不同冬小麦品种(系)拔节期低温胁迫生理反应及抗寒性评价
2022-09-23曹燕燕葛昌斌齐双丽廖平安李雷雷乔冀良张振永
曹燕燕, 葛昌斌, 齐双丽, 廖平安, 黄 杰, 王 君, 李雷雷, 乔冀良, 张振永
(漯河市农业科学院/河南省抗赤霉优质小麦种质创制及新品种培育工程技术研究中心,河南漯河 462300)
小麦是我国主要粮食作物,小麦的可持续发展关系到我国的粮食安全、人民温饱和社会稳定。近几年,随着极端天气频繁出现,低温冻害已成为影响小麦生长发育的重要环境因素之一。特别是春季低温,小麦正处于拔节至孕穗期,此时小麦组织器官幼嫩、含水量大,抵抗霜冻的能力较弱;同时小麦穗分化正处于雌雄蕊分化至小花原基分化期,对低温反应尤其敏感,如遇低温很容易造成伤害。黄淮麦区是我国冬小麦主产区之一,目前低温冷害已成为该区主要气象灾害之一,发生频率高、面积大、危害重,严重制约小麦生长,因此对黄淮麦区的小麦品种进行抗寒性鉴定研究具有重大意义,不仅关系到今后小麦的育种和推广问题,也直接关系到国家的粮食安全和社会稳定等问题。
小麦遭受低温胁迫后,植株会产生一种应激反应起到保护作用,主要表现在小麦体内可溶性糖、可溶性蛋白、游离脯氨酸等渗透调节物质的含量明显增加,细胞膜结构与成分改变以及一些新酶的产生等。于晶等研究表明,抗寒性强的小麦品种脯氨酸和可溶性蛋白含量均高于抗寒性弱的品种;刘艳阳等研究表明,超氧化物歧化酶(SOD)活性、丙二醛(MDA)含量是小麦抗寒性的2个抗寒指标;余海波研究发现,小麦拔节期遭受低温胁迫后,对SOD和过氧化物酶(POD)活性、脯氨酸和可溶性蛋白质含量、可溶性糖含量、MDA含量等都有影响;卫秀英等的研究发现,抗寒性弱的小麦品种MDA含量高于抗寒性强的品种,而SOD活性和POD活性均低于抗寒性强的品种。在生产中,单一的生理指标很难准确反映各个小麦品种抗寒性的强弱,因此可结合隶属函数法和产量指标等进行综合评价。
漯麦906于2019年通过河南省审定,审定编号为豫审麦20190064;漯麦47于2021年通过湖北省审定,审定编号为鄂审麦20210007;漯麦36已完成试验程序,即将国审。本研究以小麦新品种漯麦47、漯麦906、对照周麦18、参试新品系漯麦36、漯麦6098、漯麦956和漯麦C101共7个试验材料为研究对象,室外盆栽种植,至小麦拔节期连盆一起移入春化室模拟低温处理,探索拔节期低温对不同品种(系)小麦叶片生理生化特性的影响,并运用隶属函数对各品种(系)抗寒性进行分析,结合产量表现对供试材料的抗寒性做出综合评价,以期为小麦品种漯麦906和漯麦47,将审定品系漯麦36做出抗寒性评价以及后期示范推广和育种亲本的选择等提供技术支撑。
1 材料与方法
1.1 试验材料与设计
试验于2020—2021年在河南省漯河市农业科学院试验田网室进行,采用盆栽方法。供试材料为漯麦47、漯麦906、周麦18、漯麦36、漯麦6098、漯麦956和漯麦C101共7个品种(系)小麦,种子由漯河市农业科学院小麦研究所提供。盆栽用土为大田 0~30 cm耕层土,有机质含量为14.13 g/kg,全氮含量为0.86 g/kg,硝态氮含量为25.56 mg/kg,速效磷含量为42.59 mg/kg,速效钾含量为168.99 mg/kg,pH值为7.47。盆直径30 cm,高60 cm,底部用网袋包裹,埋入大田,盆内土壤表面与地面持平。播前每盆施复合基肥6 g,土壤水浇透沉实后于2020年10月15日播种。3叶期定苗,每盆留苗9株,每个品种种植9盆,其中3盆用于低温取样,3盆用于低温胁迫后测产,3盆用于常温对照,共计63盆。浇水量根据土壤墒情决定,每盆浇水量一致。其他管理同大田,2021年6月2日收获。
1.2 低温处理方法
2021年春季,各小麦品种生长至拔节期,通过显微镜观察幼穂分化,各品种进入雌雄蕊原基分化期。将各品种6盆从大田移入春化室模拟春季低温胁迫,春化室内温度设置为白天0 ℃、夜间-4 ℃,温度误差为0.5 ℃,空气相对湿度为70%。采用人工光源光照,光照时间为08:00—18:00。低温处理0、1、2、3、4 d时取主茎最上部的展开叶用于生理指标检测,每个品种取3盆,3个重复。各个品种剩余3盆在低温处理后移至自然条件下,直至收获,用于产量性状方面的调查。
1.3 测定项目及方法
生理生化性状的测定:每个供试材料测定3次取其平均值。采用硫代巴比妥酸比色法测定MDA含量;采用氮蓝四唑(NBT)法测定SOD活性;采用愈创木酚法测定POD活性;采用过氧化氢法测定过氧化氢酶(CAT)活性;采用考马斯亮蓝G-250比色法测定可溶性蛋白质含量;采用酸性茚三酮法测定脯氨酸含量。
产量因子的测定:低温胁迫处理4 d后,将未取样的盆栽小麦放置到大田原位置,自然条件下自然恢复生长。蜡熟期后收获,分别记载单株穗数、单株穗粒数和单株产量。
1.4 抗寒性评价及方法
应用隶属函数法评价抗寒性,计算公式如下:
(1)与抗寒性呈正相关的各指标(SOD活性、POD活性、可溶性蛋白含量和脯氨酸含量等)用公式()=(-)/(-)计算。
(2)与抗寒性呈负相关的各指标(MDA含量) 用公式()=1-(-)/(-)计算。式中:()为第个品种、第天相对百分数的隶属度;表示第个品种、第天某一指标测定值与其对应对照值的百分数;、为所有供试品种某一具体指标在某一天最大值和最小值的相对百分数值。对各个指标所求的隶属函数值进行累加、求取平均值。之后按照从大到小的顺序对各个品种的隶属函数平均值进行排序,对应的得到各个品种抗寒性的强弱。
1.5 数据处理
利用WPS Office进行数据统计和作图,利用DPS 15.10的Duncan’s新复极差法进行差异显著性分析。
2 结果与分析
2.1 低温胁迫对拔节期小麦叶片SOD活性的影响
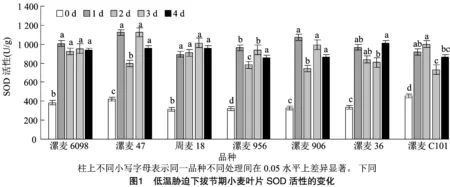
如图1所示,拔节期持续低温胁迫后,各小麦品种(系)SOD活性出现不同程度增加,以增强对逆境的适应能力,与常温对照相比,均差异显著。随着低温持续时间的延长,SOD活性整体表现为先升后降的趋势。低温处理1 d后,品种(系)间升高幅度存在明显差异,漯麦906升幅最大(227.42%),漯麦C101升幅最小(101.09%),其余品种(系)升幅介于两者之间;低温处理2 d,除周麦18和漯麦C101外,其他品种(系)SOD活性出现下降;随着低温处理时间延长至3 d和4 d,大部分品种(系)SOD活性表现先上升后下降。由试验可知,在持续低温胁迫下,不同小麦品种(系)SOD活性存在差异,但都有较大幅度提高,以抵御低温胁迫对植株造成的伤害。
2.2 低温胁迫对拔节期小麦叶片POD活性的影响
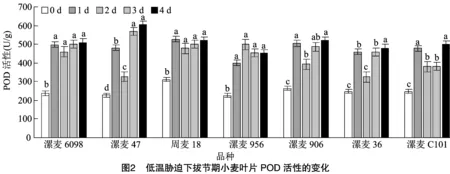
如图2所示,低温胁迫后各品种(系)的POD活性均有升高,且升高程度不同,也都与对照差异显著(除漯麦36处理2 d与对照差异不显著外)。低温胁迫1 d后,各品种(系)之间以漯麦47升幅最高(111.29%),其次为漯麦6098(109.74%);低温胁迫2 d,除漯麦956活性继续增加外,其余6个品种(系)POD活性均下降;低温胁迫3 d和4 d,各品种(系)POD活性持续上升,与对照相比,增幅为68.33%~167.19%,其中漯麦47和漯麦6098增幅最大,分别为167.19%和115.24%。说明在长期的低温胁迫下,漯麦47和漯麦6098的POD的抗氧化能力仍持续较高,而其他5个品种(品系)的抗氧化能力稍弱,但对活性氧自由基仍有一定的清除作用。
2.3 低温胁迫对拔节期小麦叶片CAT活性的影响
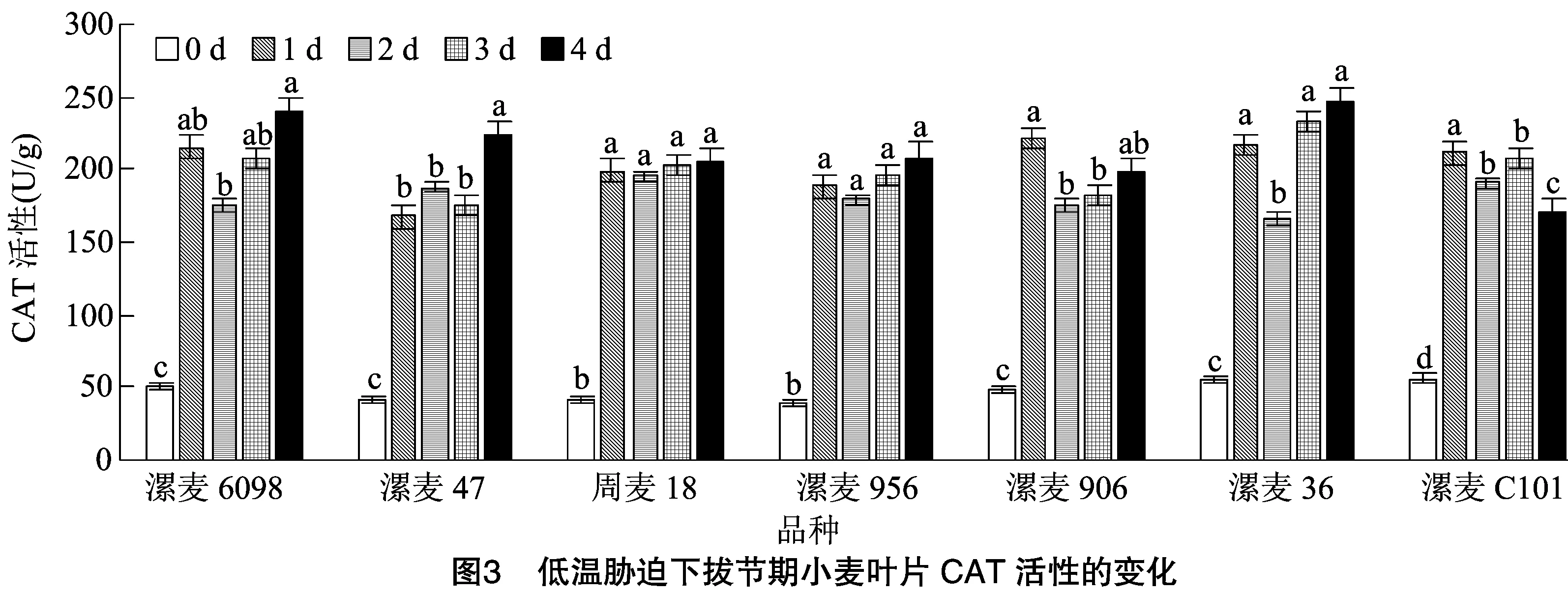
如图3所示,低温胁迫后各品种(系)的CAT活性均有增加,也都与对照差异显著。低温胁迫处理1 d后,7个小麦叶片中的CAT活性平均升高328.74%,但品种(系)间升高幅度存在明显差异;除漯麦956和漯麦C101外,随着低温胁迫时间的延长,各品种(系)CAT活性呈现“升高—降低—升高—升高”的趋势;低温处理 4 d 后,与对照相比,漯麦47增幅最高,为444.89%,漯麦C101增幅最少,为203.37%,其他材料介于两者之间。这表明持续低温胁迫后,各供试材料清除活性氧的能力有差异,但在低温胁迫4 d后,除漯麦906和漯麦C101外,其他品种(系)CAT活性仍较高,可以将低温对植物的伤害控制在一定范围内。
2.4 低温胁迫对拔节期小麦叶片丙二醛活性的影响
如图4所示,低温胁迫后各品种(系)的MDA含量均有增加,也都与对照差异显著。低温胁迫 2 d,漯麦47和漯麦C101的MDA含量下降,其他5个材料MDA含量略有上升;与处理3 d相比,低温胁迫4 d后漯麦6098、漯麦47、周麦18和漯麦906的MDA含量出现下降,其他3个材料MDA含量则继续上升。这说明低温加速了各品种(系)膜脂过氧化,活性氧已伤害到各品种(系),但根据MDA含量的不同,各品种(系)间受伤害的程度存在差异。

2.5 低温胁迫对拔节期小麦叶片脯氨酸含量的影响
由图5可以看出,低温胁迫后各品种(系)的脯氨酸含量均有增加,也都与对照差异显著。随着低温胁迫时间延长,漯麦47、周麦18、漯麦906和漯麦36的脯氨酸含量一直呈现上升趋势;与胁迫1 d相比,漯麦6098、 漯麦956和漯麦C101的脯氨酸含量呈现先上升后降低再上升的趋势。由此可以看出,低温胁迫下,各品种(系)会积累较多的脯氨酸,以维护细胞膜内代谢的平衡和细胞膜结构的稳定性,以减轻低温引起的伤害,但各品种(系)间脯氨酸含量变化趋势存在差异。

2.6 低温胁迫对拔节期小麦叶片可溶性蛋白质含量的影响
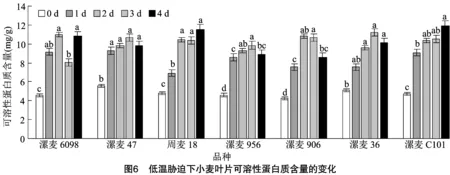
如图6所示,各供试品种(系)在拔节期遭受低温胁迫后,可溶性蛋白质含量均有不同程度增加,除漯麦36外,其他6个均与对照呈显著性差异。低温胁迫1 d后,各品种(系)之间以漯麦6098升幅最高(100.07%),周麦18升幅最低(43.98%),其他供试材料介于两者之间;与低温胁迫2 d相比,低温胁迫 3 d,漯麦6098、周麦18和漯麦906可溶性蛋白质含量出现下降,其他4个持续上升;与低温胁迫3 d相比,低温持续胁迫4 d,漯麦6098、周麦18和漯麦C101可溶性蛋白质含量出现上升,其他4个品种(系)出现下降,漯麦6098表现显著差异。由此可知,低温胁迫时间越长,不同品种(系)间可溶性蛋白含量差异越大。
2.7 拔节期低温胁迫对小麦籽粒产量构成因素的影响
由表1可知,与CK相比,小麦在拔节期受到低温胁迫4 d后,各供试材料不孕小穗数显著升高,单株分蘖数、穗粒数、结实小穗数、千粒质量和单穗产量均减少,不孕小穗数增加。其中漯麦47单株减产率最低,为39.57%,漯麦956减产率最高,达到75.73%。综合产量各因素变化数据,在本试验环境下,拔节期低温胁迫对小麦不孕小穗数和单穗产量影响较大,而对结实小穗数和穗粒数影响比较小。
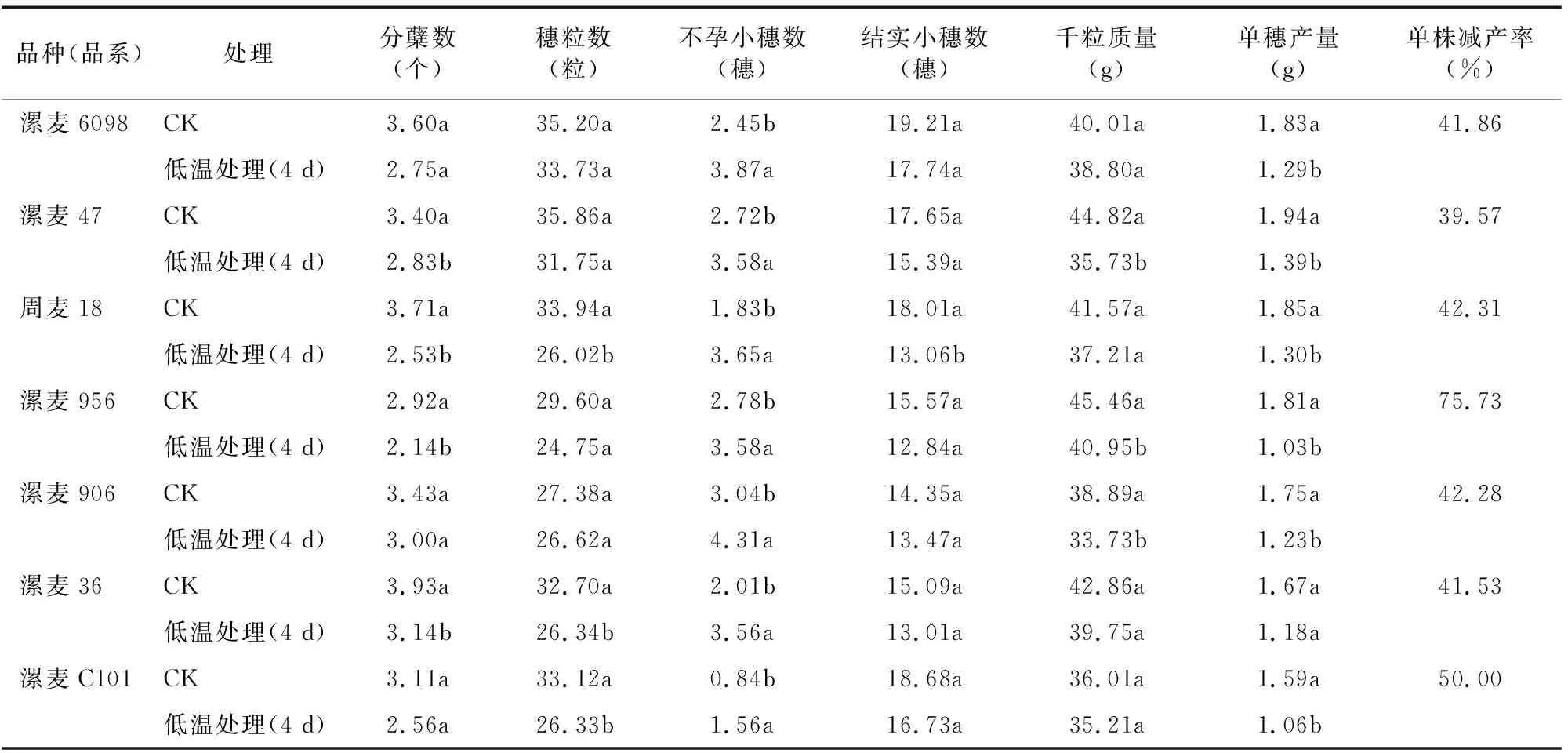
表1 拔节期低温胁迫对单株小麦产量的影响
2.8 小麦品种抗寒性的隶属函数值综合分析
从表2可知,在低温胁迫下,品种间隶属函数值差异非常明显,同一品种(系)不同指标的隶属函数值也存在差异。每一品种(系)各项测定指标的隶属值的平均综合值越高,表明该品种(系)的抗寒性越强。根据综合值,将7个供试品种(系)可分为3类:第Ⅰ类为漯麦6098、漯麦47、周麦18和漯麦906,属于抗寒性强的品种(系);第Ⅱ类为漯麦36和漯麦C101,属抗寒性较好的品系;第Ⅲ类为漯麦956,属抗寒性弱的品系。试验结果与各品种(系)的单株减产率基本一致,且与田间各品种(系)的抗寒性表现基本一致。

表2 低温胁迫下小麦叶片各生理指标的隶属值
3 讨论与结论

持续低温胁迫下,当保护酶的清除能力不能及时清除多余的氧自由基时,就会引起膜脂不饱和脂肪酸发生过氧化,产生膜脂过氧化产物MDA,膜脂过氧化的程度随温度降低加重,同时与低温胁迫时间呈正相关。本试验中,低温胁迫1 d后,各供试材料的MDA含量相比于对照,均显著上升,表明低温胁迫后,各供试材料抗氧化能力虽有所增强,但仍不能及时清除活性氧,引起膜脂过氧化,对小麦造成伤害;随着低温胁迫的持续,各供试材料MDA含量变化有升有降,说明各个品种(系)对低温的适应调节能力存在差异;低温胁迫4 d与胁迫 3 d 相比,抗寒性强的4个品种(系)漯麦6098、漯麦47、周麦18和漯麦906,MDA含量出现下降,说明抗寒性强的品种,在持续低温胁迫后,能够通过自身调节使质膜透性部分修复。
脯氨酸通过其亚氨基促进蛋白质水合而成为重要的渗透调节物质之一,可溶性蛋白的增多有利于维持低温状态时酶的构象,因此脯氨酸是植物抗寒生理中的一个重要指标。本试验中,在低温胁迫1 d后,各供试品种(系)脯氨酸含量均有不同程度增加,均与对照呈显著性差异。随着低温胁迫时间延长,漯麦47、周麦18、漯麦906和漯麦36的脯氨酸含量一直呈现上升趋势;与胁迫处理1 d相比,漯麦6098、漯麦956和漯麦C101的脯氨酸含量呈现先上升后降低再上升的趋势。说明漯麦47、周麦18、漯麦906和漯麦36这4个品种(系)在持续低温胁迫下可以通过积累较多的脯氨酸来提高自身抗寒性。
可溶性蛋白作为渗透调节物质广泛存在于植物细胞内,作用主要是为了调节细胞内渗透压的平衡、维持细胞正常的生理代谢活动。研究表明,植株细胞内可溶性蛋白含量的高低与其自身抗寒性存在密切的关系。低温胁迫后植株体内可溶性蛋白含量升高,以提高植物在低温下的渗透调节能力和代谢功能,进一步提高植株的抗寒能力。本试验中低温持续胁迫4 d后,漯麦6098、周麦18和漯麦C101可溶性蛋白质含量出现上升,与胁迫处理3 d相比,漯麦6098表现显著性差异。说明这3个供试材料在长期低温胁迫下,通过提高体内可溶性蛋白的含量来增强自身的抗寒能力,漯麦6098表现更突出。随着低温胁迫时间的延长,其他试验材料可溶性蛋白含量存在差异。
拔节期低温冻害对小麦穗部发育和单株产量影响较大,不孕小穗数显著提高,穗粒数减少,千粒质量降低,且各材料间单穗产量下降幅度不同。本试验中漯麦47单株减产率最低,为39.57%,漯麦956减产率最高,达到75.73%。综合各产量因素变化数据得出,拔节期低温胁迫对小麦单株产量和不孕小穗数影响较大,而对结实小穗数和穗粒数影响相对较小。
通过隶属函数综合值分析评价小麦拔节期的抗寒性,供试材料被分为3类。漯麦6098、漯麦47、周麦18和漯麦906,属于抗寒性强的品种(系);漯麦36和漯麦C101,属抗寒性较好品系;漯麦956,属抗寒性弱的品系。这是检测各材料几个生理生化指标综合反应的结果,能够较准确地反映出各品种(系)间抗寒性的差异,再结合各材料的产量指标,来综合评价各供试材料的抗寒性,更加准确。
拔节期是小麦营养生长的重要时期,也是对低温敏感的时期。低温胁迫下,植物细胞多种生理生化指标发生改变以适应低温条件和进行自我调节。小麦应对低温逆境胁迫涉及一系列复杂因素,既有外部因素,也有内部因素。本研究根据试验需要,筛选了7个供试材料,研究了拔节期持续低温胁迫下生理生化指标和产量的变化,为收集和鉴定抗寒性小麦种质资源提供依据。目前分子生物学技术发展迅猛,植物耐受低温胁迫机制的研究也从生理生化水平向分子水平转变,下一步将开展相关抗冻基因表达等方面的研究,将分子育种手段有效融合到常规育种中,为培育抗寒、耐倒春寒的小麦新品种提供技术支撑。
