欧洲山毛榉不同月份光合特性研究
2022-10-02卢绍辉袁国军龚东风马俊青
卢绍辉,袁国军,龚东风,杨 昆,马俊青
(河南省林业科学研究院,河南 郑州 450008)
欧洲山毛榉(Fagus sylvaticaL.),属于壳斗科(Fagaceae),水青冈属(Fagus)植物,树冠呈塔形、卵形或圆形,叶片单叶互生,呈卵形,全缘或具圆齿及波状,秋季叶片为黄色,叶背淡绿色,且叶片继续保留在枝干上直到来年春天。花单性,雌雄同株,4~5月开花;果壳斗,9~10月成熟;适宜在阴暗潮湿及排水良好的酸性土壤环境中生长,对环境变化敏感,不宜在土壤板结或气候炎热的条件下生存。欧洲山毛榉广泛分布于北美、西欧,是温带阔叶落叶林的主要构成树种之一[1]。在欧洲、北美和新西兰,欧洲山毛榉是一种非常受欢迎的公园和庭院观赏树木,因欧洲山毛榉木材坚硬,纹理密致,适为家具和农具用,亦为优质的铁路枕木[2],其用途非常广泛。
植物对外界环境的响应是生理和代谢活动的特定变化,外界刺激限制了其功能的完整性。植物的光合作用是将太阳能转换为化学能的过程,是植物进行物质转换和能量转换的重要途径,叶片光合作用是受影响最严重的活动之一,因为它需要通过控制气孔CO2吸收量保持与树木水分的平衡[3-4]。树木生长主要受到光合作用的影响,而光合作用是一个非常复杂的树木生理现象,受树木生长环境中很多环境因子(光照、温度、CO2浓度和空气湿度等)的影响[5-6]。植物进行光合作用的能力直接决定了植物的生存和繁殖能力,植物对环境变化的生理响应主要表现为气孔交换特点的变化,通常用净光合速率(Netphotosynthetic rate,Pn)、蒸腾速率(Transpiration rate,Tr)、气孔导度(Stomatal conductance,Gs)和胞间二氧化碳浓度(Intercellualr CO2concentration,Ci)等指标来衡量植物的光合特性差异。光响应曲线(Light response curves,PLC)描述了植被净光合速率(Pn)与光合有效辐射(Photosynthetically active radiation,PAR)的关系,常用于计算最大净光合速率(Maximum netphotosynthetic rate,Pn max)、光饱和点(Light saturation point,LSP)、光补偿点(Light compensation point,LPC)、暗呼吸速率(Dark respiration rate,Rd)、表观量子效率(α)等生理指标,光合指标和光响应曲线能较为直观的反映植物的光和能力[7],对研究植物光合能力、光合潜力及生态适应性具有重要意义[8],这为研究植物生长发育规律和良种选育提供了重要的理论依据[9]。目前,国内外学者对树木光合特性有很多方面研究,例如作用机理、超微结构以及酶等的研究[10-12],在胁迫条件下(干旱、水淹、盐胁迫、低温或耐阴等)的光合特性研究[13-19]。植物的光合速率受到气孔导度和其他光合特性的影响[20-21],与植物的环境适应能力有着直接或者间接的关系[22],研究植物的光合特性,有利于了解植物对光能的利用效率,阐明植物光合的生态学特征[23]。通过测定植物光响应曲线及相应参数(光补偿点和光饱和点)判断该植物对光的适应范围,以此来确定植物适宜生长区域。
欧洲山毛榉具有很高的观赏价值和经济价值,对环境适应能力较强,在国际间的引种日益增多。目前,国内外对欧洲山毛榉[24-26]和南极假山毛榉(Nothofagus antarctica)[27-28]干旱胁迫下生理特性的影响研究较多,但对欧洲山毛榉光合特性相关方面的研究相对较少,发现仅对其形态特征和树种分布有所报道,不同月份的光合特性差异目前尚不清楚。研究欧洲山毛榉光合作用的日变化和月份变化特点,通过分析其变化规律找出影响欧洲山毛榉净光合速率变化的生理生态因子,明确欧洲山毛榉不同时期的光合生产力,以期对欧洲山毛榉栽培技术研究提供理论依据。
1 材料与方法
1.1 试验区概况
试验地点位于河南省济源市思礼镇,经纬度坐标为北纬35°06'34.85″、东经112°30'31.78″,地处河南西北部,太行山南麓,与山西省毗邻,海拔为160 m。属暖热带季风气候,四季分明,气候温和,光、热、水资源丰富,但受季风影响显著,干旱或半干旱季节明显,春季气温回升快,多风少雨、干旱频发;夏季炎热,热量充足,降雨集中,局部易涝易旱;秋季凉爽,气温降幅较大,雨量减少;冬季寒冷,雨雪稀少。全年平均气温14.6℃,全年日照1 727.6 h,全年降水量860 mm。
1.2 试验材料与仪器
试验材料为欧洲山毛榉容器移栽苗,5年生,行距3 m,株距1 m,苗高约0.9 m。试验仪器为Li-6400XT型便携式光合测定系统分析仪(LI-COR公司,美国)。
1.3 测定方法
1.3.1 欧洲山毛榉光合日变化测定
采用Li-6400XT型便携式光合测定系统分析仪,在阳光充足无风的晴天,使用开放气路随机选取受光、叶位一致的叶片进行测定。测量参数包括:净光合速率、气孔导度、蒸腾速率、胞间CO2浓度、光合有效辐射、叶片温度(Tleaf)、气温(Tair)。
试验于2017年6~9月,每月中旬选择1天(晴朗无风,具代表性)进行测定,共测定4天,光合日变化测定时间为6:00~18:00,每隔2 h测定一次。大田内5点取样,每点测定5株,共测定25株,每株选3片中上部功能叶片,每片叶测3次,30 min内完成,取平均值。
1.3.2 欧洲山毛榉光响应曲线测定
利用LED红蓝光源测定梯度光照强度下欧洲山毛榉的光合速率,测定时间为9:00。测定前,测定对象在自然光强下诱导,此时光强为1 200~1 800μmol/m2·s,以充分活化光合系统,使用开放气路,外界空气CO2浓度在400μmol/mol左右,测定6片叶,设定12个光强梯度,分别为2 500、2 000、1 500、1 000、500、200、150、100、80、50、20、0(μmol/m2·s)。测定对象与日变化选定的测定对象相同,自动取值间隔设置为最小等待3 min,最大等待5 min。光补偿点(LCP,μmol/m2·s)、光饱和点(LSP,μmol/m2·s)、最大净光合速率(Pnmax,μmol/m2·s)、表观光量子效率(Apparent quantum yield,AQY,mol/mol)和暗呼吸效率(Rd)等参数根据光合软件拟合曲线计算,将各参数代入非直角双曲线模型(Thornley)[25]中计算得到不同光强下的净光合速率。
1.4 数据分析
使用Excel 2007和SPSS 20.0统计分析软件对数据进行整理分析。
2 结果与分析
2.1 不同月份欧洲山毛榉光合特性日变化
2.1.1 不同月份欧洲山毛榉净光合速率(Pn)日变化
欧洲山毛榉6月份测定的Pn值日变化为双峰曲线,峰值分别出现在8:00和16:00,最高峰出现在8:00,峰值为8.90μmol CO2/m2·s。在14:00有明显的“午休”现象,Pn值为4.34μmol CO2/m2·s。8月份Pn值日变化与6月份相似,也为双峰曲线,两个峰值分别出现在8:00和16:00,在14:00有明显的“午休”现象,其Pn值仅为0.14μmol CO2/m2·s。7月的Pn值日变化为单峰曲线,10:00达到高峰,峰值为3.28μmol CO2/m2·s,然后开始下降,18:00下降到最低值1.11μmol CO2/m2·s,没有“午休”现象。9月Pn的日变化与7月份较为相似,为单峰曲线。10:00达到高峰,峰值为1.69μmol CO2/m2·s(图1)。
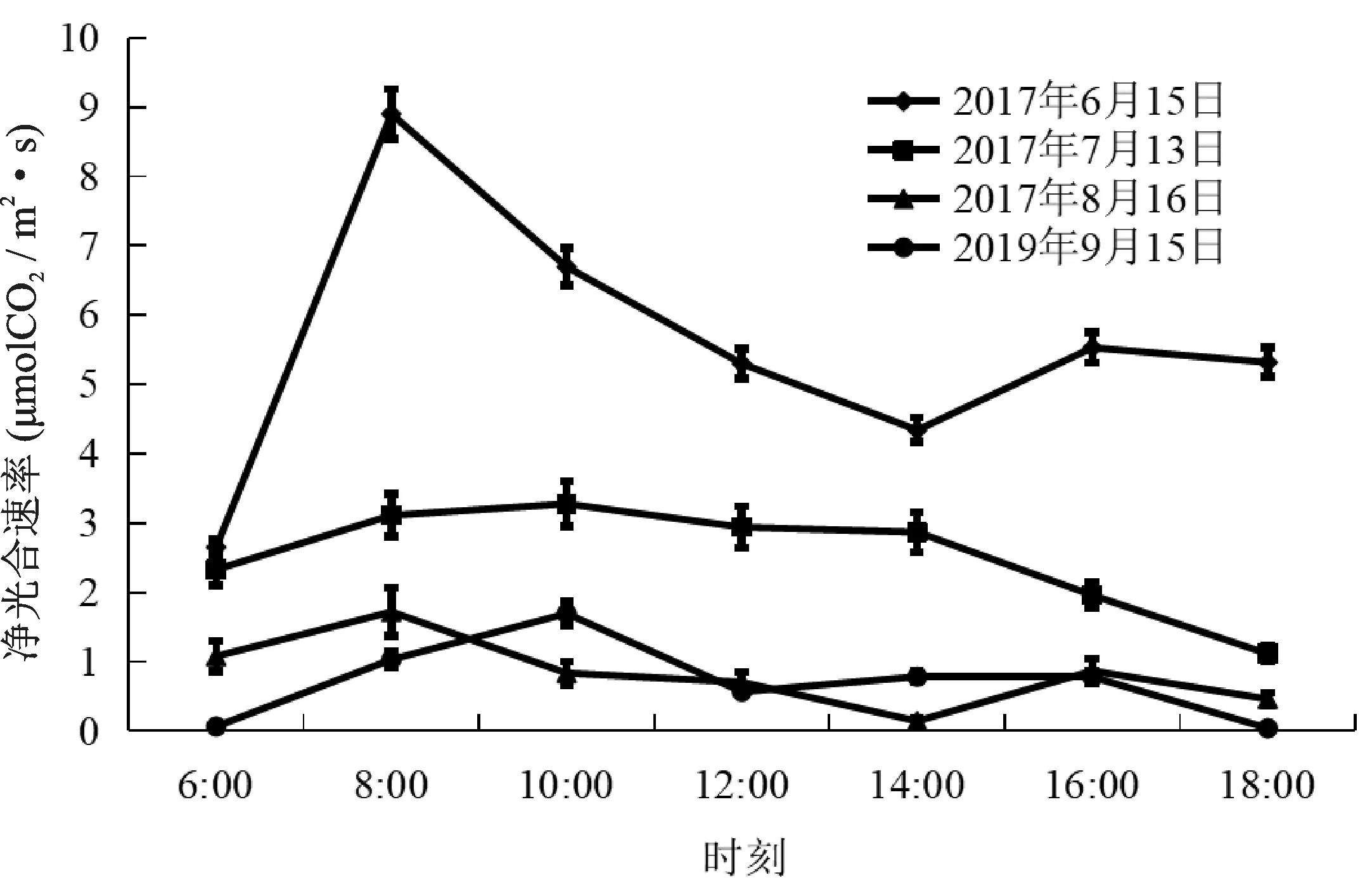
图1 不同月份欧洲山毛榉净光合速率日变化Fig.1 Diurnal variation of Pn in different months of F.sylvatica
2.1.2 不同月份欧洲山毛榉蒸腾速率(Tr)日变化
欧洲山毛榉不同月份的蒸腾速率均呈单峰型曲线。6月上午6:00~8:00 Tr缓慢增大,8:00~10:00 Tr迅速增大,最大值为6.26 mmolH2O/m2·s,然后开始下降,18:00降到最小值3.05 mmolH2O/m2·s。7月上午6:00~10:00 Tr呈缓慢下降趋势,10:00~12:00 Tr略有增加,12:00以后持续下降,到18:00下降到最小值0.91 mmolH2O/m2·s。8月份上午6:00~12:00 Tr缓慢增大,直到12:00出现最大值为2.37 mmolH2O/m2·s,然后开始下降,18:00降到最小值1.26 mmolH2O/m2·s。9月上午6:00~8:00 Tr略有变小,8:00~12:00 Tr缓慢增大,并且达到最大值为1.04 mmolH2O/m2·s,然后平缓下降,18:00降到最小值0.14 mmolH2O/m2·s(图2)。
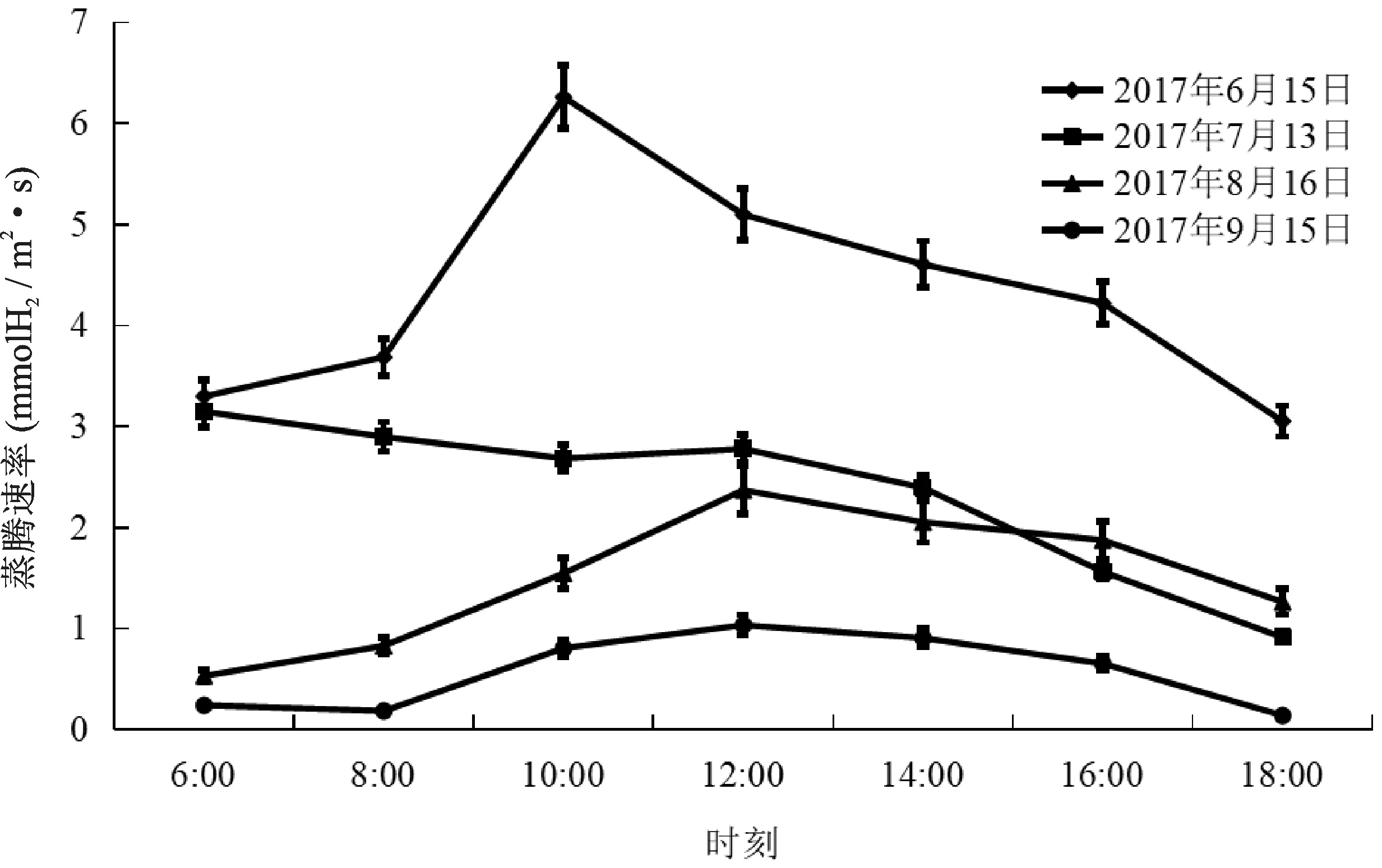
图2 不同月份欧洲山毛榉蒸腾速率日变化Fig.2 Diurnal variation of Tr in different months of F.sylvatica
2.1.3 不同月份欧洲山毛榉胞间CO2浓度(Ci)日变化
6月上午6:00~8:00 Ci呈下降趋势,8:00~16:00 Ci基本稳定在290μmolCO2/mol左右,最高值出现在早上6:00,为399.0μmolCO2/mol。最低值出现在18:00,为274.9μmolCO2/mol。7月份6:00~14:00 Ci呈下降趋势,14:00~18:00 Ci呈上升趋势。最高值出现在6:00,为479.3μmolCO2/mol。最低值出现在14:00,为229.3μmolCO2/mol。8月份6:00~10:00 Ci呈下降趋势,10:00~18:00 Ci在310~350μmol/mol之间波动,最高值出现在早上6:00,为510.9μmolCO2/mol。9月上午6:00~10:00 Ci呈下降趋势,10:00~14:00 Ci在330~400之间变化,16:00~18:00又上升到457.8μmolCO2/mol,最高值出现在早上6:00,为481.7μmolCO2/mol(图3)。
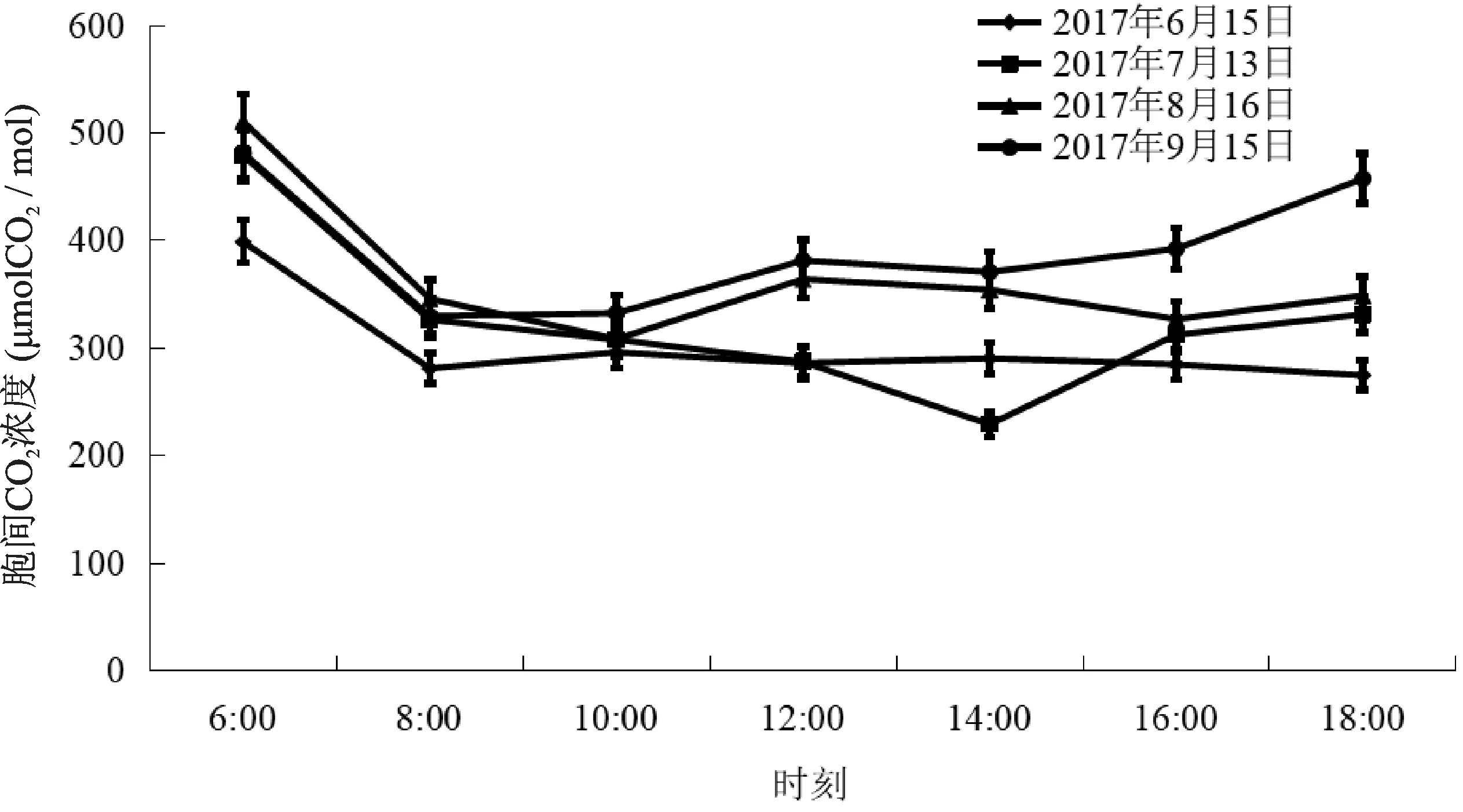
图3 不同月份欧洲山毛榉胞间CO2浓度日变化Fig.3 Diurnal variation of Ci in different months of F.sylvatica
2.1.4 不同月份欧洲山毛榉气孔导度(Gs)日变化
6月上午6:00~8:00 Gs呈上升趋势,8:00~14:00呈下降趋势、14:00~16:00缓慢上升,16:00~18:00又开始下降,1天有2个峰,分别出现在上午8:00和下午14:00,最大值为0.135μmolH2O/m2·s。7月份6:00~12:00 Gs呈上升趋势,12:00~18:00呈下降趋势,全天只有1个峰,出现在中午12:00,最大值为0.099μmolH2O/m2·s。8月上午6:00~10:00 Gs呈上升趋势,上午10:00达到最大值0.061 μmolH2O/m2·s;10:00~18:00 Gs变化不大,基本在0.045~0.055μmolH2O/m2·s之间波动。9月Gs变化趋势与7月一致。上午6:00~12:00 Gs呈上升趋势,12:00~18:00呈下降趋势,全天只有1个峰,出现 在 中 午12:00,最 大 值 为0.043μmolH2O/m2·s(图4)。
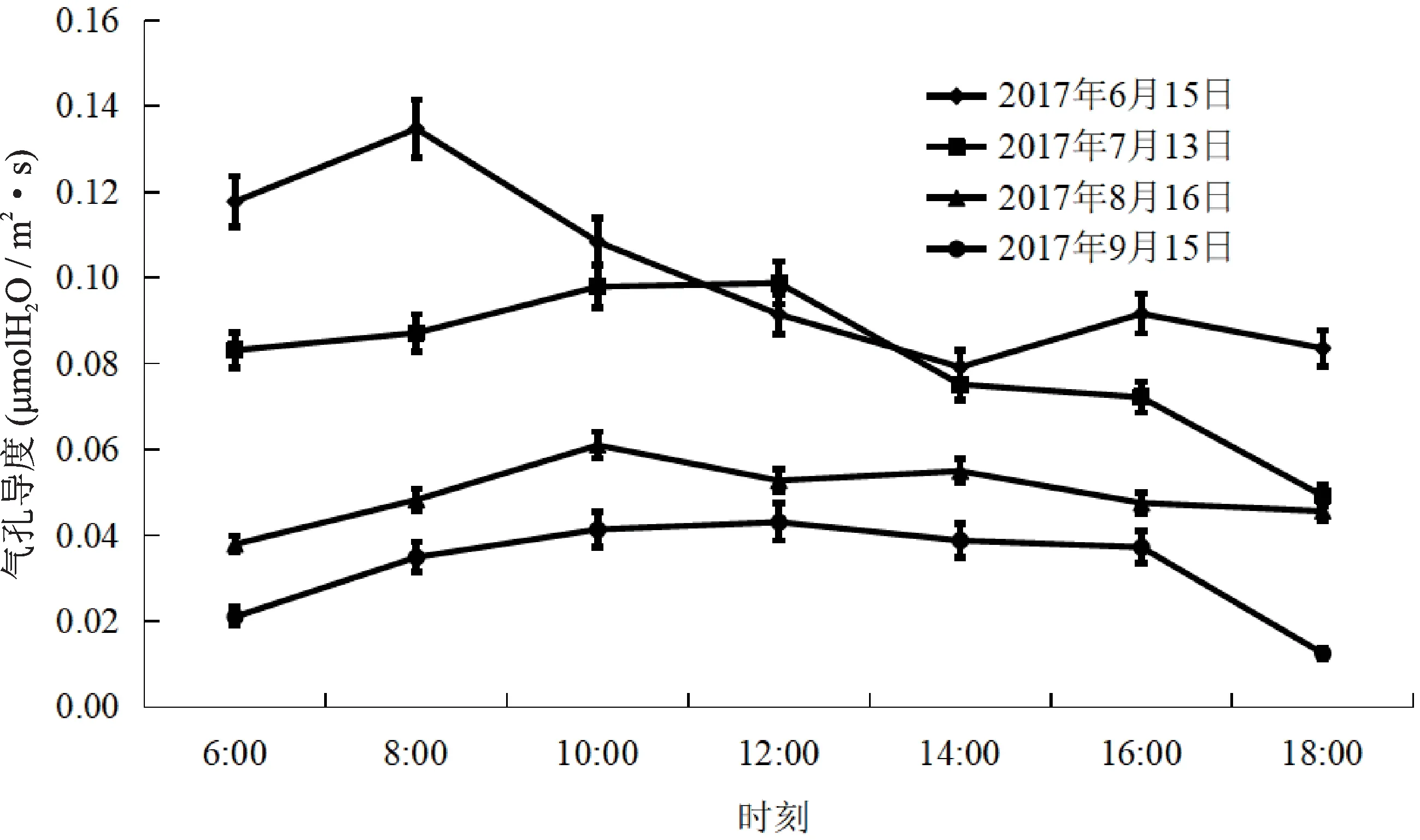
图4 不同月份欧洲山毛榉气孔导度日变化Fig.4 Diurnal variation of Gs in different months of F.sylvatica
2.2 不同月份欧洲山毛榉叶片光合作用特性的差异
测得欧洲山毛榉的净光合速率(Pn)、蒸腾速率(Tr)和气孔导度(Gs)3项光合指标在6月最大,随着月份增加逐渐减小。胞间CO2浓度(Ci)的变化则与之相反,随着月份的增加逐渐增加。6月的Pn值显著高于7月,7月的Pn值显著高于8月和9月,8月和9月的Pn值无显著差异。6月的Tr值显著高于7月和8月,7月和8月的Tr值无显著差异,9月的Tr值显著低于7月和8月。Gs值以6月份最高,9月份最低,4个月份之间均存在显著差异。6月的Ci值显著低于9月份,其余月份之间的Ci值无显著差异(图5)。
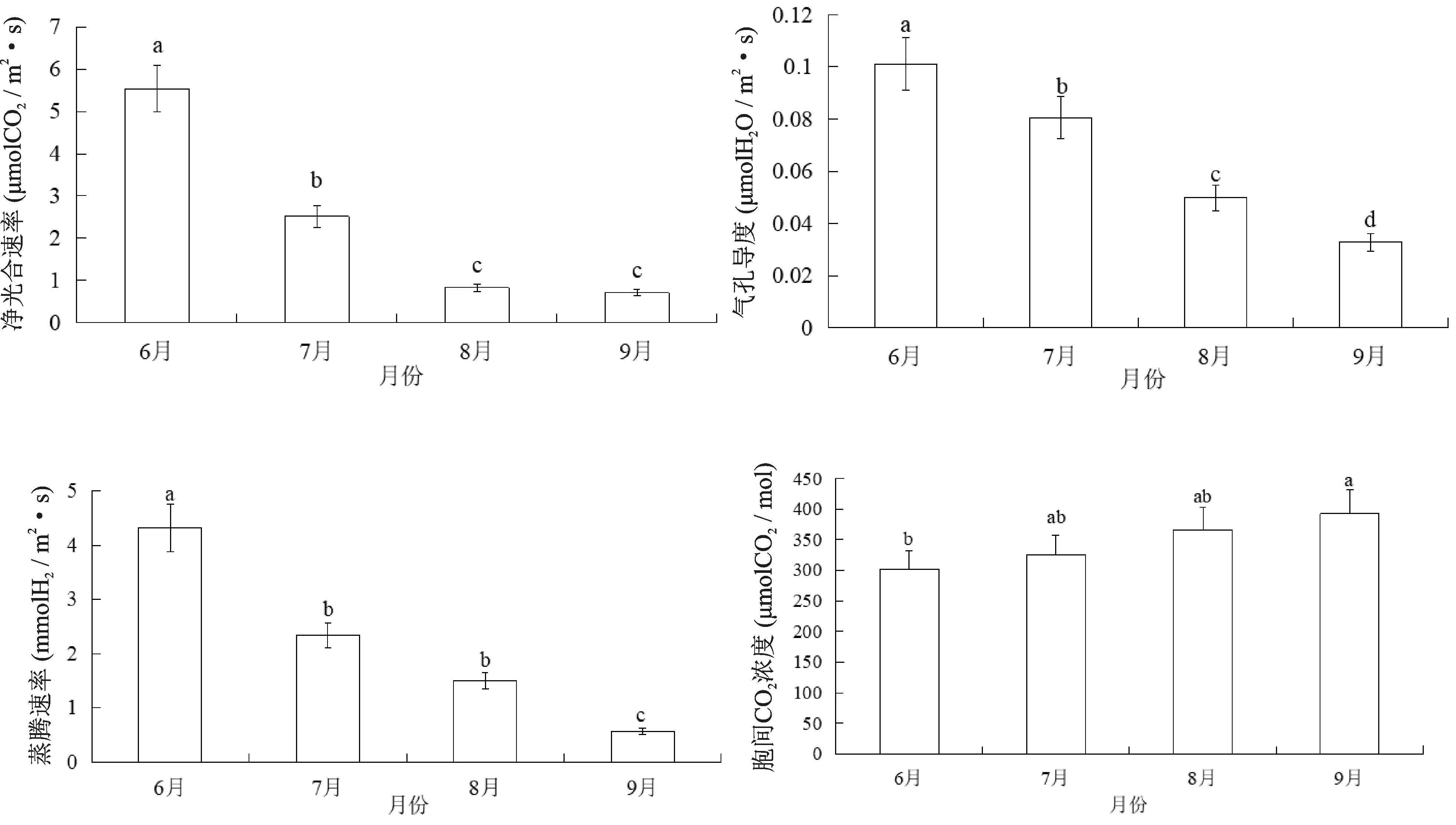
图5 不同月份欧洲山毛榉光合特性比较Fig.5 Comparison of photosynthetic characteristics of F.sylvatica in different months
2.3 Pn与各因子之间的相关性分析
6月份和8月份Pn与内、外各环境因子的相关性均没有显著差异,这可能是欧洲山毛榉在这两个月的光合作用受多种因素共同影响产生的结果;7月份Pn与Gs呈极显著正相关,与Tr呈显著正相关,说明7月份光合作用主要受气孔导度的影响,加快蒸腾速率更有利于欧洲山毛榉生长发育,9月份欧洲山毛榉Pn与胞间浓度(CO2)呈极显著正相关,说明加快欧洲山毛榉呼吸作用更有利于光合作用(表1)。
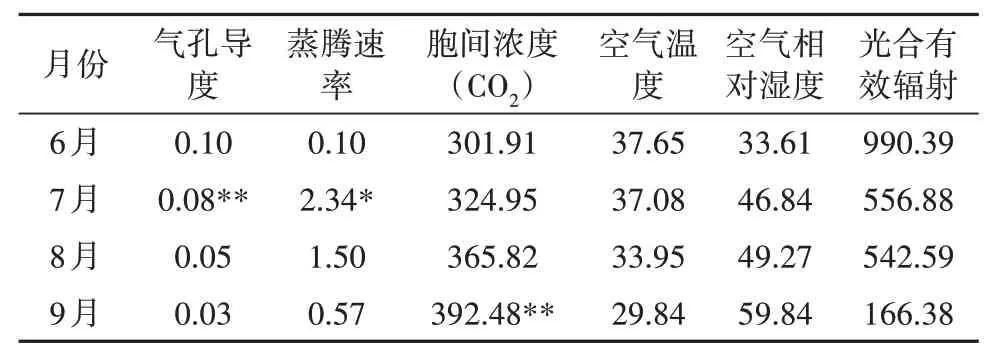
表1 欧洲山毛榉不同月份Pn与各因子之间的相关性分析Tab.1 Correlation analysis of Pn and various factors of F.sylvatica in different months
2.4 欧洲山毛榉对光的响应
以Pn为因变量,PAR为自变量,分析欧洲山毛榉净光合速率(Pn)与光合有效辐射(PAR)相互关系,根据实际测得的试验数据绘制欧洲山毛榉响应曲线。PAR在0~200μmol/m2·s范围内,Pn随着光强的增加,呈线性上升趋势,PAR在200~2 000μmol/m2·s范围内,Pn随着光强的不断增大,上升趋势减缓。光强在0~200μmol/m2·s范围内与净光合速率进行线性回归分析,得线性回归方程为:y=0.024 1x-0.882 4,R2=0.982 2。由此可知,欧洲山毛榉的表观量子效率为0.024 1,暗呼吸速率为-0.882 4μmol/m2·s,光补偿点为36.614μmol/m2·s。经光合作用与光强在0~2 500μmol/m2·s范围内进行二次多项式曲线回归,得回归方程为:y=-2×10-6x2+0.007 7x+0.041 47,R2=0.870 6,计算得欧洲山毛榉光饱和点为1 925μmol/m2·s,最大净光合速率为7.661 3μmol/m2·s(图6)。
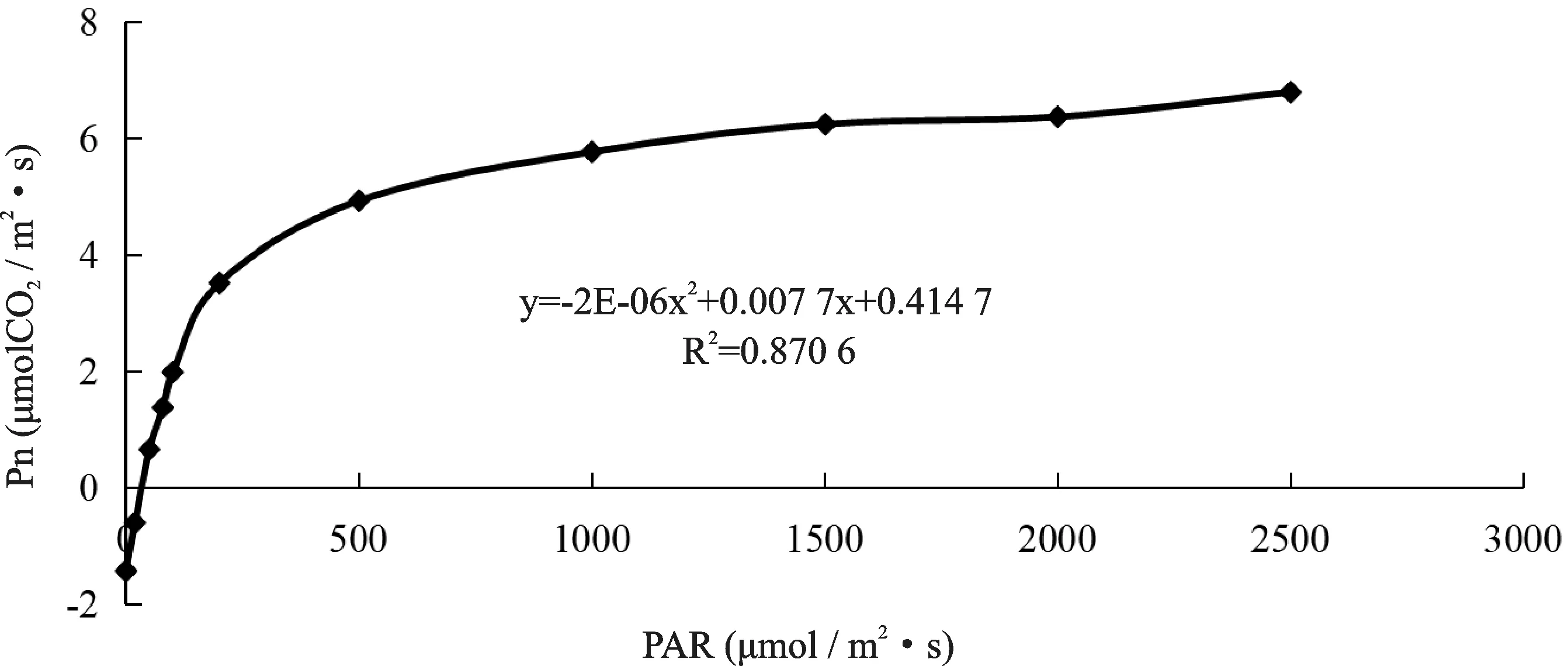
图6 欧洲山毛榉Pn与PAR的响应曲线Fig.6 Response curve of Pn and PAR
3 讨论
在光照不强或温度不高的阴雨天气,光合作用趋势通常呈单峰曲线[29]。晴朗天气呈双峰曲线变化趋势,午间高温会影响反应酶活性,导致叶绿体结构变化甚至被破坏,从而影响Pn,为了避免高温下叶片剧烈蒸腾失水,气孔部分关闭导致CO2供应不足[30]。温度过高或过低均容易使叶绿体分解,而叶绿素含量直接影响光合速率,加速叶片老化。欧洲山毛榉叶片6、8月份Pn呈现“双峰型”曲线,表现出明显的光合“午休”现象,7、9月份呈现“单峰型”曲线,不同月份Pn日变化趋势不相同。许多研究表明,引起植物Pn“午休”现象的植物自身因素分为气孔限制因素和非气孔限制因素,前者是由于气孔关闭导致外界CO2供应降低,后者是由于叶肉细胞光合性能下降[31]。只有当Pn和Ci变化相同,同时变大或者变小,且气孔定值增大时,才能认为光合速率的下降和气孔限制因素有关,否则归因为非气孔因素[32]。本试验中,6、8月份Pn变化和Ci变化趋势相反,表明8月份由于外界环境条件的变化导致了数周的净光合作用和光化学反应的减少,从而导致了整个8月份光合作用的大量丧失,加之此时由于叶片逐渐衰老使叶肉细胞光合性能下降,迫使8月份欧洲山毛榉光合作用降低。植物叶片遭受剧烈蒸腾作用后,水分补充不足,待到高温环境缓解后,树木能够迅速恢复光合速率,但完全恢复到预应力水平需要较长的时间,因为一定程度的气孔和光化学限制可能持续存在[33-34];事实上,叶片的光合作用具有典型的季节性寿命,在此之后,即使环境条件仍然有利,光合作用也会下降,衰老也会开始[35-38]。
本试验中6月Pn值比较高,这时的环境条件适合欧洲山毛榉进行光合作用,是物质积累、光合作用产物形成的重要时期,此时要加强田间管理,充分发挥其Pn高的优势。7月、8月高温、高湿、强光的环境不适宜欧洲山毛榉光合作用,Pn呈下降趋势。8月Pn总体水平略高于9月,但实际Pn值已经较低,且光饱和点相对较低这一结论也得到了印证。4个不同月份测定的数据表明,上午8:00前后欧洲山毛榉光合作用较强,此时的Pn、Tr、Gs均处于全天最大值。欧洲山毛榉的Pn、Tr、Gs大小顺序表现为6月>7月>8月>9月,这说明欧洲山毛榉6月份生长速度高于其他月份。因此,在栽培管理中,要更加注重6月份的水肥管理,以便增加当年的生长量。
光合作用受光照、温度、环境CO2浓度等内在环境因子的变化影响大[39];Pn直接反映叶片光合能力大小[40];Cs反映叶片对外界气体的交换能力[41];Ci反映植物利用CO2能力,Ci越小,光合作用越强[42]。CO2在叶肉中的传导性被认为是光合作用变化的进一步原因,光合作用变化是对环境驱动的内部植物水分平衡波动的反应[43-44]。本试验中,6月份的Ci显著低于9月份的Ci,说明6月份的光合能力显著高于9月份,这可能是因为9月份气温较高造成的,气孔部分关闭引起的光合作用减弱,因此在炎热的夏季应注意采取保障园区通风措施以保障CO2的供应。气孔作为叶片与外界进行气体、水分交换的场所,在许多植物中都发现了Cs和Pn的相关性[45]。董星光等(2015)研究发现我国野生山梨光合速率与气孔导度呈显著的指数曲线关系,山梨光合速率主要受气孔限制的影响[46],这与本文所述7月份欧洲山毛榉光合速率与气孔导度呈极显著正相关一致;Gs是影响植物水分代谢的最主要因素,进而影响到Pn,6月份测试的净光合速率、蒸腾速率和气孔导度与7、8和9月有显著性差异,呈现逐月递减的趋势,与前人的类似研究结论一致[47]。这可能是欧洲山毛榉本生长在海拔较高地区,移栽到海拔较低地区时不耐高温,使叶片气孔闭合,导致净光合速率、蒸腾速率和气孔导度降低,这就造成光合作用逐月减弱,呼吸作用逐月增强,导致胞间CO2浓度含量从6月到9月份逐月增加,这也说明高温对欧洲山毛榉的生长发育有很大的影响。
植物的光响应曲线对于描述植物的光化学进程中的光化学效率具有非常重要的意义[48]。表观量子效率主要反映植物在弱光下对光能感知的能力,表观量子效率代表植物叶片含有的色素蛋白复合体的多少,反映对弱光的吸收与转换能力[7]。植被的光合作用对光的响应过程,可以分为两个阶段:光合速率随光照强度增加的阶段,以及光合速率不再随光照强度增加的光饱和阶段,不同的光响应模型在表述这两个阶段有其各自的优势,NRH模型主要是针对光饱和阶段进行修正[22]。光合作用的某些生理参数尤其是光饱和点和光补偿点可以直观体现植物对光照强度的需求适应反应,可为植物在实际栽培管理中提供理论基础[49]。从欧洲山毛榉光合作用及光的反应曲线特征来看,欧洲山毛榉的光饱和点较高,为1 925μmol/m2·s,光饱和时Pn为7.661 3μmolCO2/m2·s,光补偿点较低为36.614 μmol/m2·s,加之6至7月份为欧洲山毛榉主要生长发育期,在此期间应当合理施肥,避免过度“午休”拉取遮阴网,减小光抑制现象,增加欧洲山毛榉旺季的生长时间,本试验为欧洲山毛榉更好的在不同地区进行推广和种植提供科学理论依据。
