不同生境赤皮青冈种子萌发及幼苗高的生长规律研究
2022-09-22陈国兴
陈国兴
福建省建瓯市林业局,福建 建瓯 353100
种子是木本植物在生活史上的重要阶段,也是其在空间上逃避不利环境,并且确保其成功定居及更新的保障[1]。种子的萌发及幼苗的生长是种群更新过程中重要环节,且木本植物实生苗的更新在植物群落的演替、植被的恢复及其生物多样性的维持等都具有重要作用[2]。木本植物种子的萌发特性和自然分布区的气候及其生境条件密切相关,是木本植物对自然分布区生境长期适应的结果[3]。因此种子的萌发及幼苗的生长受多种因素的影响,它不仅能反映植物对生境资源的竞争能力以及利用效率,还能影响种群的生存与发展及森林的发育与更新[4]。赤皮青冈(Cyclobalanopsis gilva)属于壳斗科(Fagaceae)青冈属(Cyclobalanopsis),常绿乔木,主要分布于浙江、贵州、福建等地,在分布区内是主要的建群种之一[5],其生长速度较快,适应性强,其木材可用于制作工艺品、上等家具及运动器材,果实能制作粉条等美食。但由于长期不规范的管理及过度砍伐,赤皮青冈种群缩小,林分更新十分困难,有濒危的趋势[6]。以往对赤皮青冈的研究主要集中在群落学[7]和生理特性[8]等方面,对种子的研究主要集中在对外源激素的处理[9]和组织培养技术[10]。有关赤皮青冈不同生境的实生苗种子萌发及生长发育特点的研究还未见报道,而这却是赤皮青冈天然林植被恢复和可持续经营管理中亟需解决的问题。本文研究不同生境(苗圃地和野外山地)下种子萌发和幼苗生长特征,以Logistic曲线方程为模型,对不同生境的赤皮青冈幼苗苗高进行拟合分析,旨在建立赤皮青冈苗期的生长模型,分析其生长参数,从而掌握其生长规律,为赤皮青冈野生资源保护和持续经营及科学的开发利用提供理论依据。
1 材料与方法
1.1 种子处理
在福建省建瓯市擎天岩的赤皮青冈林中,拾取从母树上自然掉落的果实,并选择无病虫害、完整且颗粒饱满的赤皮青冈果实进行分层贮藏在阴凉的地下室湿沙中;为保持与野外的条件一致,贮藏的果实不进行浇水、杀虫、除菌等处理。3个月后,将贮藏的果实取出泡入水中,选取沉入水底的赤皮青冈果实在室内晾到3天后在苗床和野外播种。
1.2 播种
3月16日进行播种,在福建省建瓯市卓坑苗圃的播种方法采用条播的方式,行距为20 cm,每行播种约20粒种子,覆土厚2~3 cm;野外采用点播的方式在赤皮青冈母树林、赤皮青冈与毛竹混交林、赤皮青冈林缘(与锥栗林相邻)、近山顶裂斗锥树下、近山顶林缘设置样方,为使数据完整及消除不同立体条件的影响分别设置6、6、3、2、1个1 m × 1 m的小样方进行野外种子萌发试验,数量为50粒·m-2,覆土厚2~3 cm,共18个样地,依次进行编号。不同生境的生态因子特征见表1。
1.3 调查
从3月16日到12月8日对野外和苗圃分别进行跟踪观察和记录。当赤皮青冈出苗整齐后进行编号,选定25株标准株,在5月27日开始生长量测量,每10 d进行一次苗高测定。
1.4 数据处理
利用SPSS软件和Excel 对数据进行整理及分析比较。在一定时期内,植物的生长过程可通过Logistic模型进行定量描述[11],且非线性Logistic模型更易拟合植物的生长曲线,方法简易[12-13]。Logistic曲线的拟合方法众多,本文采用三点法进行估计值,再根据对数线性化的方法求解参数和。模拟动植物生长过程的数学表达式为:

式中:y表示观测植物指标的累积生长量;K表示植物的终极生长量;e表示自然对数的底;a、b表示待定的常量;t表示植物的生长时间。按照微分方程理论估计值,为:
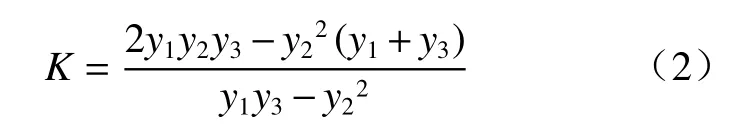
式中,y1、y2、y3分别表示t1、t2、t3时刻植物累积的生长量,并要求t2-t1=t3-t2。由式(1),求出a、b的参数值。张连翔[14]等人估算出植物生长速生期的始点、中点及终点值,并估出速生期的长度,公式为:
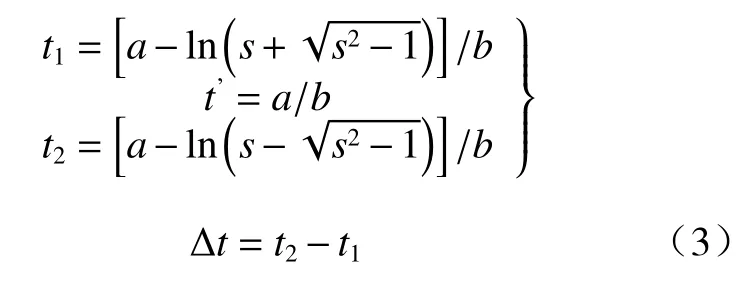
式中:t1、t’、t2、Δt分别表示速生期的始点、中点、终点及长度;s=[(a+1)+ea(a-1)]/(ea-1),其中a、b表示Logistic曲线的参数值。将t1和t2代入式(1)中,得出植物速生期始点(y1)和终点的累积生长量(y2)。则植物速生期内的累积生长量为:Δy=y2-y1,依此,估算出植物速生期始点(P1)和终点(P2)及速生期内的累积生长百分率(ΔP),公式为:
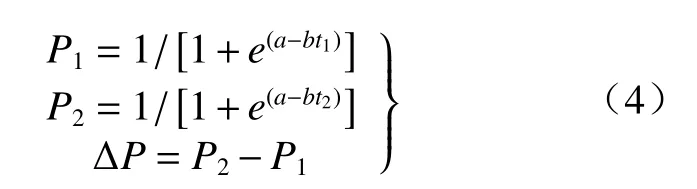
到后慢期内的累积生长量(y3)与后慢期内的累积生长百分率(P3)的公式为:y3=K-K2;P3=1-P2。在进行实际应用时,应注意先设定调查日期的起始日为“0”,将其转化为具体时间(d),方能拟合Logistic曲线。最后由式(3)求出t1、t’和t2,t1和t2可将植物的生长划分为生长前期、速生期及生长后期。
2 结果与分析
2.1 不同生境对种子萌发的影响
通过一年的跟踪观察(见图1),苗圃地5月27日发现约10%的种子幼苗出土,而野外5个生境的出土时间较晚,约在7月12左右;不同生境的赤皮青冈种子萌发的幼苗出土保存率总体均呈现出“慢—快—慢”的趋势。苗圃地和近山顶林缘在同样光照条件下,苗圃地的种子幼苗出土时间早于近山顶林缘,说明在海拔低的地方种子幼苗出土时间较早,海拔对种子萌发的幼苗出土时间影响较大。

表1 不同生境赤皮青冈生态因子特征Tab.1 Characteristics of ecological factors of Cyclobalanopsis gilva in different habitats
由图1可知,赤皮青冈母树林和与毛竹的混交林幼苗出土保存率高于其他生境,分别为25.67%和25%;近山顶林缘的幼苗出土保存率为18%;近山顶裂斗锥树下的幼苗出土保存率最低,出土总数量仅有2粒,幼苗成长期间1株枯死;赤皮青冈林缘的出土总数量为43粒,其中一半的幼苗枯死,保存率为14%,幼苗死亡率较高。幼苗出土率见表2,结果表明母树林内的幼苗出土率最高,为31.67%,其次是毛竹林,为31.00%,近山顶裂斗锥树下的幼苗出土率最低,仅为2.00%,但都低于苗圃地的幼苗出土率(约60%)。
2.2 不同生境幼苗高生长
苗圃地与野外山地的种子播种到幼苗出土再到幼苗生长停止所用天数大致相同(见表3),约266 d,但野外山地赤皮青冈的幼苗全年苗高平均值均低于苗圃地(29.1 cm),近山顶裂斗锥树下仅有1株苗(另一株枯死),且苗高生长量也最低,近山顶林缘的赤皮青冈幼苗苗高生长量最高,为9.98 cm,其次是混交林,值为9.13 cm(见表4);苗圃地出现两次高峰期,而野外山地的5种生境仅出现1次高峰期,说明赤皮青冈在近山顶裂斗锥树下的环境不适合赤皮青冈种子萌发,苗圃地的环境比较适合赤皮青冈的种子萌发。
2.3 苗高生长动态模型拟合
运用SPSS软件对赤皮青冈的苗高进行拟合(见表5),不同生境的拟合结果相关系数为0.984~0.958,平均值为0.969,苗圃地的相关系数高于其他野外山地的相关系数,为0.984,赤皮青冈林缘的相关系数最低,为0.958。通过公式(3)得出t、t1、t2,可将幼苗苗高的生长进程划分为生长前期、速生期和生长后期3个时期。结果见表6,不同生境的赤皮青冈幼苗苗高生长特点存在差异,赤皮青冈林缘地的赤皮青冈苗高生长峰值出现最晚(105 d);速生期持续时间最长的是母树林内和赤皮青冈林缘(115 d),持续时间最早的是近山顶林缘(107);6个不同生境的速生期占总生长期的比率在54.31%~58.51%之间,速生期内生长量占总生长量的比率在43.29%~55.19%之间,说明速生期在整个生长期中占比高,体现了速生期在幼苗苗高生长进程中的重要性。从图3可见,6个Logistics 拟合曲线的实测值与拟合值间的符合程度较高,具有较高的准确性,且基本符合“S”型生长规律,说明拟合效果好,拟合曲线的相关系数达到显著水平,结果较可靠。
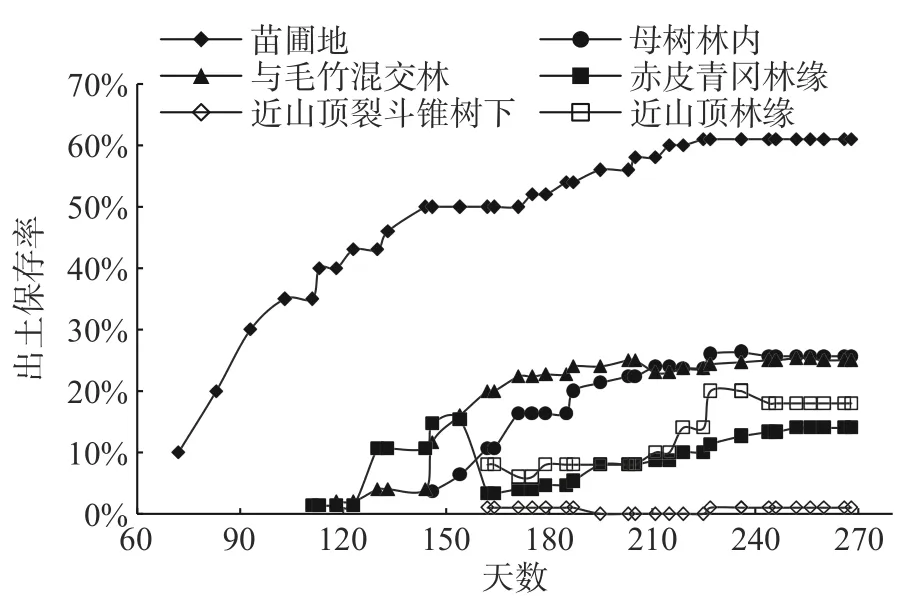
图1 苗圃地幼苗出土率Fig.1 Seedling emergence rate in nursery
3 讨论与结论
种子的萌发是种子生命进程的开始,萌发的速度影响植物在种群中的数量,影响植物在种群中的地位和作用[15]。本研究发现,赤皮青冈种子萌发总体呈现“慢-快-慢”的曲线,苗圃地的赤皮青冈种子萌发时间在5月27日,比野外的提早一个多月,且幼苗出土率也高于野外,可能是由于苗圃地海拔低,光照足等环境因子的影响;在野外,母树林内的赤皮青冈幼苗出土率和保存率最高,近山顶裂斗锥树下的最低。种子萌发阶段,植物便是从一生中风险最小阶段跨越到风险最大的阶段,也是植物最为脆弱的阶段[16],种子向幼苗和幼树过渡的时期是众多的珍稀濒危植物生长瓶颈之一[17]。苗圃地的赤皮青冈幼苗全年苗高平均值为29.1 cm,远高于野外山地的幼苗苗高平均值,可能是由于苗圃地的赤皮青冈种子萌发时间早于野外山地;在野外山地中的幼苗苗高平均值从从高到低依次排序为近山顶林缘>混交林>母树林>赤皮青冈林缘>近山顶裂斗锥树下,发现近山顶林缘的赤皮青冈种子保存率低,枯死数量达到总数量的一半,但生长量高,植株长势好,近山顶裂斗锥树下的赤皮青冈幼苗仅只有一株,且长势最差,近山顶处的光照比近山顶裂斗锥树下强,说明赤皮青冈喜光,在光照强度大的地方存活率较高。在种子萌发阶段,赤皮青冈种子更适合在海拔低的苗圃地进行培育;在野外山地,母树林以及毛竹混交林2个地点比较适合进行培育。
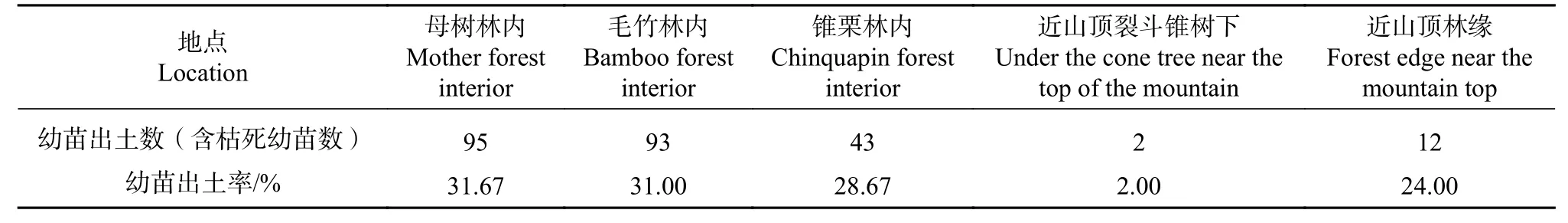
表2 野外幼苗出土率Tab.2 Seedling emergence rate in the field

表3 苗圃地幼苗生长情况Tab.3 Seedlings growth condition in nursery
利用Logistic方程对不同生境的赤皮青冈幼苗苗高进行拟合,描述赤皮青冈幼苗苗高随时间变化的生长规律,6个生境的相关系数均达到0.95以上,回归性均达到极显著水平(见图3)。赤皮青冈幼苗实测数据的曲线与拟合曲线基本吻合,生长曲线为典型的“S”型,且野外山地的赤皮青冈幼苗生长曲线与苗圃地的差别较大,从中发现人为适度的干预赤皮青冈幼苗的生长更有助于赤皮青冈种群的延续。根据Logistic方程上的2个拐点,将赤皮青冈幼苗苗高的生长进程准确地划分为生长前期、速生期和生长后期。在生长前期,赤皮青冈幼苗由于环境因素和自身条件的限制,生长缓慢,在苗圃地播种时,由于光照强烈且出苗时间早,应注意遮阳,覆膜保湿;在速生期,因赤皮青冈幼苗在速生期内生长量占总生长量的比率较大,比率在43.29%~55.19%之间,所以此时期是赤皮青冈幼苗苗期管理的关键时期,建议在苗圃地播种后40 d~60 d增加水肥管理,在正午时分盖上遮阳网,创造适宜的条件,以此延长其速生期的时间;在生长后期,幼苗苗高生长缓慢,在此期间建议对苗圃地的赤皮青冈幼苗进行适量的浇水,以防止幼苗徒长,再进行炼苗,以提高后期移栽的成活率。
由于试验条件以及资源的限制,本文研究的结果不能深入的进行动态变化分析。影响赤皮青冈幼苗生长的因素众多,要进行全面的调查以及了解其完全的生活习性和状态,需要在实际调查中不断的修正和调整。因此在此后的工作中,运用3S遥感技术进行全面客观的分析赤皮青冈苗木生长的各环节的特点,为提供更准确的数据及科学指导。
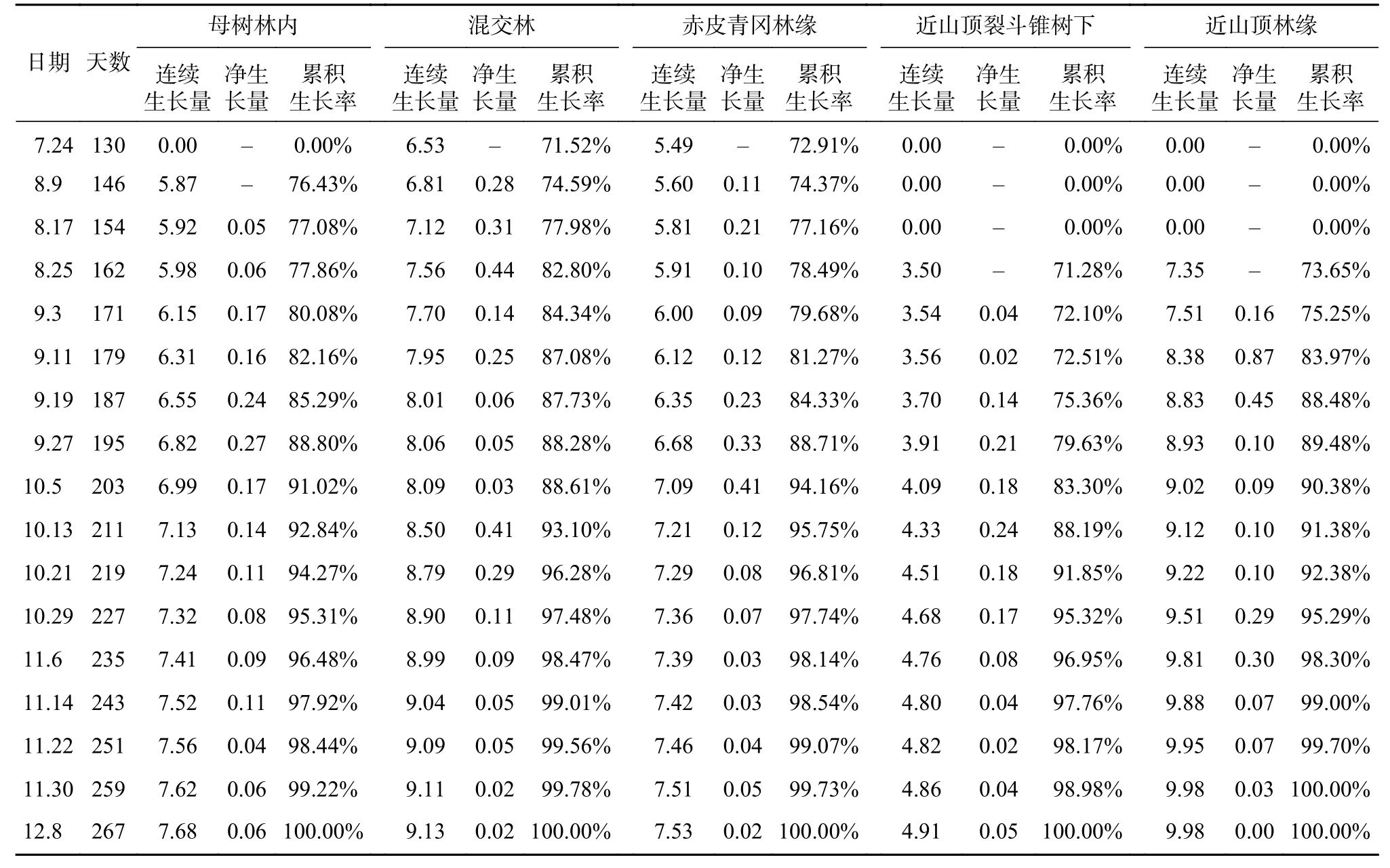
表4 野外山地幼苗生长情况Tab.4 Seedlings growth condition in the wild mountains
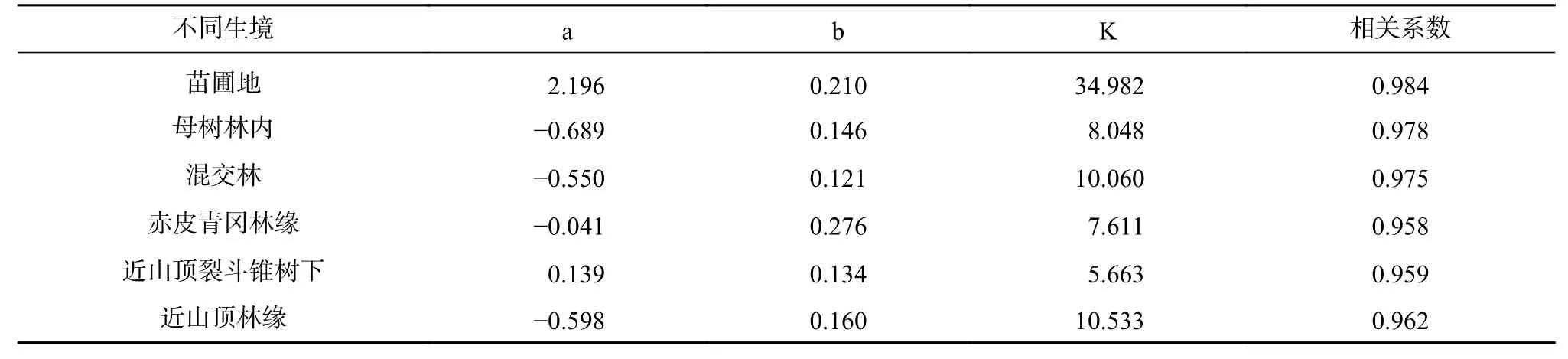
表5 幼苗苗高生长Logistic模型Tab.5 Logistic model of seedling height growth

表6 幼苗苗高生长特征值Tab.6 Seedling height growth characteristic values
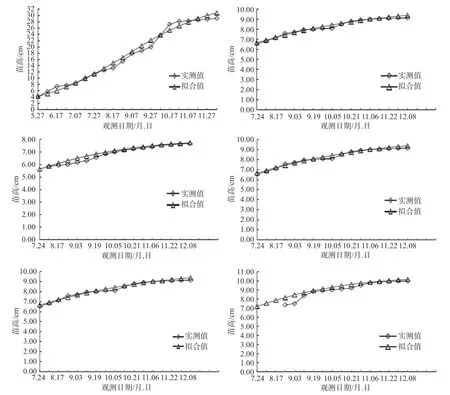
图3 赤皮青冈1年生幼苗苗高Logistics 拟合曲线Fig.3 Logistics fitting curve of seedling height of one-year-old C. gilva seedling
