罗氏沼虾组织和性别特异性表达基因的挖掘研究
2022-09-21任晋东陈红林牛宝龙詹炜楼宝
任晋东,陈红林,牛宝龙,詹炜,楼宝
(浙江省农业科学院水生生物研究所,浙江 杭州 310021)
罗氏沼虾(Macrobrachium rosenbergii)是长臂虾科、沼虾属动物,具有生长速度快、养殖存活率高、抗病力强和广泛的环境适应性等特点。原产于印度太平洋地区,生活在各种类型的淡水或咸淡水水域。20世纪60年代以来,先后移养于亚洲、欧洲、美洲等一些国家和地区,是全世界养殖量最大的淡水养殖虾类品种。1976年自日本引进我国,主要在南方10多个省(市、区)推广养殖,是我国重要的淡水养殖虾类引进品种。2020年,我国罗氏沼虾产量达14万t,超全球罗氏沼虾养殖产量的一半以上,且连续20年位列世界第一,但与以色列等国相比,我国每667 m均产仍相差100 kg以上。与其他甲壳类动物类似,罗氏沼虾也具有性别两态性,同龄雌、雄虾个体的大小、生长速度悬殊,单性化培育可提高我国罗氏沼虾单产,但关于罗氏沼虾个体消化发育、性别分化的分子机制尚不清楚。
目前,已有研究表明,组织和性别特异性基因是组织细胞分化、性别发育成熟的关键调控因子和标记分子。组织特异性基因在调控生物体器官再生、胚胎发育方面也有着重要的作用,其表达翻译也是组织器官发育成熟的标志及指示工具,在研究个体发育、胚胎发育过程中有着重要的作用,Wang等研究发现,组织特异性基因在南美白对虾(Litopenaeus vannamei)的免疫防御中也有着重要作用。性别特异性表达基因在生物性别发育分化、性别特异性表型形成、性别转化等过程中均有重要作用,目前关于甲壳类生物中老虎虾、南美白对虾的组织特异性遗传标记均有报道,但在罗氏沼虾器官组织和性别特异性基因挖掘方面,还未见报道。
现有研究表明,罗氏沼虾的眼柄、精巢、促雄性腺和卵巢等器官组织参与性别发育分化,而肝胰腺是罗氏沼虾体内分化形成的最大消化腺体组织。现以罗氏沼虾的眼柄、肝胰腺、精巢、卵巢和促雄性腺体各器官组织为研究对象,通过分析转录组数据,利用组织特异性参数(τ),挖掘罗氏沼虾各器官组织和性别差异表达基因,并进行功能分析,探明其作为组织和性别发育成熟标记(Marker)的潜力。
1 材料与方法
1.1 试验材料
采集的性成熟罗氏沼虾来自浙江省嘉兴市红宝石有限公司养殖基地,分别收集3只成年公虾的眼柄(Es)、肝胰腺(Hp)、精巢(Tt)、促雄性腺(AG)和3只成年母虾的眼柄、肝胰腺、卵巢组织(Ov),共计7个组织、21个样品,进行转录组测序分析。转录组RNA测序及注释由北京诺禾致源科技有限公司完成。转录组表达数据定量验证所用RNA提取试剂盒、反转录试剂盒和定量分析试剂盒购于南京Vazyme公司。
1.2 方法
1.2.1 转录组测序及注释
利用Clontech SMARTer PCR cDNA合成试剂盒,并按照BluePippin转录本片段,选择系统方案后,制备转录组测序文库。因罗氏沼虾无参考基因组序列,故将所有器官组织样品的混合样进行三代全长测序,获得所有基因序列,作为二代转录序列拼接比对。三代测序基因去除冗余序列和校正后,根据序列特异性形成唯一性转录本序列及基因ID。利用Blast软件,以P值<10为条件,分别在NT、NR、KOG、Swiss-Prot和KEGG数据库进行比对分析,以得分最高的作为该基因的注释基因名。
1.2.2 转录基因定量分析
对21个样品器官组织进行二代转录测序,组装后以三代鉴定基因结果ID为标识,统计各个基因的序列条数,用来计算每个样本中每个基因的相对表达量,并计算所有基因的相对表达量数据FPKM(每千个碱基的转录每百万映射读取的碎片)值。依据组织特异性和性别特异性分析结果,选择5个组织特异性高表达基因,设计定量分析引物(表1),进行qPCR定量结果验证分析,确定转录组基因相对表达量的可信度。

表1 qPCR定量分析引物
1.3 统计分析
利用DESeq的R软件包,按组织分组,进行差异基因表达比较分析,获得各器官组织特异性差异表达基因;以性别为分组,进行性别特异性差异表达基因分析,获得各性别特异性差异表达基因。以各器官组织各个基因平均相对表达量FPKM值为基础,利用软件R计算各器官组织的τ,计算公式如下:
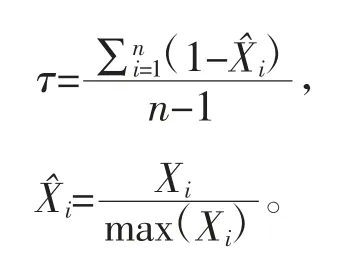
式中:X——基因在第i个器官组织内的相对平均表达量FPKM值;
i——试验包含的所有器官组织样品中第i个组织;

以τ>0.85为组织特异性基因和τ>1为组织唯一性高表达基因判定标准,鉴定组织特异性及唯一性表达基因。
2 结果与分析
2.1 转录组基因聚类分析及注释
21个罗氏沼虾样本器官组织,三代测序结果经去除冗余和校正后,获得唯一有效转录本21 345个,其中15 070个在至少一个注释基因库中存在对应的注释结果,另外有6 275个在所有注释库中无同源基因。另外雌雄罗氏沼虾的7个组织样品的平均FPKM值统计表明,精巢和卵巢平均FPKM值显著高于(P<0.05)其他组织(图1)。随机选择的5个基因相对表达量与其FPKM值呈极显著相关(P<0.01)。

图1 雌雄罗氏沼虾不同器官组织相对表达量FPKM值分布
2.2 组织特异性基因鉴定分析
以器官组织为分组单位进行差异表达基因分析,性成熟罗氏沼虾眼柄组织特异性高表达基因1 874个,特异性低表达基因3 409个;肝胰腺存在的特异性高和低表达基因分别为2 700和5 231个;精巢的特异性高和低表达基因分别有1 698和612个;促雄性腺体特异性高和低表达基因最少,只有270个和24个;卵巢的特异性高表达基因最多,有4 216个。各器官组织特异性高、低表达基因与其他组相对表达量比较分析表明,不论是高表达基因还是低表达基因,其平均相对表达量与其他器官组织都存在极显著差异(P<0.01),结果见图2(a)(b)(c)(d)(e)。

图2 罗氏沼虾器官各组织的组织特异性基因相对表达量
在剔除各组织中所有基因的相对表达量FPKM平均数<1的基因后,计算各基因的τ,获得τ≥1的组织唯一性基因835个,0.85≤τ<1的组织特异性高表达基因7 711个。分析获得眼柄、肝胰腺、精巢、促雄性腺和卵巢5个组织唯一性高表达基因数依次分别为33,29,39,5和729个,这个分布趋势与0.85≤τ<1的组织特异性高表达基因数分布一致(表2)。

表2 组织特异性基因按τ值大小分类统计
2.3 性别特异性基因鉴定分析
通过雌雄个体所有器官组织基因差异比较分析鉴定,获得罗氏沼虾性别特异性差异表达基因455个,其中雄性个体特异性高表达基因162个,雌性个体特异性高表达基因293个。剔除差异表达基因中同一性别中不表达或低表达(FPKM值<1)的差异表达基因后的性别表达基因数为217个,其中雄性和雌性特异性表达基因分别为87个和130个,且雄性高表达基因表达水平显著高于雌性,而雌性个体低表达基因组中平均表达水平显著高于雄性(图3)。另外对最终的性别差异基因分析τ值表明,性别特异表达基因41%以上的τ>0.8,26%表达基因>0.85。
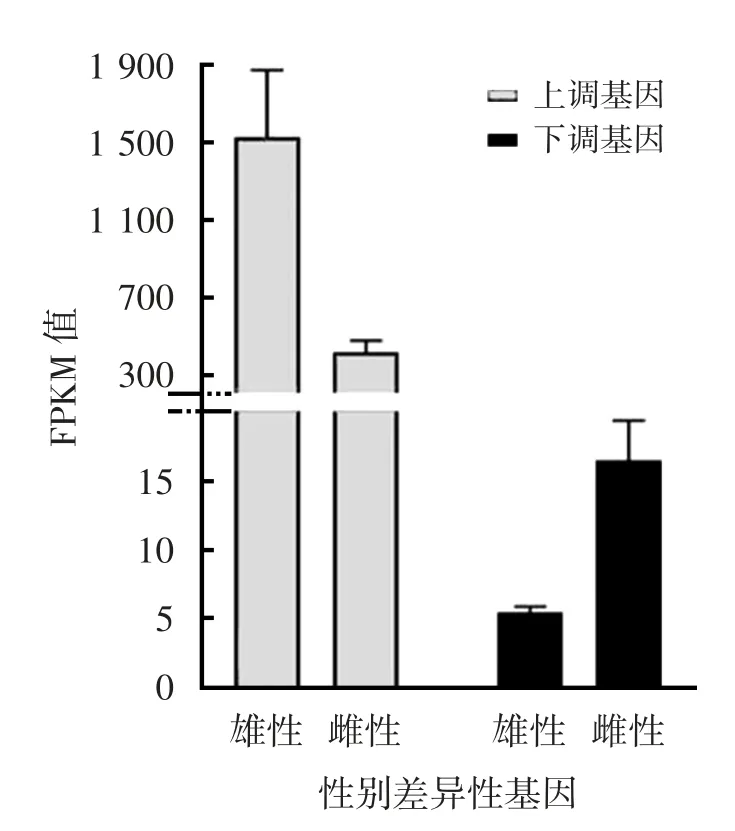
图3 罗氏沼虾性别差异基因平均相对表达量
通过对罗氏沼虾性别特异性表达基因的进一步分析发现,性别特异性表达基因在单一性别个体中的平均表达量均显著高于另一性别个体,雄性个体各器官组织相对表达水平显著高于雌性各器官组织,且各器官组织间同一性别相对表达量较为稳定一致(变异系数<0.5)[图4(a)]。如功能注释为人体寄生动物翅亚纲虱目动物假设蛋白的基因,该基因在雄性个体表现出显著的高表达特性,且同一性别各组织之间呈现变异系数<0.5的趋势。雌性性别特异性基因的在各器官组织的相对表达水平显著高于雄性,且变异系数也<0.5的基因。如功能注释为负鼠(Monodelphis domestica)核糖体蛋白大亚基L28e的基因,在雌性个体各器官组织表现稳定的表达峰度,且各器官组织均显著高于雄性个体[图4(b)]。
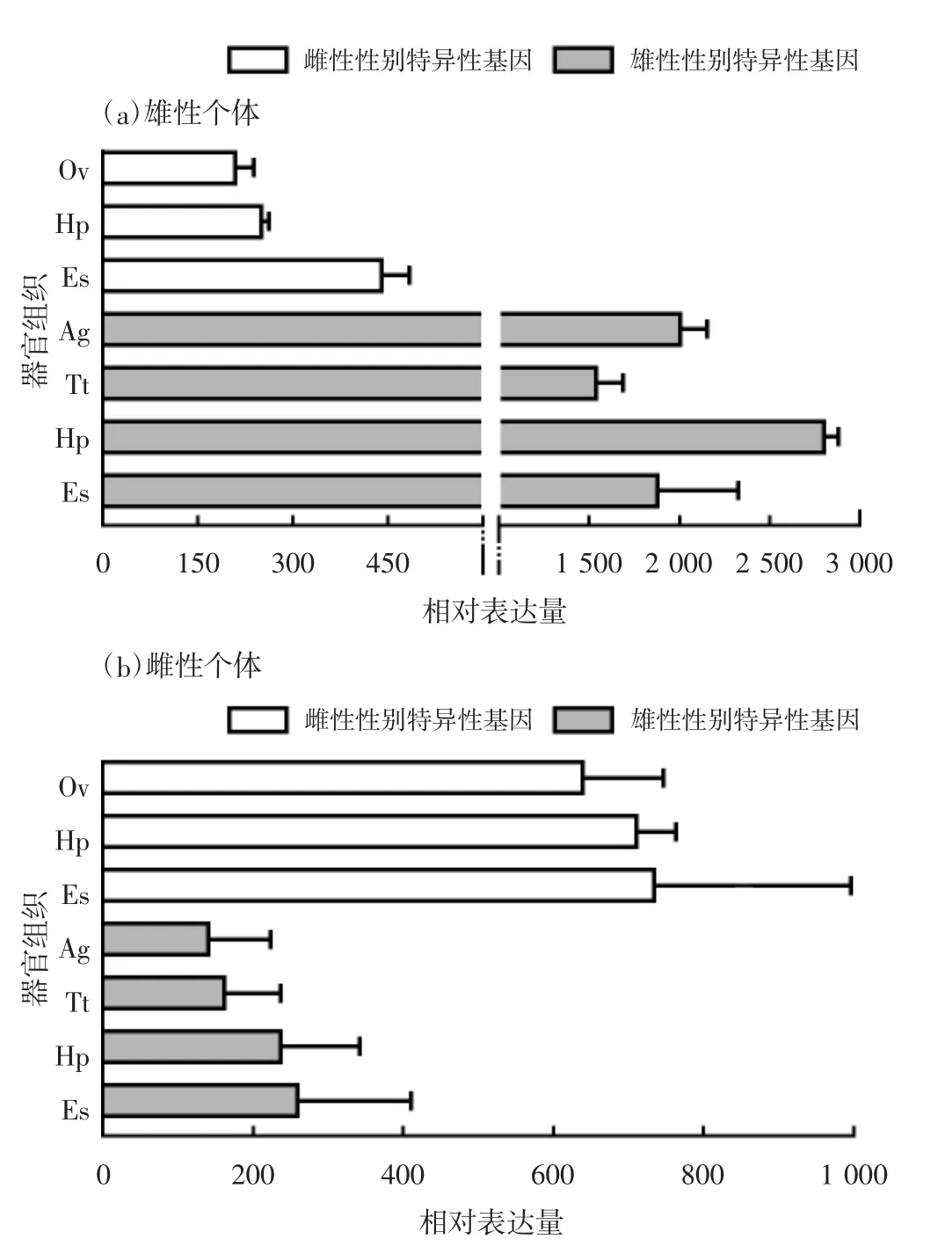
图4 性别特异性表达基因在罗氏沼虾不同器官组织中的相对表达量
3 讨论
3.1 罗氏沼虾性别及器官组织特异性基因挖掘进展
随着罗氏沼虾规模化繁育和人工选育技术的不断更新,罗氏沼虾已经初步实现全雄化和全雌化单性群体培育,也是目前唯一完成两个性别单性化培育的养殖虾类动物。其他养殖虾类品种,如南美白对虾和红螯螯虾(Cherax quadricarinatus),还处于关键基因探索阶段。虽然以色列研究人员已经报道了罗氏沼虾性别发育调控的关键基因iag(insulin-like hormone in androgenic gonad),但关于iag调控雄性个体性腺发育和雌性性腺逆转发育,特别是实现所有相关组织器官向雌性化方向逆转发育方面还未有明确报道。本研究通过挖掘罗氏沼虾雌雄个体消化和性腺组织器官特异性表达关键基因,提供了有针对性的组织和性别特异性的靶基因对象库,提高了性别发育的研究效率。
本研究采用的差异基因鉴定分析和τ,有效挖掘了罗氏沼虾各器官组织的特异性表达基因,建立了罗氏沼虾消化和性别决定5个主要器官的特异性表达基因库及器官性成熟的有效表达基因Marker,这与Kryuchkova-Mostacci等和Yanai等对τ的验证结果一致。目前组织特异性表达基因在生物器官组织功能、发育、甚至疾病的发生过程中都有重要的相应分子,而本研究挖掘的消化、性腺的组织特异性基因,弥补了罗氏沼虾发育研究中组织特异性标识基因缺少的空白。与眼柄器官视觉功能相关的组织特异性基因的发现表明,组织特异性表达基因与生物进化有着密切的关系,同类生物的组织特异性基因有高保守性特征。
3.2 罗氏沼虾视觉及消化腺特异性基因挖掘
本研究发现的罗氏沼虾眼柄组织特异性高表达基因主要包括两大类别功能基因。一类是KEGG(Kyoto Encyclopedia of Genes and Genomes)信号通路注释为与视觉功能相关的视紫红质基因,另一类为眼柄特异性的转录因子,如上游激活因子spp27的亚基基因。其中罗氏沼虾眼柄视紫红质基因与顶切叶蚁(Acromyrmex echinatior)、欧洲熊蜂(Bombus terrestris)、佛罗里达木蚁(Camponotus floridanus)、热带家蚊(Culex quinquefasciatus)等昆虫的视紫红质基因高度同源,眼柄特异性表达的上游激活转录因子基因与负鼠的高度同源,表明罗氏沼虾是甲壳类水生动物向陆地生昆虫演变的中间体,具有研究节肢动物从水生变为陆生的价值。
肝胰腺是虾类等甲壳类动物体内最大的组织器官之一和消化腺体,具有分泌消化酶、免疫、造血和免疫等功能,还具有为虾类动物性成熟储备营养物质的功能。本研究发现的肝胰腺唯一高表达基因有29个,其中5个在KEGG基因注释库中存在同源注释信息,其中3个分别为与非洲爪蟾(Xenopus tropicalis)抗原样蛋白E同种型X4基因、果蝇(Drosophila erecta)未定性蛋白和顶切叶蚁的酚氧化酶2基因高度同源,另外2个与蟒蛇(Python bivittatus)未定性蛋白高度同源。已有研究表明,酚氧化酶存在于虾类动物血细胞中,是虾类动物重要的免疫系统成员。本研究中,肝胰腺是酚氧化酶的唯一来源,表明肝胰腺不仅是罗氏沼虾的消化腺体,也是其唯一的造血器官。
3.3 罗氏沼虾性腺组织及性别特异性基因挖掘
在罗氏沼虾性腺组织器官精巢和卵巢的组织特异性基因挖掘中发现,卵巢组织特异性基因表达最为丰富。研究表明,与精巢相比,卵巢发育过程更加复杂,发育过程参与的基因数量更多。本文在精巢、促雄性腺和卵巢等3个性腺组织特异性基因数量方面的研究结果与现有的报道结果一致,即卵巢中存在更多唯一性高表达基因,其中峰度最高的基因为与KEGG注释库中欧洲熊蜂血细胞素同种型的X2基因。该基因是免疫球蛋白和半乳糖结合双功能域唾液酸酶的同源基因。在精巢唯一性高表达基因中已知注释功能的有参与染色体结构维持的中心体蛋白的rho鸟嘌呤核苷酸转换因子基因,该基因产物与成员蛋白协同作用形成中心粒,进而参与减速机体有丝分裂。虾类精子生成过程中精确控制染色体的均匀分布,该基因的高表达和现有研究表明,该基因是罗氏沼虾保证精子状态正常的重要因素。在促雄性腺体中发现的KEGG注释为四域蛋白酶抑制剂的基因,可裂解外源生物大分子,从而保护机体精源组织免受外源生物的侵扰。同时促雄性腺体还存在特异性高表达的丝氨酸蛋白酶抑制基因,而丝氨酸蛋白酶抑制基因是雄性动物精子顶体成熟的重要调节因子,也是精子成熟所必需。表明罗氏沼虾促雄性腺体具有与哺乳动物附睾类似的功能,是促进精子进一步成熟的组织器官。
对罗氏沼虾性别特异性表达基因研究发现,罗氏沼虾雄性个体在眼柄、肝胰腺、精巢和促雄性腺中存在共同的雄性特异性高表达、表达稳定的功能(变异系数小于0.5)基因,如与体虱(Pediculus humanus corporis)高度同源的假定蛋白T01223。雌性个体在眼柄、肝胰腺和卵巢中也存在共同的雌性特异性高表达、表达稳定的功能(变异系数小于0.5)基因,如与负鼠高度同源核糖体蛋白L28基因。研究表明,生物体雌雄个体相同组织之间差异基因占37%以上,但是多个组织共有且具有性别特异性的基因还未见报道。这也表明,罗氏沼虾等水生生物虽然具有ZW染色体系统的性别决定机制,但由于性别可塑性和单个基因决定的因素,该物种性别决定机制更为复杂,还需深入探索性别特异性基因如何与组织特异性基因协同发挥作用,并决定雌雄两性个体不同的表型特征。
