlier in spring(Zimova et al.,2021) and complete spring migration faster than shorter-winged conspecifics (Stolt and Fransson,1995;Potti,1998;Cooper et al.,2011;Hahn et al.,2016).This may provide an adaptation to faster or less predictable progression of spring onset dates in some species(Hahn et al.,2016),where commencement of migration from wintering grounds is determined by environmental cues(such as photoperiod)that are unconnected to conditions on the breeding grounds(Gwinner,2003;Newton,2008).However,it has also been suggested that faster migration may make a species more vulnerable to changing climate(Hurlbert and Liang,2012).Thus,the observed increase in wing length for these species may benefit them by allowing faster spring migration or an earlier return to the breeding sites,which would not be detected by our research into autumn migration phenology.
5.Conclusions
Impacts of climate change on the timing of autumn migration are likely to be variable between species,as life history traits will impact optimal times for leaving breeding sites and arrival times at wintering sites.Of seven species of passage migrants which use Mai Po Marshes Nature Reserve as a stopover site during their autumn migration,we found that four have changed some aspect of their migration phenology over the study period,but that the response varied by species.Temperatures in Hong Kong correlate to changes in two of these species.We also found evidence that wing lengths have changed over the past 30 years.These changes could be a direct response to warming temperatures or an indirect response to changing migratory timing or distances over this time period.Despite the relatively small geographical and taxonomic scale of this study,our findings are consistent with changes observed in other parts of the world and provide baseline data on the potential impacts of climate change on migration phenology along the East Asian-Australasian Flyway.We recommend further research to investigate whether similar changes are observed elsewhere along this flyway,especially through the establishment,maintenance,and coordination between long-term banding efforts at sites throughout the region.The use of museum collections along with data from these long-term banding sites could provide additional insights into the impact of climate change on morphological traits linked with changes in migration phenology and pathways.
Ethics statement
Not applicable.
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Acknowledgements
Data for this study were provided by the Hong Kong Ringing Group and the Hong Kong Bird Watching Society.Photographs for Fig.1 were provided by P.J.Leader.Management of the trapping site was carried out by WWF-Hong Kong.We thank Wenda Cheng for advice early in the project.Funding was provided by an RAE Improvement Grant to (TCB)from the Faculty of Science at The University of Hong Kong.
Appendix A.Supplementary data
Supplementary data to this article can be found online at https://do i.org/10.1016/j.avrs.2022.100052.
杂志排行

Avian Research的其它文章
- Functional and phylogenetic structures of pheasants in China
- Thermoregulatory function and sexual dimorphism of the throat sack in Helmeted Guineafowl (Numida meleagris) across Africa
- Distribution pattern and driving factors of genetic diversity of passerine birds in the Mountains of Southwest China
- Multiple lines of evidence confirm that the critically endangered Blue-crowned Laughingthrush(Garrulax courtoisi)is an independent species
- Corrigendum to “Multiple lines of evidence confirm that the critically endangered Blue-crowned Laughingthrush (Garrulax courtoisi) is an independent species” [Avian Res.13 (2022) 100022]
- Altitudinal seasonality as a potential driver of morphological diversification in rear-edge bird populations