胡蜂毒素Mastoparan的分子改造和定量构效关系研究进展
2022-09-23韩晋辉吕文平
翟 培,韩晋辉,吕文平
(1.广东食品药品职业学院,广州 510520;2.江南大学,无锡 210122)
1979年Hirai等[1]从胡蜂(Vespula lewisii)毒液中分离出一种多肽类物质Mastoparan(MP),因其具有诱导鼠肥大 细胞脱颗粒并释放组胺的功能,所以也被称为“组胺释放因子”。随着研究的深入,人们从其他胡蜂毒液中分离纯化出与MP功能类似的阳离子多肽,序列测定发现它们皆有14个左右氨基酸残基,但氨基酸的组成和序列长度略有不同,故将它们统称为“MP家族”。从数据库UniProtKB(https://www.uniprot.org/)可以检索到32条相关多肽。
MP展示出多种生物活性,作用于不同类型细胞产生效应不同,如:MP作用于肥大细胞可以使其脱颗粒释放组胺,作用于血小板可以使其分泌5-羟色胺,作用于嗜络细胞使其分泌儿茶酚胺类物质,作用于胰岛β细胞使其分泌胰岛素等[2];MP可以活化G蛋白,引发一系列相关的生物效应,如:激活磷脂酶A2、磷脂酶C、磷脂酶D、鸟苷酸环化酶[3-4];此外,MP抗耐药菌和抗肿瘤活性引起研究人员的广泛关注。Das Neves等[5]发现Agelaia-MP(UniProt P69436)和Venom protein 13a(UniProt P84915)可以有效抑制多药耐药鲍氏不动杆菌,并阻止其生物膜的形成;Raynor等[6]研究发现Mastoparan-L(UniProt P01514)、MP-X(UniProt P01515)和MP-J(UniProt P01517)均能抑制白血病HL60细胞的增殖和存活;WU等[7]实验表明,D型MP-M(UniProt P04205)可以有效抑制5种肿瘤细胞的生长,且比L型MP-M具有更高的抗癌活性;Yamada等[8]研究发现MP的类似物能增加线粒体的通透性,影响慢性髓性白血病细胞的生存能力;Annielle等[9]研究发现MP对人多形性胶质母细胞瘤细胞具有明显的促坏死作用;在具有抗癌活性的MP中,Polybia-MP I(UniProt P0C1Q4)对肿瘤细胞具有高度的选择性。它可以选择性地抑制人类白血病T淋巴细胞、前列腺癌和膀胱癌细胞的增殖,对正常(非癌)细胞几乎没有细胞毒性作用[10-11]。由此可以看出,MP在抗多重耐药菌和抗肿瘤方面表现出良好的生物活性,其作用方式不同于传统药物,尤其是在抗肿瘤细胞方面,具有多靶点杀伤作用,作用效应呈多样性,是一种不可多得的天然活性多肽,具有重要的医学价值和良好的应用前景。
1 Mastoparan分子改造研究进展
研究表明,MP的两亲α螺旋结构和阳离子性对其生物活性至关重要,其作用机制为通过静电作用与细胞膜结合,N端位于脂质双层的外侧,酰胺化的C端嵌入脂质双层内部,利用“地毯模型”与膜相互作用,即:MP在膜表面聚集,从而导致细胞逐渐渗漏,其屏障特性短暂中断,细胞内容物发生渗漏,接下来,双层膜以不均匀的方式恢复其连续性,最终肽被夹在两个并列的膜之间,形成高肽脂比的肿块。但MP作用于不同细胞,由于肽的浓度不同、膜的成分不同等因素,其作用机制有所差异,如:Hartman等利用电镜观察MP作用于大肠杆菌时,发现有膜泡及膜皱褶形成;Das Neves等[5]观察MP作用于耐药鲍曼芽胞杆菌时,发现细胞膜有孔洞形成。
MP具有优良的抗菌和抗肿瘤特性,但由其引的炎症反应和细胞毒性等问题极大限制了MP的临床应用。基于MP构效关系的分子改造,可以有效解决上述问题,已成为MP研究的热点。尽管有大量关于优化MP理化参数提高其生物活性的报道,如:阳离子性和疏水性提高有利于MP的抗菌广谱性;降低疏水性可减少MP溶血活性等,但多肽的各种结构参数并非独立发挥作用,一种因素的改变通常会导致其他因素的变化,因此MP的结构改造并没有通用的规则可以使用。目前,国内外对MP结构改造策略主要有两种:(1)通过氨基酸替换、D型改造、序列环化、化学修饰等方法,提高MP生物活性和酶稳定性;(2)通过与其他功能多肽杂合或延伸其末端,提高MP生物利用效率。
1.1 Mastoparan结构特征 MP的抗菌和抗癌细胞活性与其结构特征密切相关。MP结构简单,一级结构由14个左右的氨基酸组成,富含疏水氨基酸和碱性氨基酸,其中疏水氨基酸以亮氨酸、异亮氨酸、缬氨酸和丙氨酸为主,碱性氨基酸以赖氨酸为主,含量一般为2~4个;N端大多由异亮氨酸占据,C末端大部分被亮氨酸占据,且酰胺化;二级结构在水溶液中呈自由卷曲状态,与生物膜或对百日咳毒素敏感性G蛋白的α亚基作用时表现为两亲性的α-螺旋结构[12-16]。MP主要利用静电作用和两亲螺旋结构,通过改变膜的渗透性或直接在膜上形成孔洞来发挥抗菌和抗肿瘤功能[17]。利用多重序列比对软件MAFFT,将UniProtKB数据库检索到的32条MP序列进行比对,其保守序列见图1。
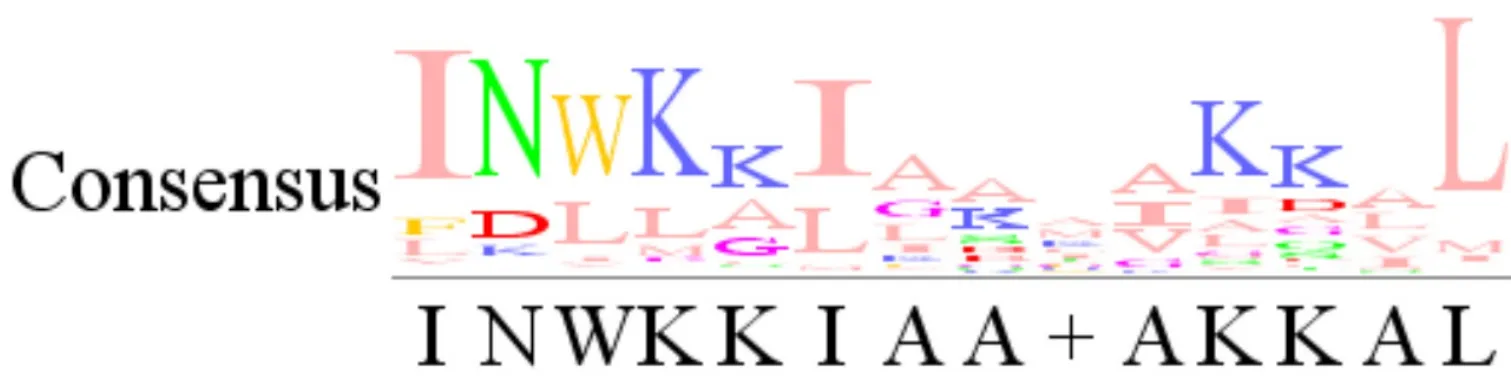
图 1 MP保守序列Fig.1 MP conservative sequence
1.2 Mastoparan的氨基酸替换 John研究团队对MP的构效关系进行了较为系统的研究,他们将MP-L第5位和第8位的丙氨酸用赖氨酸替换,第10位的丙氨酸用α-氨基异丁酸(Aib)替换,目的在于增加亲水面阳离子密度的同时,用α-氨基异丁酸促进螺旋结构的形成,增加肽的酶稳定性。改造后的设计肽被命名为:Mitoparan(MitP),该多肽具有广谱的抗菌特性(S.aureus:MIC 6-8mg/L;E.coli:MIC 4mg/L;C.albicans:MIC 27 mg/L),且同样具有促肥大细胞脱颗粒作用,最重要是MitP穿过细胞膜后在线粒体内聚集,能够促进细胞释放细胞色素C从而引起细胞凋亡[18-19];随后的研究表明,MitP可以在N端连接其他的结构元件:RGD肽、Fas配体模拟物等,携带其一起穿过生物膜,并作用于线粒体[20-21]。John认为MitP可以作为优势骨架(privileged scaffold)对结构进行修饰和改造,以改善生物活性,如:MitP利用丝氨酸替换末端亮氨酸后,由于在疏水面末端引入极性的羟基,使得[Ser14]MitP不仅对真核细胞没有毒性,而且不会引起肥大细胞脱颗粒,但其抗菌活性较MitP有所下降(S.aureus,E.coli,C.albicans,MIC:>32 mg/L),对新型隐球菌C.neoformans保持较高抗菌活性(MIC 5 mg/L)。实验表明[Ser14]MitP可在线粒体内聚集,但不会激活电压依赖性阴离子通道(VDAC),从而可避免细胞凋亡的发生。
Irazzabal等[22]将MP-L第5和第8位丙氨酸分别用异亮氨酸和精氨酸替换,衍生肽命名为[I5,R8]MP、MP-L及[I5,R8]MP螺旋轮图见图2。该多肽具有良好的抑菌活性(S.aureus:MIC 25 μmol/L;E.coli:MIC 12.5 μmol/L;C.albicans:MIC 12.5 μmol/L),体外实验表明[I5,R8]MP对红细胞及HEK-239没有细胞毒性(IC50>200 μmol/L)。该设计将第8位换为精氨酸后,MP净电荷从+3增加到+4,且根据Pace-Scholtz's α螺旋倾向表[23],精氨酸更倾向于形成α螺旋结构;氨基酸替换后未改变多肽整体极面角度Φ和疏水力矩(hydrophobic moment; Ile和Arg疏水指数:4.5和-4.5)。
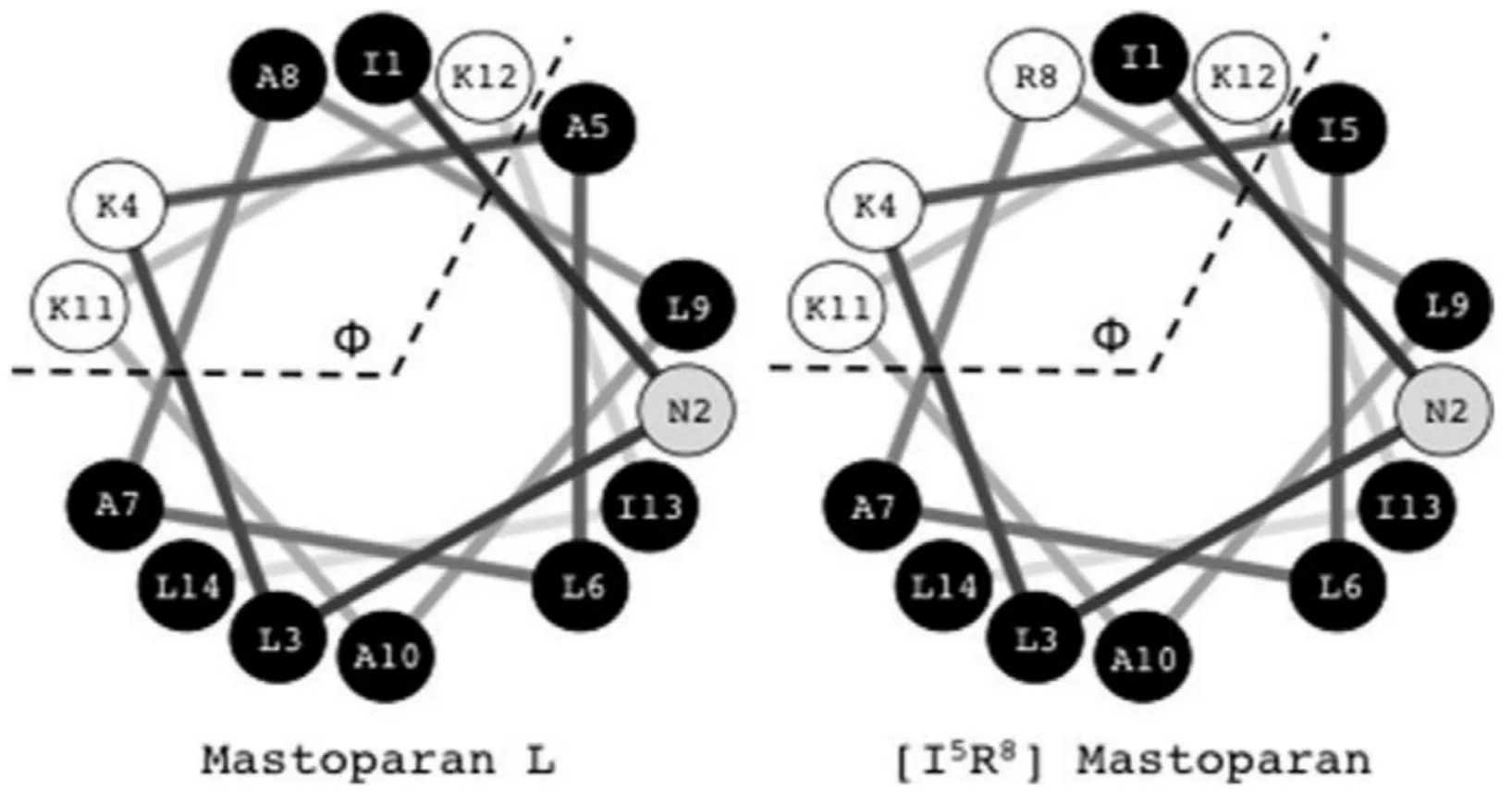
图2 MP-L及[I5,R8]MP螺旋轮图Fig.2 Helical wheel representation of MP-L and [I5,R8]MP
1.3 Mastoparan的D型及全反改造 某种多肽如果通过与特定受体或酶结合发挥其生物功能,那么D型多肽由于侧链方向的改变将无法发挥其原有生物功能;当D型多肽保持生物活性时,说明多肽可能是与磷脂相发生作用,而没有特定的受体。因此多肽D型及全反设计是研究结构与活性关系的常用手段,也是增加多肽酶稳定性的有效方法[24]。Sarah等[25]将MP-L和MitP进行了D型改造及逆向改造,序列见表1。实验表明MP-L及MitP的D型异构体保持了其对肿瘤细胞的毒性、肥大细胞脱颗粒活性及膜活性,但逆向构型和全反构型却几乎没有活性,说明MP-L及MitP的生物功能与结构密切相关,但并非通过特定受体或酶发挥作用。

表1 MP、MitP 及其异构体氨基酸序列及分子量Table 1 Amino acid sequence and molecular weight of MP, MitP and their isomers
该实验还发现D型MP-L有潜力成为抗酶解穿膜肽(CCP)。D型MP-L在5 μmol/L浓度下,与穿膜肽Penetratin和TAT相比,D型MP-L可以更快、更有效的进入U373MG细胞,主要聚集在中性的溶酶体中。
1.4 Mastoparan的环化改造 Chen等[26]对Mastoparan C(LNLKALLAVAKKIL-NH2)进行了改造研究,在MP-C末端添加二硫键环化,设计出环形MP类似物cMP-C(CLNLKALLAVAKKILC-NH2)。经过环化之后的MP-C在血清中的稳定性增加,但其抗菌活性较母肽明显下降,且溶血活性也有较大的增加;值得注意的是该实验结果显示α螺旋含量最高的cMP-C最快观察到渗漏现象,但其抑菌活性却不及其他两个MP类似物,其原因可能是MP-C经环化后,疏水氨基酸更加集中,导致溶血性增加,但末端增加2个半胱氨酸之后,反而因为增加氨基酸使得阳离子变的分散,不利于抑菌活性。cMP-C对哺乳动物细胞的毒性和膜渗透能力显著大于母肽MP-C。由此可见,α螺旋结构对膜渗透有直接影响,但并不是多肽生物活性的唯一决定因素。
1.5 Mastoparan的杂合改造 宋竟婧等[27]以MitP为分子骨架结构,设计出MitP的阴离子伙伴肽MitPE(INLEHLAHL(Aib)EEIL),并将MitP与MitPE通过二硫键连接起来,设计出具有非常优异的抗肿瘤特性的AMitP,其分子结构见图3。在正常酸碱度下,MitP肽段与MitPE肽段之间由于静电作用及分子间作用的影响,使得AMitP失去了生物活性;在酸性条件下,MitP与MitPE肽段之间的静电作用减小,AMitP恢复活性构象。酸度依赖特性使该设计肽具有细胞选择性:对于正常细胞,其胞外环境为中性,因此AMitP对正常细胞几乎没有伤害;对于肿瘤细胞,因其胞外环境为酸性,AMitP可以选择性的通过静电作用与肿瘤细胞结合,并通过两亲结构与肿瘤细胞膜作用,从而杀死肿瘤细胞。

图3 酸活化抗菌肽AMitP分子结构Fig.3 Molecular structure of acid-activated antimicrobial peptide AMitP
Chen等[26]在MP-C的N末端添加细胞穿透肽TAT(RKKRRQRRR),设计出MP类似物tMP-C(RKKRRQRRRLNLKALLAVAKKIL-NH2)。tMP-C与母肽相似,保持了较高的抑菌活性,且具有更强的抗肿瘤细胞活性(对5种测试肿瘤细胞的IC50小于4 μmol/L),但tMP-C对正常细胞HMEC-1的IC50<10 μmol/L。tMP-C抗肿瘤活性是母肽的5倍,但其膜渗透作用大大降低,推测tMP-C抗肿瘤活性可能与线粒体膜的作用有关。
Transportan(TP)由一个赖氨酸残基将甘丙肽(galanin)N端的12位氨基酸与MP相连构成。TP能跨膜转运生物大分子物质如绿色荧光蛋白,它具有富集于细胞膜附近、高效携带大分子进入细胞的能力[28]。TP包含了甘丙肽N端的活性片段,可被甘丙肽受体识别。此外,研究显示,TP对人黑色素瘤细胞膜上的基底GTP酶有抑制作用,虽然产生这种抑制作用所需的TP浓度通常高于转运物质所需的浓度,但这个特性仍是载体肽的重要瑕疵。为了减小TP与甘丙肽受体的亲和力以及它与G蛋白的相互作用。Soomets等[29]合成了TP的9条类似物,研究发现,类似物TP10即使在高浓度下也不会影响基底GTP酶活性,也不会被甘丙肽受体识别,这些特性使得TP10有望成为新的、副作用显著减小的载体肽。TP10含有21个氨基酸残基,由1个赖氨酸将长度为6个氨基酸的甘丙肽片段和14个氨基酸的MP连接而成。与母肽TP相比,TP10的穿膜能力更强,并且不会抑制RIN-m5F细胞膜基底GTP酶活性,毒性更低。近年来有研究报道,TP10具有较好的细胞选择性,能优先进入微生物细胞膜并杀死细菌(S.aureus:MIC 7.33 μmol/L,E.coli:MIC 7.33 μmol/L,C.albicans:MIC 7.33 μmol/L)及耐药菌(A.baumannii:MIC 3.67 μmol/L),却几乎不会损伤哺乳动物细胞[30]。这些研究显示MP杂合肽TP10具有发展为抗微生物尤其是抗多重耐药细菌的新型抗菌物质的潜力。Rusiecka等[31]发现TP10对人类宫颈癌细胞和人类骨肉瘤Os143b细胞株具有体外抗癌活性。
2 Mastoparan的定量构效关系研究
多肽的分子结构与生物活性密切相关,因此,结构参数是生物活性改良研究中的重要参考指标。定量构效关系QSAR(quantitative structure activity relationship)是应用数学统计学方法来定量研究分子结构与特定的生物活性之间的关系。它是多肽设计的一种有力工具,也是在缺乏准确作用机制情况下,有效避免盲目改造的合理方法。良好的QSAR模型生成取决于训练集和测试集在结构多样性和属性值分布方面的质量。由于MP具有卓越的药理作用、相对简单的结构,已经成为QSAR研究的一种模型肽。
基于Čeřovský等[32]测得的40条MP及其衍生物对枯草芽孢杆菌Bacillus subtillis的最小抑菌浓度(minimal inhibitory concentration, MIC)数据,早在2011年Avram等采用多元线性回归分析(multiple linear regression analysis, MLRA)对MP进行了二维定量构效关系(2D-QSAR)研究,并建立了MP结构特征(理化参数)与抑制枯草芽孢杆菌MIC值的相关模型[33],其中预测效果较好的5条回归方程见表2,该模型对MP衍生物的预测相关系数为0.81~0.83。

表2 MP二维定量构效关系模型Table 2 Two dimensional QSAR model of MP
随着3D-QSAR技术在生物学上的成功应用,2012年Avram等[34]在前期研究基础上,利用比较分子力场分析法(comparative molecular field analysis, CoMFA)和比较分子相似性指数分析法(comparative molecular similarity in-dices analysis,CoMSIA)重新对MP构建模型,用以评估氢键供体/受体、疏水性、空间性和静电性在MP对枯草杆菌抗菌活性中的贡献值,由于该方法对分子的排列很敏感,作者考虑了两种叠加方式:Cα原子的叠加(QSAR模型1)和骨架原子的叠加(QSAR模型2),从而将肽叠加在最有效的结构上。该研究获得良好的统计参数,从结果来看,用较少原子结构对准的方法(模型1),预测结果更加有效,非常接近实验值(pMIC值偏差在0.7以内)。
2015年,Mariya等[35]把氨基酸作为多肽的基本单位,即:端点=功能,采用蒙特卡洛方法计算MP的QSAR模型,利用3种模型预测33种MP衍生物对枯草芽孢杆菌的抑菌活性,预测结果优于2D-QSAR模型。
QSAR模型的预测精度取决于样本的数量、训练集和测试集的恰当选择以及选择最相关的分子描述符和合适的统计方法等,其中选择最合适的分子描述符是获得准确QSAR模型的先决条件,但有效分子描述符的选择依赖于问题,并没有通用规则。本文提到的三种建模方式各有优缺点,多肽的各种结构参数并非独立发挥作用,而是密切相关的,一种因素的改变通常会导致其他因素的变化。因此,在多肽设计时,应以构效关系为基础,综合权衡各项结构参数,以期获得生物活性最优良的多肽。
3 总结与展望
胡蜂毒素的C端非常保守,通常由疏水的亮氨酸占据,研究人员将MP衍生物C端的亮氨酸用丝氨酸替换后,将不会引起肥大细胞脱颗粒;本实验室将胡蜂毒素与天蚕素A进行杂合设计,设计杂合肽的抗细菌活性比MP-L提高4~8倍,且溶血活性大大降低。由以上可知,对胡蜂毒素保守序列的合理修改或重新设计将极大改善其活性,有助于胡蜂毒素在微生物防治和药物开发的利用。随着胡蜂毒素相关研究的深入,人们对其结构特征、生理作用及生物活性有了新的认识,在此基础上进行的分子改造目标也更加明确。一方面,通过分子改造探索其在抗肿瘤和抗耐药菌方面的潜力;另一方面,胡蜂毒素具有惰性穿膜肽的特质,通过分子改造,可以丰富穿膜肽的种类,并作为一类先导化合物用于开发安全有效的多肽药物。在过去的十多年里,天然的、改良的或杂合的MP已经被用作许多神经疾病的潜在药物进行研究[36]。目前MP分子改造的方法和理念已经发生转变,更趋向于通过结构活性关系研究,利用计算机辅助的理性或半理性设计,这将大大提高分子改造的效率。
蜂毒用于治疗疾病已有几百年的历史,可有效地治疗多种疾病。我国拥有非常丰富的蜂毒资源,但对于蜂毒多肽的药理以及临床应用的研究、开发和利用却还不够深入。以胡蜂毒素Mastoparan为代表的蜂毒多肽具有广泛的药理作用以及临床应用价值,应当加速开展蜂毒多肽在临床医学、生物工程以及其他领域的各项研究。
