宁夏河东沙地生物土壤结皮对土壤性质及入渗过程的影响
2022-09-26董智今张呈春展秀丽
董智今,张呈春,展秀丽
宁夏大学地理科学与规划学院, 银川 750021
生物土壤结皮(BSCs)是细菌,真菌,地衣,藻类和苔藓等生物同表层土壤颗粒相互作用形成的复合体[1—2],是干旱区地表主要的覆盖形式[3—4],其存在改变了土壤理化性质[5—7]、土壤生态[8]及水文过程[9],对荒漠生态系统的物质与能量流动产生重要影响。随着生物土壤结皮的发育与演替,其下伏土壤对降水的再分配和储存利用方式发生明显改变[1,10—11],进而形成了有别于原有荒漠生态系统的独特的生态水文过程[2—3,12]。研究沙区不同生物土壤结皮对土壤性质以及水分入渗的影响,对干旱区生态系统修复与保护意义重大。
近年来,众多学者开展了关于生物土壤结皮对土壤影响的研究,主要包括了不同类型(发育程度)生物土壤结皮对土壤理化性质[5—6,13—17]、水分入渗[10—12,18—20]、水分动力学[18,21—22]、水汽凝结与蒸发[23]等的影响研究,以及冻融交替[24]、降水冲刷[25]、踩踏[4,20]、植物根系[21,26—27]等干扰对生物土壤结皮渗透性影响的研究,研究区重点集中在黄土高原[8, 18,21—23]、青藏高原[28—29]、腾格里沙漠[13—14,30—31]、毛乌素沙地[11,15]等生态脆弱区,其中生物土壤结皮能够显著影响土壤性质已成为研究共识[2,13—17],但有关生物土壤结皮对土壤入渗影响的研究结果仍不尽相同,例如由于受多种气候以及微环境等因素的影响,生物土壤结皮具有发育复杂、空间异质性等特点,使得其对土壤水分入渗产生促进[27—28,31]、阻碍[18,30]和无显著作用[25]这三方面的影响,因此,定量化描述不同区域生物土壤结皮对水分入渗的影响仍是目前研究的重点。本研究区固沙植被主要为灌木、生物土壤结皮与一年生草本植物,其中发育晚期的藓类结皮与草本植物混生代表着沙区植被-土壤系统处于良好的恢复和相对稳定的阶段,这增加了沙区土壤生态水文过程的多样性与复杂性,因此研究草本与生物土壤结皮混生对土壤水分入渗过程的影响具有重要意义。
宁夏河东沙地一直是国家和宁夏生态治理的重点区域,近年来,该地区防沙治沙工作取得了重大效果,地表覆被面积增加显著。但是目前有关该区域生态系统的深层研究较为薄弱,特别是沙漠化治理过程中生物土壤结皮的形成及覆盖对下伏土壤性质以及对土壤水分再分配的影响研究较少。本文选择由不同类型生物土壤结皮(藻类结皮、藓类结皮和草本-藓类结皮)覆盖下的风沙土以及流动沙丘作为对照,采用双环入渗试验和模型模拟,对3种生物土壤结皮和裸沙的土壤性质和入渗特征展开研究,以期为该区域建立合理稳定的固沙植被,实现沙区生态系统的修复与重建提供基础数据与理论借鉴。
1 研究地区与研究方法
1.1 研究区概况
宁夏灵武市白芨滩(37°49′05″—38°20′54″N,106°20′22″—106°37′19″E)位于毛乌素沙地西南缘,面积748.43 km2。该保护区属于中温带干旱气候区,夏季炎热干燥,冬季寒冷,年均温10.4 ℃,年均降水量206.2—255.2 mm,年均蒸发量1933.3 mm。研究区地带性土壤以灰钙土为主,非地带性土壤主要为风沙土,土壤结构松散。植被以一年生草本植物、沙地植被和荒漠植被为主,代表性植物群落有柠条(Caraganakorshinskii)、猫头刺(Oxytropisaciphylla)、花棒(Hedysarumscoparium)等。生物土壤结皮是该地区固定与半固定沙丘常见的土表微景观,研究区内生物土壤结皮在固定沙丘的迎风坡上优先发育,丘间低地广泛发育斑块状藓类结皮,坡顶与迎风坡灌丛植被间有连片发育的藻类结皮。研究区藻类与藓类结皮的优势种为具鞘微鞘藻(Microcoleusvaginatus)、真藓(Bryumargenteum)[32]。
1.2 试验方案
2021年10月,在白芨滩自然保护区选择地势较平坦、生物土壤结皮覆盖度广(>90%)的区域作为试验观测点,分别为无结皮覆盖的裸沙(BS)、发育早期藻类结皮覆盖土壤(AC)、发育晚期藓类结皮覆盖土壤(MC)以及藓类结皮与草本植物混生覆盖土壤(H-MC),每个试验点面积为25 m2(5 m×5 m)。其中位于固定沙丘顶的AC样地中藻类结皮相对盖度>85%,结皮厚度为2.95—3.47 cm,表面较光滑,颜色为浅灰色;位于丘间平地的MC样地中藓类结皮相对盖度>80%,结皮层厚度为10.08—11.48 cm,表面较粗糙,颜色较深;H-MC样地为柠条灌丛下高覆盖度(>85%)的藓类结皮,地表生长一年生杂草(覆盖度为60%)。
土壤水分入渗采用双环入渗仪(QT-IN12-W,北京渠道科学仪器有限公司,中国)测定,双环入渗仪内环直径为20 cm,外环直径为35 cm,高度为35 cm。在试验点内,针对AC、MC和H-MC,分别选择藻类结皮、藓类结皮和草本-藓类结皮完全覆盖的区域(结皮覆盖面直径> 35 cm)作为入渗试验样点,将双环放置在样点内垂直地面下埋15 cm,并使用大孔隙单层纱布覆盖保护表层结皮。由于本研究区沙土初始入渗过快,仪器所配马氏瓶不能满足供水需求,因此使用2个1000 ml标准量筒分别向内环和外环同时均匀供水,当内外环中的水层高度同时达到3 cm时[33],开始计时并读取内环相应时间内增加的水量。记录不同时段的入渗水量:0—2 min每隔30 s记录1次,2—12 min每隔1 min记录1次,14—22 min每隔2 min记录1次,27—67 min每隔5 min记录1次,试验过程中内外环水面固定保持在3 cm高度以此来保持水头稳定[33]。当至少3次相同测定时间内供水量恒定时可视为达到稳定[22,33],停止试验,测定入渗时间不少于65 min。
由于本研究区沙土入渗过快,野外试验中初始入渗量存在误差,参考前人研究[10—11,21—22],本文将入渗前3 min的入渗速率定义为初始入渗速率;将单位时间内的入渗量趋于稳定时的入渗速率定义为稳定入渗速率;将达到稳渗时的累积入渗量与达到稳渗所用时间的比值定义为平均入渗速率;将60 min内的累积入渗总量定义为累积入渗量[22],入渗速率计算公式如下:
(1)
式中:Vn为一定温度下n时刻的土壤入渗速率(mm/min);Ln为n时刻的内环供水量(cm3);S为内环面积(cm3);10为cm与mm的转换系数;Δn为观测间隔时间差(min);T为时段内的平均水温(10 ℃)。
1.3 样品采集
采用烘干法测定土壤含水量[23],入渗试验前在每个样地用铝盒分别采集0—1 cm、1—10和10—20 cm深处土壤样品,每个深度重复取三次,共取铝盒样品36个;采用环刀法测定土壤容重[10],入渗试验前在每个样地分别取表层,10 cm和20 cm处取环刀样品,每个深度重复取三次,共取环刀样品24个;采用游标卡尺测量生物土壤结皮厚度(取3—5次重复)[21]。
入渗试验前在样地内选取3—5个采样点进行分层采样,分层为生物土壤结皮层(0—1 cm)和结皮覆盖下土壤(1—10和10—20 cm),同层样品进行混合并装袋后带回室内备用。
1.4 实验室分析
土壤饱和含水量与土壤孔隙度在室内测定[34];采用重铬酸钾外加热法测定土壤有机质含量[16];马尔文激光粒度仪(Mastersizer 3000,Malvern Instruments Ltd.,UK)测定土壤机械组成。
1.5 土壤水分入渗模型
模型模拟公式选取Philip、Kostiakov和Horton 3种常用的入渗模型[35],对入渗过程模拟并评价其适用性。
Philip模型:
f(t)=0.5s/t1/2+A
(2)
Kostiakov模型:
f(t)=b/ta
(3)
Horton模型:
f(t)=fc+(f0+fc)/ekt
(4)
式中:f(t)为入渗速率(mm/min);f0为初渗速率(mm/min);A和fc为稳渗速率(mm/min);t为入渗时间(min);s为吸湿率(mm/min1/2),表征土壤入渗能力;a,b为常数,分别表征第一个单位时间段内的平均入渗速率与入渗速率随时间减小的程度;k为参数,表征入渗速率随时间减少的程度。
1.6 数据处理
使用Microsoft Excel 2019对数据进行处理与分析,IBM SPSS 24. 0进行单因素方差分析(one-way ANOVA)、Pearson相关分析(α=0.05)和逐步回归分析(α=0.1),通过Origin 2018作图。
2 结果与分析
2.1 不同生物土壤结皮对土壤性质的影响
从表1可知,研究区0—20 cm藻类、藓类、草本-藓类3种类型生物土壤结皮覆盖下土壤以及裸沙的机械组成均以砂粒为主(>90%)、粉粒含量较少(<9%)、黏粒含量极少或无(<0.02%)。生物土壤结皮覆盖的表层土壤与裸沙相比,粉粒含量增多(3.6%—5.8%);极细砂粒含量增多(0.4%—1.7%);细砂含量减少(7.8%—33.7%)且藓类结皮的细砂粒含量显著减少;中砂含量增多(2.6%—20.9%);粗砂含量增多(1.2%—8.1%)且草本-藓类结皮的粗砂含量显著增加。生物土壤结皮的存在,使得土壤粒径分布更加均匀:粉粒、极细砂、中砂和粗砂含量均有所增加,细砂含量减少。

表1 不同发育程度生物土壤结皮覆盖下土壤的颗粒组成
如表2所示,随着生物土壤结皮的发育,结皮厚度显著增加。与裸沙相比,藻类与藓类结皮覆盖下土壤容重变化不显著,藓类结皮初始含水量增加了2.4倍,藻类结皮初始含水量变化不显著,土壤总孔隙度分别减少了4.0%和6.5%,饱和含水量分别减少了6.8%和11.1%,土壤有机质含量是裸沙的5—6倍。草本-藓类结皮覆盖下土壤与裸沙相比有机质增加了6倍,总孔隙度增加了9.1%,饱和含水量增加了12.6%,初始含水量变化不显著。
2.2 不同生物土壤结皮覆盖下土壤入渗特征
由图1可知,不同生物土壤结皮覆盖下土壤水分入渗速率均呈现出先降低再波动最终趋于稳定。4种样地入渗速率之间差异显著(P=0.036<0.05),累计入渗量表现为H-MC>BS>AC>MC,表明发育程度更高的藓类结皮能够显著降低土壤入渗特性,而草本与藓类结皮混生的复合表层,由于植物根系的存在入渗速率与累计入渗量显著增加。
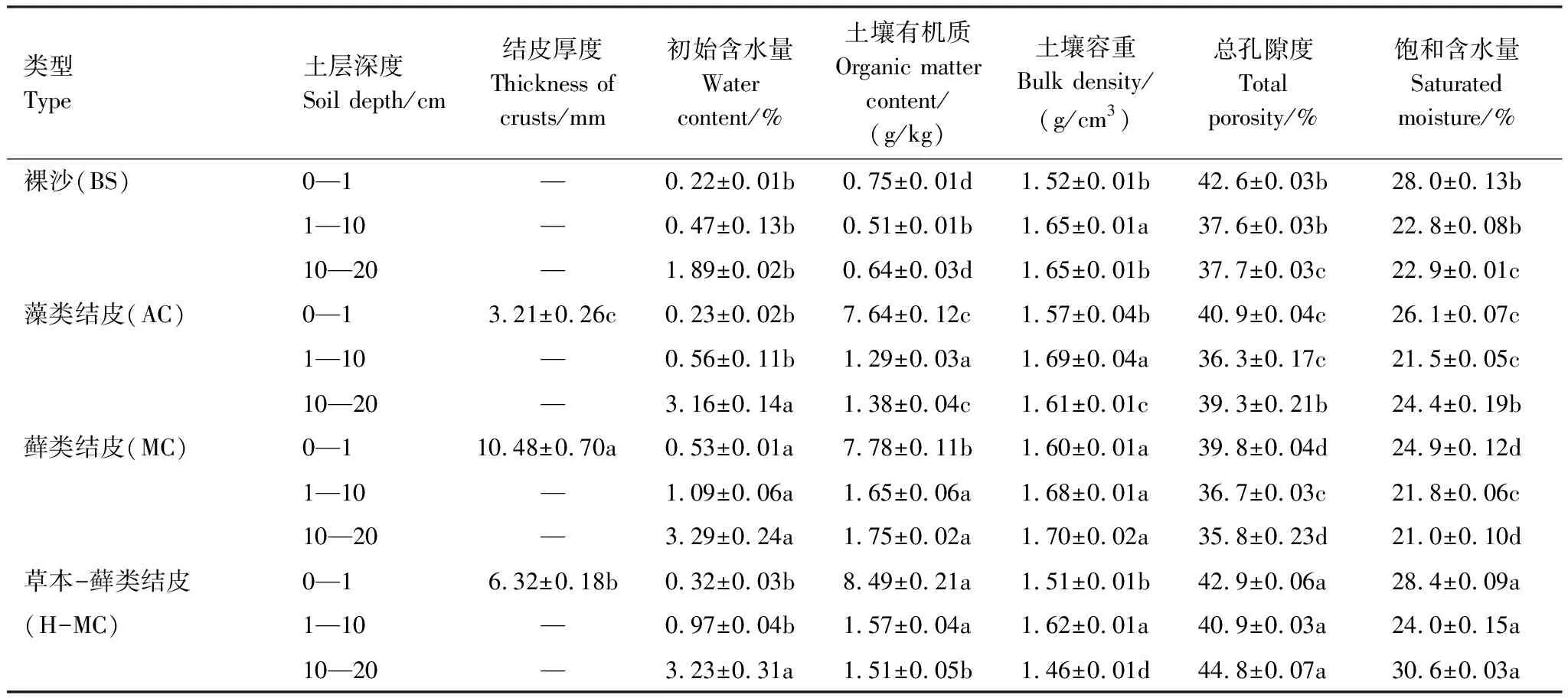
表2 不同发育程度生物土壤结皮覆盖下的土壤性质

图1 不同发育程度生物土壤结皮覆盖下土壤的水分入渗过程与累计入渗量Fig.1 Infiltration rate /cumulative infiltration volume of the biocrusts in different developmental degrees
由图1可知,土壤入渗可分为初期瞬减、中期波动和后期稳定三个阶段。如表3所示,第一阶段,裸沙初渗速率显著高于有结皮覆盖的土壤 (P<0.05),表明藻类、藓类和草本-藓类结皮的存在明显降低了初渗速率,降低幅度依次为14.3%、37.2%、11.8%;第二阶段入渗速率变化剧烈(图1),裸沙的入渗速率仍然显著高于发育程度较高的藓类结皮;第三阶段,由于此时土壤孔隙被水分充满达到饱和进入稳渗阶段,与裸沙相比,藻类和藓类结皮覆盖下土壤达到稳定入渗的时间更长,表明藻类和藓类结皮能明显降低土壤稳渗速率,降低幅度分别为14.4%和18.3%,草本-藓类结皮覆盖下土壤入渗达稳定的时间则变短,稳渗速率较裸沙增加了4.5%。表明生物土壤结皮的发育程度越高,阻止土壤水分的入渗能力越强,而草本-藓类结皮复合体则会促进土壤水分入渗。
2.3 影响土壤入渗性能的因素
如表4所示,土壤入渗性能与土壤理化性质的相关性表现为:初渗速率与结皮厚度和初始含水量呈显著负相关,与其余因素相关性不显著;稳渗速率、累计入渗量均与土壤容重呈现极显著负相关,与土壤总孔隙度以及饱和含水量呈极显著正相关,与结皮层厚度、土壤机械组成和有机质相关性不显著;平均入渗速率与土壤容重呈显著负相关,与总孔隙度与饱和含水量呈显著正相关,与其他因素相关性不显著。

表3 不同发育程度生物土壤结皮覆盖下土壤的水分入渗参数

表4 土壤入渗性能与影响因素的相关性
在此基础上分别对初渗速率与稳渗速率进行逐步回归分析可得出:
Ii= -2.066 TOC+14.197SM+12.676BD+17.500 (R2=0.999)
(5)
Si= -1.202BD+0.225WC+ 0.00125C+9.055 (R2=0.999)
(6)
式中:Ii为初渗速率(mm/min);Si为稳渗速率(mm/min);TOC为结皮厚度(mm);SM为饱和含水量(%);BD为土壤容重(g/cm3);WC为初始含水量(%);C为黏粒含量(%)。
由方程(5)可知,初渗速率主要受结皮厚度、饱和含水量和土壤容重影响,结皮层越厚、饱和含水量越低与土壤容重越大均会阻碍土壤的初始入渗能力;由方程(6)可知,稳渗速率主要受土壤容重、初始含水量和黏粒含量影响,土壤容重越大、初始含水量越低、黏粒含量越少则会降低土壤的稳定入渗能力。
2.4 不同生物土壤结皮覆盖下土壤入渗过程模拟
由表5可以看出,不同模型对于土壤水分入渗过程的拟合效果不同。Philip模型拟合结果显示,拟合A(稳渗速率)值为4.86—6.65 mm/min与实际的7.96—10.18 mm/min相差较大,R2均小于0.89,说明Philip模型拟合效果较差。Horton模型拟合结果显示,fc(稳渗速率)值与实测值相差很小(0.05—0.315 mm/min),k值在0.79—1.54之间,最大值出现在裸沙,最小值在藓类结皮,反映了入渗速率递减情况,与实测结果基本相符,R2均大于0.94,模型拟合效果较好。Kostiakov模型拟合结果显示,参数b值在7.74—11.72 mm/min之间,b值越大,入渗曲线的斜率越大,瞬时入渗率衰减越快,拟合出衰减快慢顺序为:BS>H-MC>AC>MC,与实测结果一致,R2均大于0.96,此模型拟合效果较好。
3 讨论
3.1 生物土壤结皮覆盖对土壤性质的影响
生物土壤结皮通过改变地表微形态使其下伏土壤的性质发生变化[5]。首先,不同发育程度的生物土壤结皮对下伏土壤机械组成影响不同。本研究发现,与藻类结皮相比,藓类结皮表层土壤砂粒含量降低,粉粒含量增加,土壤结构细化,颗粒物分布更均匀,这是因为在藓类结皮以及草本-藓类结皮混生覆盖下,藓类与草本使得土壤表面粗糙度加大,对于细颗粒物的拦截作用明显提高,这与高广磊等[15]在宁夏盐池县以及孙福海等[18]在黄土高原的研究结果一致。

表5 不同发育程度生物土壤结皮覆盖下土壤入渗过程的模型模拟结果
其次,不同发育程度的生物土壤结皮对下伏土壤总孔隙度和土壤含水量影响不同。本研究发现草本-藓类结皮覆盖下土壤总孔隙度较高,说明草本植物根系对结皮下伏土壤的影响显著。藓类结皮覆盖下土壤表层含水量最高,这是因为在本研究区藓类结皮广泛分布于丘间低地,灰黑色藻类结皮分布于固定沙丘坡面-丘顶,蒸发较强,而草本-藓类结皮则是由于生长季植物需水,使得下伏土壤表层含水量与裸沙相比差异较小。
最后,不同发育程度的生物土壤结皮对下伏土壤有机质影响不同。本研究发现藻类和藓类结皮的覆盖使得表层土壤的有机质含量增加,一方面归因于生物土壤结皮的覆盖使表层土壤结构优化,利于土壤团聚体形成,增加有机质含量[5];另一方面则由于生物土壤结皮可以为微生物的生长提供环境,进而提升下伏土壤的有机质含量[13]。
此外,生物土壤结皮对土壤性质的影响还具有“表层性”[36],即生物土壤结皮和下伏土壤的双层结构,使得结皮对土壤性质的影响主要集中在表层[17,21],这在本研究中也得到了证实。
3.2 生物土壤结皮覆盖对土壤水分入渗的影响
本研究发现藻类和藓类结皮明显阻碍了土壤水分入渗过程,且生物土壤结皮发育程度越高,土壤渗透性能越低。这是因为,第一,生物土壤结皮截留的细粒物质填充土壤孔隙,降低了孔隙间的连通性[37—38];第二,本研究区内高盖度藻类和藓类结皮中的藻丝体、真菌菌丝、藓类假根与砂粒缠绕胶结增加了结皮层厚度[22],结皮层吸水性增强进而阻碍水分的入渗[21],这与Xiao等[37]研究结论一致,并且与裸沙相比,藓类结皮覆盖下表层土壤初始含水量明显增加,降低了沙土的土壤水力梯度,进而减缓了初渗速率[19];第三,生物土壤结皮覆盖增加了表层土壤有机质,且藓类植物体分泌的胞外多糖等化学物质堵塞土壤孔隙进而降低入渗速率[11]。最后,本研究中藓类较藻类结皮表现出更高的阻渗能力,这与Zhang等[14]研究结果不同,因为本研究选取地势平坦区域作为观测试验点,无地形因素对结皮层的影响,而且本研究发现藓类结皮能够完全覆盖表土层孔隙,藓类植物的毛尖、茎叶微结构使得土壤更封闭疏水性更强,这对入渗的阻碍作用更显著[26]。
本研究中还发现草本-藓类结皮复合体能够有效的促进土壤水分入渗,这是因为,草本植物为发育在柠条灌丛下一年生的杂草,受到杂草和柠条根系的影响,表层结皮层被破坏,水分运移方式被改变[39],同时,植物根系在土壤中交错生长分布,形成土壤大孔隙,提高了土壤的渗透性能[40]。李渊博等[21]在黄土高原的研究也表明草本植物和藓类结皮混生覆盖下会对水分入渗有一定的促进作用,与本研究结果一致。
3.3 土壤入渗性能差异的因素分析
本研究发现,不同发育程度的生物土壤结皮入渗性能的差异与土壤性质密切相关,并且在不同的入渗阶段,土壤性质对其影响也各不相同。在入渗的初始阶段,结皮层越厚,其吸水性越强,入渗速率也越低,而且初始阶段土壤初始含水量越多,水力梯度越小入渗速率也越低[19]。在本研究中藓类结皮的覆盖,明显提升了结皮层厚度与土壤初始含水量,从而减小初渗速率,这与王闪闪等[41]研究结果一致。达到稳渗阶段时,入渗速率与土壤容重、总孔隙度与饱和含水量呈现极显著相关性,在该阶段,生物土壤结皮对入渗的影响减弱,而下伏风沙土的性质以及植物根系产生的裂隙则成为影响入渗的主要因素[39,42]。
本研究进行逐步回归分析后发现,结皮层厚度是影响初渗速率最重要因素,而土壤容重则是影响稳渗速率最重要因素,这进一步说明,入渗时间越长,生物土壤结皮对入渗的影响越小,而土壤性质以及植物生长产生的土壤裂隙对入渗的影响越大。
3.4 入渗模型适用性评价
本研究中采用了三种经典的入渗模型来模拟生物土壤结皮覆盖下水分入渗规律,结果发现,Kostiakov模型对不同样地入渗过程的模拟效果最好(R2>0.96),拟合出入渗速率的快慢为:BS>H-MC>AC>MC,与实测结果一致。而Philip模型是物理模型,与有结皮覆盖的土壤相比,裸沙的水分入渗过程拟合效果更好(R2=0.89),说明该模型不适用于模拟生物土壤结皮覆盖下的非均质土壤,更适用于均质土壤的一维入渗[10]。 Horton模型属于纯经验公式[15],对4种样地类型拟合效果均较好(R2>0.94),对初始入渗速率值拟合精度高,但是对稳定入渗速率的拟合精度不够。
综上所述,研究区内生物土壤结皮覆盖改变了下伏土壤的性质,进而影响了土壤水分入渗过程。在对该区域生物土壤结皮的研究中,未来还需要深入探讨固沙植物根系对水分入渗过程的影响,进一步开展生物土壤结皮对该区域土壤水文过程的研究。
4 结论
(1)生物土壤结皮对浅层土壤性质影响明显。相较裸沙,生物土壤结皮覆盖下的土壤砂粒含量减少,粉粒含量增加,土壤有机质含量增加;藓类结皮土壤初始含水量最高;藻类和藓类结皮覆盖下土壤总孔隙度与饱和含水量降低,草本-藓类结皮其下伏土壤总孔隙度与饱和含水量增加。
(2)生物土壤结皮改变下伏土壤性质进而对风沙土的入渗性能够产生显著影响。生物土壤结皮发育程度越高,阻止土壤水分的入渗能力越强,而草本-藓类结皮复合体则促进土壤水分入渗;在入渗初期,结皮层厚度、土壤初始含水量是影响入渗的主要因素,而在稳渗阶段,土壤容重则是影响入渗特征的主要因素,且植物根系的存在减小了生物土壤结皮的阻渗作用。
(3)Kostiakov模型最适用于模拟本研究区不同发育程度生物土壤结皮覆盖下土壤水分入渗过程,Horton模型次之,Philip模型较差。
