小兴安岭白桦次生林内红松光合对林隙大小与林隙内位置的适应性研究
2022-09-26田博宇牟长城李佳欣
田博宇,牟长城,李佳欣,叶 林
(1.东北林业大学 林学院生态研究中心,黑龙江 哈尔滨 150040;2.黑龙江省伊春市林业科学院,黑龙江 伊春 153000)
林隙在森林恢复和管理中发挥着重要作用[1-2]。林隙不仅为种子萌发和幼苗生长提供机会,从而影响林分结构[3-4],而且在维持种群动态和增加森林生态系统多样性方面也具有重要作用[5-8]。
林隙大小和林隙内位置是林隙的两个关键特征,直接影响到达地面的太阳辐射量,引起林内微气候和土壤养分可利用性变化,进而对树种更新与幼苗生长起着至关重要的作用[9-12]。现有研究表明,北方森林在林隙(20~5 000 m2)内更新的幼苗种类较郁闭林冠下和裸地上更加丰富,且在小林隙(300 m2)内各位置上的更新组成相似,在大林隙内自光照充足北部到遮阴南部更新种数呈增加趋势[13]。各树种更新对林隙大小需求也不同,不耐阴种或早期演替种通常只占据较大的林隙[14-16],其更新所需最小林隙为1 000 m2或400 m2[17-18];而耐阴种或后期演替种在较小或较老的林隙中更新与生长更好[15-16,19]。不耐阴物种更新最理想林隙直径/周围树高的比值d/h>2,中等耐阴物种d/h在1.0~2.0之间,而耐阴物种d/h<1.0[20]。可见,早期研究多集中于幼苗的更新与生长对林隙响应方面,缺乏有关幼树光合与生长对林隙响应方面的认识。
近年来进一步从生理生态方面探寻了林隙内幼苗光合和生长对林隙微环境的响应规律及机制。研究发现寒温带落叶林[21]和热带雨林[22]中多个树种的幼苗在林隙内光合速率与生长明显高于林内,但不同树种随林隙的增大却表现出不同的光合适应性,有些种光合速率随林隙的增大而提高,而有些种则变化不大;北方森林中红云杉Picea rubens的幼苗在中等大小林隙(100~340 m2)内生理生态指标与生长指标均最好[23-24];温带次生林中蒙古栎Quercus mongolica、五角槭Acey mono和紫椴Tilia amurensis幼苗光合与生长对林隙大小及林隙内位置的适应能力也有所不同,不耐阴的蒙古栎幼苗响应最为敏感(光合能力在大林隙的中心位置最强,生物量在大林隙内最高),中等耐阴性的槭树比较敏感(仅生物量在大林隙内最高),而耐阴性强的椴树则不敏感[25];核桃楸Juglans mandshurica幼苗的净光合速率在大、中林隙显著高于小林隙和林内,且在各大小林隙内的中心区和过渡区显著高于边缘区,但红皮云杉Picea koraiensis幼苗的净光合速率却不随林隙大小及林隙内位置的变化而改变[26],并发现早期演替种主要以形态调整(叶厚度、比叶面积及气孔密度等)来适应林隙内强光,而后期演替种则以生理过程调整(超微结构变化调整光合过程)来适应强光[27-29]。可见,这些研究多是针对更新幼苗光合和生长对林隙环境的响应,而有关幼树阶段光合和生长对林隙环境的响应研究甚少,仅见有温带长白山原始红松林中红松低龄阶段幼树(15 a)最大光合速率随林隙增大而增大至林隙面积267 m2时开始降低[30],而对于东北林区次生林通过“栽针保阔”途径恢复的阔叶红松林中较大龄阶段红松幼树(30 a)光合与生长对林隙大小及林隙内位置是如何响应的目前仍不清楚,这势必会限制地带性顶极植被阔叶红松林恢复经营理论与技术的发展。
本研究以我国东北温带小兴安岭中期“栽针保阔”红松林(白桦次生林林冠下栽植红松35 a)为研究对象,在生长季同步测定不同大小林隙(大,201~211 m2;中,169~174 m2;小,100~102 m2)内各位置(中心区、过渡区、边缘区)和林内的红松幼树最大净光合速率、蒸腾速率和气孔导度等光合响应参数及其微环境因子(透光率、叶片温度等),揭示白桦次生林内红松幼树在不同林隙大小和林隙内位置的长期光合适应规律及其适应机制,为次生林恢复地带性顶极植被阔叶红松林经营管理提供科学依据。
1 研究区概况与研究方法
1.1 研究区概况
研究区域位于黑龙江省伊春市带岭区大青川林场353林 班,地理坐标为47°0′50″N、129°09′48″E,平均海拔高度为225 m,该区域为温带大陆性季风气候,冬季寒冷干燥,夏季温热多雨,温差较大,年均最高气温7.5℃,年均最低气温-6.6℃,年均气温-0.3℃,年均降水量为676 mm[31]。地带性土壤为暗棕壤。地带性植被为原始阔叶红松林,其蓄积量高(一般为400~500 m3/hm2,高者可达700~800 m3/hm2),生态功能强,稳定性好[32],但因过度开发利用,原始阔叶红松林已退化为低质、低效次生林和人工林(91.3 m3/hm2和44.9 m3/hm2)(前八次森林资源清查平均数据)[32-34],故如何恢复阔叶红松林已成为本区亟待解决的重大科学问题[32]。本文的研究对象为中期“栽针保阔”白桦红松林,1983年在白桦次生林(10 a)下人工栽植红松(2 000~2 500株/hm2),并于1989年对上层阔叶树进行了不同强度的透光抚育(透光抚育强度为蓄积比1/7~1/4),并由上层阔叶树种白桦Betula platyphylla和山杨Populus davidiana形成了不同大小的林隙微环境;2018年开展本试验时,白桦次生林林龄为45 a,林下红松为35 a,且红松幼树在各大小林隙内及其林隙内的不同位置(中心区、过渡区和边缘区)均有分布,加之红松幼树长期处于不同的林隙微环境之中,红松林木在生长方面存在着较大的差异性,这为研究红松光合与生长对林隙大小和林隙内位置的长期适应规律提供了比较理想的场所。
1.2 样地设置
本研究以伊春市带岭区大青川林场的中期“栽针保阔”白桦红松林为研究对象,在已建立长期试验地的基础上,选取立地条件相近区域(阳坡下坡位),依据扩展林窗的定义(由林窗空隙周围树木的树干围成的土地面积或空间[35-36])及林分现实状况设计了3个不同大小的林隙和林内(图1)(大、中、小林隙和林内的面积依次为201~211、169~174、100~102和50~52 m2;透光率依次为37%、31%、28%和9%)4种处理,每个处理3次重复,共计12个样地;在各林隙样地内按照林隙的中心区、过渡区、边缘区(按照半径等分方法划分林隙各位置)选择红松样木3株,林内则随机选择红松样木3株,3次重复,共计选择红松样木36株,测定其生长季的光合参数及其微环境因子,揭示红松光合对林隙大小及林隙内位置的适应规律及适应机制。各林隙样地的具体状况见表1。

表1 各林隙样地基本特征†Table 1 Basic characteristics of various forest gaps
1.3 测定方法
1.3.1 光合作用的测定
光合测定时间分别为2018年4月20日—5月20日(春季)、6月20日—7月20日(夏季)、8月20日—9月20日(秋季),每月测定1次(取树冠中部同一高度的向阳方位和背阴方位叶片),每次3 d,为保证试验数据的准确和试验的顺利进行,测定时间为8:30—12:30,尽量选择在晴朗适宜的天气,第1天按照中心区、过渡区及边缘区的顺序,第2天顺序相反,第3天按照过渡区、边缘区及中心区的顺序重复测定各林隙样地内不同位置和林内的红松光合参数。
利用汉莎科学仪器公司生产的CIRAS-2光合测定仪进行测定,使用仪器内的红蓝光源提供标准光强,光照强度梯度自低到高设定为0、100、200、400、600、800、1 000、1 200、1 400、1 600、1 800、2 000 μmol·m-2s-1,测量前等待机器稳定,稳定后每15 s记录一个数值,每个梯度测定3组数据,计算平均值作为最后光合参数指标的值。测量的光合参数值包括叶片单位面积的净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(GS)、叶片温度(LT)。测定后取阴阳面叶片数据的平均值,然后将春、夏和秋三季数据加以平均得到生长季平均光合数据。再通过非直角双曲线模型[37]拟合光合光响应曲线,得到其最大净光合速率(Pmax)、暗呼吸速率(Rd)、表观量子效率(α)、光补偿点(LCP)和光饱和点(LSP),而其水分利用效率(WUE)按照公式(1)计算。

式(1)中:Pn为实际测定的光合速率;Tr为蒸腾速率。
1.3.2 透光率的测定
采用半球面影像法,利用冠层仪通过鱼眼镜头(尼康相机CoolPix 950)记录林隙内不同位置的天空影像,测量时将相机水平放置,朝向林冠上层拍摄照片。利用Digital Hemispherical Photography 4.5.2处理半球面影像,通过将林冠照片按照林隙中心区、过渡区和边缘区分成3个同心圆环,统计各个圆环中林冠空隙所占的比值,以此计算林隙内各位置及林内透光率,本研究中的林隙形状近似于椭圆形,计算公式如下:

式中:S为林隙的面积;π为圆周率;L为林隙的最长轴长度(m);W为与其垂直的短轴长度(m)。
1.3.3 比叶面积与叶绿素的测定
将测定光合作用的针叶取下并带回实验室,分两份分别测定叶面积和叶绿素含量,将测定完叶面积的叶子在80℃烘干72 h后称质量,获得比叶面积,比叶面积(SLA)=叶面积/叶片干质量。
取另一份针叶剪碎混合一起后,称取0.1 g放置于20 mL盛有90%乙醇丙酮(1∶1,v/v)混合液的试管中,将试管塞上瓶塞并放入阴暗避光的水浴锅中,100℃水浴振动12 h,直到叶片完全发白为止。测定时采用90%乙醇丙酮混合液作为空白对照,提取的浸提液用紫外分光光度计进行比色测定,在663 nm与645 nm的波长下测定提取液的吸光度值,每个样品测定3次,结果取实测数据的平均值。计算叶绿素值公式如下:

式中:A663与A645分别为提取液在663 nm与645 nm处的吸光值,公式所得的色素含量单位为mg/L。本研究测的是单位质量的叶绿素含量,因此需要换算:单位质量色素含量(mg/g)=[溶液中色素质量浓度(mg/L)×提取液总量(L)]/样品鲜质量(g)。
1.4 数据分析
数据分析采用Excel 2010和SPSS 19.0软件进行方差分析和多重比较,应用最小显著差异法(LSD)对数据组间差异进行显著性检验(α=0.05);用逐步回归方法筛选最大净光合速率的主要影响因子;图表绘制采用Sigma Plot 12.5软件完成,图中数据为平均值±标准差。
2 结果与分析
2.1 红松光合对白桦次生林林隙大小的适应
由图2A可以得到,小兴安岭白桦次生林大、中、小林隙内和林内红松生长季平均最大净光合速率(Pmax)分布在2.4~8.4 μmol·m-2s-1之间,红松Pmax在大、中、小林隙内各位置(中心区、过渡区和边缘区(小林隙除外))显著高于林内(54.2%~250.0%,P<0.05);在林隙内各位置(中心区、过渡区和边缘区)均呈大林隙(20.0%~96.4%)>中林隙(35.4%~57.1%)>小林隙的递增规律性。因此,小兴安岭白桦次生林中红松幼树(35 a)光合适应能力在林隙内强于林内;在大林隙内强于中、小林隙,中林隙又强于小林隙。
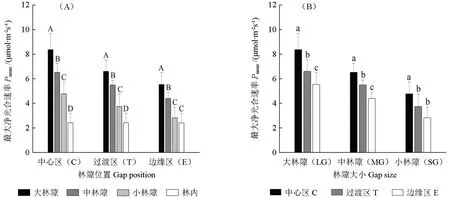
图2 小兴安岭白桦次生林不同大小林隙和林隙内位置红松幼树生长季最大净光合速率Fig.2 The maximum net photosynthesis rate (Pmax) of the P.koraiensis saplings in the forest gaps of different sizes and at different positions within forest gaps in B.platyphylla forest during growing season in the Xiaoxing’an mountains of China
由表2可知,红松光饱和点在林隙内各位置(小林隙边缘区除外)均显著高于林内(28.7%~101.0%),光补偿点却显著低于林内(27.1%~81.9%),其表观量子效率在大林隙各位置及中林隙中心区显著高于林内(69.2%~230.8%);其蒸腾速率、气孔导度和水分利用效率(除小林隙边缘区外)显著高于林内(30.8%~250.0%)。故林隙内红松的光能利用和气体交换与传输能力一般高于林内。
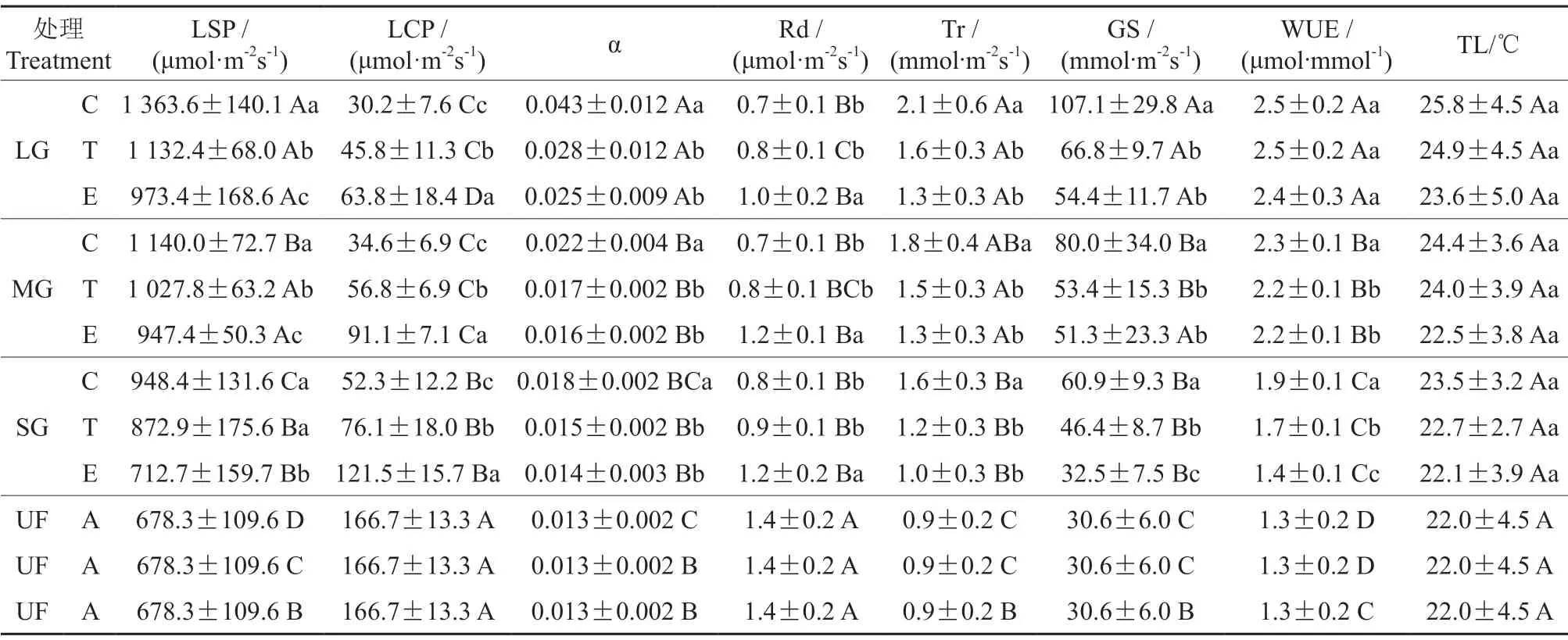
表2 红松光合参数及微环境因子†Table 2 Basic photosynthetic parameters and microenvironmental factors of P.koraiensis
在大、中林隙内各位置红松光饱和点均显著高于小林隙(17.7%~43.8%),光补偿点均显著低于小林隙(25.0%~47.5%),但仅在大林隙的中心区其光饱和点显著高于中林隙(19.8%)及在大林隙的边缘区其光补偿点显著低于中林隙(30.0%);其表观量子效率则为大林隙显著高于中林隙和小林隙(56.3%~138.9%);其蒸腾速率在大、中林隙内各位置(除中林隙中心区外)显著高于小林隙(25.0%~33.3%);气孔导度在大林隙内各位置显著高于中林隙(除中林隙边缘区外)和小林隙(25.0%~75.9%);水分利用速率在大林隙内各位置显著高于中林隙和小林隙(8.7%~71.4%)。因此,在大林隙内红松的光饱和点、蒸腾速率、气孔导度、表观量子效率和水分利用速率均最高,而其光补偿点却最低。
2.2 红松光合对白桦次生林林隙内位置的适应
由图2B得到,白桦次生林中红松最大净光合速率在林隙各位置间也存在差异性。红松Pmax在大、中林隙内呈中心区(18.2%~52.7%)>过渡区(20.0%~25.0%)>边缘区的递增规律性,但在小林隙内呈中心区(29.7%~71.4%)>过渡区=边缘区的递增趋势。因此,红松光合适应能力在白桦林各大小林隙中心区最强,过渡区强于边缘区(大、中林隙)或与边缘区相近(小林隙)。
由表2得到,红松光饱和点在大、中林隙内呈中心区(10.9%~40.1%)>过渡区(8.8%~16.3%)>边缘区,小林隙内呈中心区=过渡区(22.5%,P<0.05)>边缘区的增加趋势;其光补偿点在大、中、小林隙内均呈中心区(31.3%~62.0%)<过渡区(28.2%~37.7%)<边缘区的递减趋势;其表观量子效率在各大小林隙内均为中心区显著高于过渡区与边缘区(20.0%~72.0%)。其蒸腾速率和气孔导度在大、中、小林隙内均为中心区显著高于过渡区与边缘区(20.0%~61.5%和49.8%~96.9%);但其水分利用效率仅在小林隙内呈中心区(11.8%~35.7%)>过渡区(21.4%)>边缘区,在中林隙内呈中心区(4.5%)>过渡区=边缘区,而在大林隙内各位置间无差异性。因此,白桦林中红松光能利用能力在各大小林隙内均呈中心区>过渡区>边缘区递增规律性,其气体交换与传输能力为中心区显著高于过渡区与边缘区。
2.3 红松针叶比叶面积和叶绿素含量对林隙大小及林隙内位置的适应
由图3可知,红松针叶的比叶面积在白桦林大、中、小林隙之间或林隙内各位置之间均存在差异性。各大小林隙内各位置中红松针叶的比叶面积分布在7.15~9.06 m2·kg-1之间,其中,红松针叶比叶面积在大林隙的中心区显著低于小林隙的中心区和林内(14.9%和21.1%)(P<0.05);在大林隙和中林隙的中心区显著低于各自的边缘区(22.4%和12.7%)。因此,红松针叶形态适应仅发生在白桦林内的大、中林隙的中心区。

图3 小兴安岭白桦次生林林隙内不同位置(A)和不同大小林隙(B)中红松幼树生长季的比叶面积Fig.3 Specific leaf area of the P.koraiensis saplings in the forest gaps of different sizes (A) and at different positions within the forest gaps (B) of the B.platyphylla forest during growing season in the Xiaoxing’an mountains of China
由图4得到,红松针叶的叶绿素a、叶绿素b、总叶绿素含量在各大小林隙的中心区和过渡区以及大林隙的边缘区均显著低于林内(15.6%~47.7%);且其叶绿素a、叶绿素b、总叶绿素含量在林隙中心区为大林隙显著低于中、小林隙(14.4%~23.2%),在过渡区大、中、小林隙相近,在边缘区则为大林隙和中林隙(叶绿素b)显著低于小林隙(17.4%~20.0%和12.2%)。其叶绿素a/b值在林隙中心区为大林隙(4.5%~18.3%)>中林隙(4.4%~13.1%)>小林隙(8.4%)>林内;在过渡区为大林隙(3.5%~14.7%)>中林隙=小林隙(8.9%~10.8%)>林内;在边缘区为大林隙=中林隙(2.8%~10.8%)>小林隙(7.0%)>林内。因此,红松针叶的叶绿素含量在林隙内(各大小林隙中心区和过渡区及大林隙边缘区)显著低于林内,且其叶绿素a/b值在林隙的中心区各大小林隙间均存在显著差异性,说明红松光合对林隙大小响应存在着超微结构方面的适应性。
此外,红松叶绿素含量在林隙内各位置之间也存在差异性。其叶绿素a、叶绿素b和总叶绿素含量在大林隙内为中心区(21.1%~31.6%)<过渡区=边缘区;在中林隙内为中心区(16.6%~18.9%)<边缘区;在小林隙内则为中心区(13.4%~14.1%)<过渡区(17.6%~19.2%)<边缘区。其叶绿素a/b值在大林隙内为中心区(3.2%)>过渡区(3.5%)>边缘区;在中林隙为中心区(2.9%)>边缘区;在小林隙则为中心区=过渡区=边缘区(1.3%~1.8%)。因此,红松针叶的叶绿素含量在林隙中心区显著低于边缘区,其叶绿素a/b值在林隙中心区显著高于边缘区(小林隙除外),这也说明红松光合对林隙位置的响应同样存在着超微结构方面的适应性。
2.4 不同林隙大小及不同林隙内位置中红松光合的影响因子
由表3可知,林隙大小和林隙内位置改变了红松最大净光合速率的主要影响因子。红松幼树的Pmax在白桦次生林内仅受光饱和点所促进;在小林隙内受光饱和点和气孔导度共同促进,前者解释了Pmax的58.2%,后者解释了41.8%;在中林隙内既受气孔导度所促进,又受光补偿点所抑制,前者解释Pmax的39.1%,后者解释Pmax的60.9%;但在大林隙内其Pmax仅受蒸腾所促进。故红松幼树的Pmax在林内仅受光饱和点所促进,在小林隙内受光饱和点和气孔导度共同促进,中林隙内受气孔导度促进及光补偿点抑制,但在大林隙内仅受蒸腾所促进。

表3 白桦次生林内红松幼树最大净光合速率的驱动因子†Table 3 The driving factors of the maximum net photosynthetic rate of P.koraiensis saplings in the B.platyphylla forest
此外,红松幼树的Pmax在白桦次生林林隙内的边缘区仅受光补偿点所抑制;在过渡区仅受气孔导度所促进;但在中心区其Pmax既受蒸腾速率促进,也受叶绿素a+b促进,且蒸腾速率和叶绿素a+b分别解释了Pmax的17.7%和82.2%。故红松幼树的Pmax在白桦次生林林隙内的边缘区仅受光补偿点抑制,过渡区仅受气孔导度促进,而在中心区受蒸腾速率和总叶绿素含量共同促进。
3 讨 论
3.1 红松光合对白桦次生林林隙大小的适应
温带小兴安岭白桦次生林中人工栽植的红松幼树(35 a)生长季平均最大净光合速率在林隙内显著高于林内,且随林隙大小增大而递增,具有明显的光合适应性。这与温带次生林[26]、寒温带落叶林[21]和热带次生林[22]中多个树种的幼苗(≤8 a)及温带原始红松林内红松低龄幼树(15 a)[30]随林隙增大表现出光合适应性相类似,不同之处则在于本研究得到地带性顶极种红松在较大年龄阶段幼树时(35 a)对次生林中的林隙大小变化仍表现出较强的光合适应性,这对我国东北次生林恢复阔叶红松林具有重要指导意义。
其原因主要在于林隙大小决定林内物种光量获取[10,39],白桦林林隙内中心区、过渡区的透光率随林隙增大而递增(大>中>小>林内),而边缘区却相近(大=中=小>林内),红松幼树长期适应林隙内这种光照异质性,导致其光合能力产生有规律性的分异。其生理适应机制主要体现在两个方面:一是光能利用方面,红松幼树的光饱和点在林隙内一般显著高于林内,光补偿点却显著低于林内,且在大、中林隙内其光饱和点显著高于小林隙,光补偿点显著低于小林隙,进而使其光能利用能力在林隙内强于林内以及在大、中林隙内强于小林隙;二是气体交换与传输方面,林隙内光照增强,温度升高[40],且树木具有调整自身光合特性以适应温度的能力[41],进而保持较高的光合速率[42]。本研究发现红松同样具有调整蒸腾速率和气孔导度等生理特性以适应温度的能力,进而提高其光合速率,林隙内红松生长季平均叶片温度高于林内,且随林隙增大其提高幅度也增大(大、中、小林隙较林内最大升温幅度依次为3.8、2.4和1.5℃),这使得其生长季平均蒸腾速率、气孔导度和水分利用效率在林隙内显著高于林内以及在大林隙内显著高于小林隙,而气孔导度的增大提升了光合反应原料的吸收速率,有助于光合碳同化速率的提升[43-44],蒸腾速率加大既提供充足的光合底物—水,又能促进光合积累物的运输,进而提高净光合速率[45]。这说明我国东北温带地带性顶极种红松在较大年龄阶段幼树时其生长对策仍具有双重性(既能够在弱光条件下维持生存,又能够在强光下快速生长),这与现有树种生长对策(光合可塑性与耐阴性间存在消长关系,即耐阴种倾向于增强耐阴性而减少其光合可塑性,而喜光种倾向于增强光合可塑性而减少其耐阴性)[21-22]的假说明显不同,故有必要为次生林中较大年龄阶段红松幼树创造适宜林隙微环境,充分释放其生长潜力。
3.2 红松光合对白桦次生林林隙内位置的适应
小兴安岭白桦次生林内红松幼树(35 a)生长季平均最大净光合速率在大、中林隙内呈中心区>过渡区>边缘区及在小林隙内呈中心区>过渡区=边缘区的递增规律性,说明其在林隙内沿光照梯度仍存在光合适应性。这与蒙古栎幼苗最大净光合速率在大、中林隙内随着与林隙中心位置的距离减少而增大相类似[46]。
其原因在于林隙内光照还取决于林隙内的位置,林隙内不同位置的光照、温度、水分状况都存在差异性[47-48]。白桦林大、中林隙内的透光率呈中心区>过渡区>边缘区及小林隙呈中心区>过渡区=边缘区的递增规律性,红松幼树长期适应林隙内这种光照异质性,其光饱和点在大、中林隙内呈中心区>过渡区>边缘区及小林隙内呈现中心区=过渡区>边缘区的递增规律性,且其光补偿点在大、中、小林隙内均呈现中心区<过渡区<边缘区的递减规律性,进而使其光能利用能力沿林隙内光照梯度呈现递增规律性。与此同时,红松生长季平均叶片温度在各大小林隙内的中心区高于过渡区和边缘区(0.4~2.2℃),过渡区又高于边缘区(0.9~1.5℃),使得红松针叶的蒸腾速率、气孔导度和水分利用效率(大林隙除外)在各大小林隙内的中心区显著高于过渡区与边缘区,而蒸腾速率和气孔导度的提高不仅能够促进光合原料的吸收[44-45],而且有利于光合积累物的运输[46],进而有助于林隙中心区红松幼树光合速率的提升。可见,红松针叶的光合作用对林隙内光照异质性的适应机制与其对林隙大小所引起的光照异质性的适应机制是完全一致的,并且同样表现出生长对策的双重性(既能够忍耐林隙内边缘区弱光条件而维持生存,又能够在林隙中心区强光条件下快速生长)。
3.3 红松针叶比叶面积和叶绿素含量对林隙大小及林隙内位置的适应
在白桦次生林林隙内红松针叶比叶面积仅在大林隙内中心区显著低于小林隙和林内,且在大、中林隙内中心区显著低于边缘区,其原因在于大、中林隙内的中心区光照相对充足(透光率43.7%~54.5%),植物叶片通常会变小变厚以减少蒸腾损失增加碳固定[49];而在林内或林隙边缘区光照较弱(透光率9.2%~11.7%),在低光环境下植物叶片可分配到更多的生物量以增加受光面积[49-50]。这也说明红松叶片的形态适应仅在大、中林隙的中心区表现得更加明显。
红松针叶的叶绿素a、叶绿素b和总叶绿素含量在次生白桦林中各大小林隙内(中心区和过渡区)均显著低于林内;在大林隙内(中心区和边缘区)显著低于小林隙;且在各大小林隙内的中心区显著低于边缘区,同样表现出明显的超微结构方面的适应性。这源于红松幼树主要是通过增加其针叶的叶绿素含量来提高其在林内、小林隙及林隙内的边缘区等弱光环境下的光能捕获能力,这与亚热带常绿阔叶林中栲树幼苗增加叶绿素含量适应弱光环境[51]完全一致。但其叶绿素a/b值在各大小林隙内均高于林内,在大林隙内各位置均高于小林隙,且在大、中林隙内的中心区均显著高于其边缘区,这可能与光照强度在林隙内强于林内,大林隙强于小林隙以及林隙内中心区强于边缘区有关,这与现有研究结论在强光下生长的叶片一般具有较高的叶绿素a/b值,有助于叶片在光照充足条件下获得更多的碳[52]相一致,而在低光环境下较低的叶绿素a/b值,则有助于叶片充分利用光能,维持光系统间的能量平衡[53]。本研究中只检测到了35年生的红松幼树针叶形态及超微结构的适应性,而对其幼苗阶段是否存在的光合形态及超微结构适应性尚需要开展研究,然而除了检测光合作用外,根系养分状况也可能很好地预测林隙中的物种分化和再生性能。
3.4 不同林隙大小及林隙内位置中红松光合速率的主要影响因子
处于白桦次生林各大小林隙内和林隙内各位置中的红松最大净光合速率的主要影响因子并不同。其Pmax在林内仅受光饱和点所促进,在小林隙内受光饱和点和气孔导度共同促进,中林隙内受气孔导度促进及光补偿点抑制,但在大林隙内仅受蒸腾促进。其原因则在于林内和小林隙中因周围林冠庇阴强烈,光照较弱(透光率9.2%~27.9%),红松光饱和点低(低于大、中林隙15.1%~30.4%),使其成为Pmax的限制因子。至于中、小林隙内其Pmax受气孔导度促进的原因,则在于中、小林隙内的气温较低(较大林隙低0.9~2.3℃),而低温会限制气孔导度[54],两者的气孔导度较大林隙降低20.1%~43.1%,进而降低了光合反应原料的吸收速率,限制了光合碳同化速率[43-44]。而在大林隙内因周围林冠遮阴影响较弱,光照相对充足(透光率36.9%),故解除了光照限制;同时,光的可利用性增强也会提升蒸腾速率[55],本研究中的大林隙内红松蒸腾速率显著高于小林隙和林内(30.0%~133.3%),这说明光照充足时光合原料的供应及光合产物的运输起着关键作用。
其Pmax在林隙内的边缘区主要受光补偿点抑制,过渡区仅受气孔导度促进,而在中心区受蒸腾速率和总叶绿素含量共同促进。其原因在于各林隙内边缘区因受到周围林冠庇阴强烈,光照最弱(透光率11.4%~11.7%),但其光补偿点却最高(显著高于中心区和过渡区39.3%~163.3%),可能源于林隙边缘区植物争夺光照或维持生存消耗能量过高,而光补偿点高对弱光的利用率较低[56]。由于林隙过渡区气温较低(低于林隙中心区0.4~0.9℃),且低温会限制气孔导度[52],其生长季平均气孔导度显著低于中心区(23.8%~37.6%),降低了光合原料吸收速率,限制光合碳同化速率[43-44]。而在林隙内的中心区光照较强,在光照得到满足的前提下,光合原料供应及产物运输可能也需要同步增强,如其蒸腾速率在林隙中心区显著高于过渡区和边缘区(20.0%~61.5%);但高温和强光也会导致叶绿素的分解[57],进而导致中心区受总叶绿素含量的限制。
4 结 论
温带小兴安岭白桦次生林内红松较大年龄阶段幼树(35 a)的光合适应能力随着林隙增大及沿林隙内边缘区至中心区的微环境梯度均呈现出递增规律性,且大林隙中红松的光能利用效率(光饱和点、表观量子效率)及气孔传输交换能力(蒸腾速率、气孔导度、水分利用效率)均最高,而在林隙内边缘区至中心区光能利用率随微环境梯度呈递增规律,其气体传输与交换能力则为中心区显著高于过渡区与边缘区,这说明次生林下红松幼树不仅在林隙中存在不同适应性,也在林隙内不同位置的微环境也存在相似的适应性差异。红松幼树在长期适应林内光照异质性的条件下,叶片的超微结构同样也存在适应机制,例如形态方面红松幼树比叶面积在大、中林隙中心区显著高于过渡区与边缘区,且红松针叶在林隙和林隙内各位置的叶绿素含量生理适应也均存在差异(在林隙内显著低于林内且在林隙中心区也显著低于边缘区)。因此建议在东北阔叶红松林恢复实践中应创建和利用好较大林隙的中心区(林隙中心区群团式栽植红松或围绕已栽植红松进行透光抚育)的资源优势,充分释放红松生长潜力,方可加速地带性顶极植被阔叶红松林的恢复进程。
