海洋哺乳动物尿素转运蛋白关键基因
——SLC14A2的分子进化
2022-09-17吴田振于振朋孙林霞郭天慧
曾 帆,吴田振,于振朋,孙林霞,郭天慧
(南京师范大学生命科学学院,江苏省生物多样性与生物技术重点实验室,江苏 南京 210023)
趋同进化(convergent evolution)是进化生物学的研究热点,是指亲缘关系较远的物种在自然选择的作用下,为了适应相似的生境而产生的相同或相似的表型、生理以及行为特征[1]. 尽管自然界中趋同进化的例子很多,但是趋同进化的分子机制研究还很少. 大熊猫(Ailuropodamelanoleuca)和小熊猫(Ailurusfulgensstyani),亲缘关系较远,通过基因组的测定,鉴定了多个与肢体发育和蛋白消化相关的基因出现趋同信号,为大、小熊猫演化出趋同的食竹食性以及具有抓握竹子的伪拇指提供了分子证据[2-3]. 蝙蝠和海豚在系统发生树上相距甚远,但均演化出回声定位的能力,是趋同进化的又一典型,近些年鉴定了与听觉相关的基因在二者之间具有平行/趋同进化位点,且在Perestin基因的系统发育树中聚在一起,为二者趋同进化提供了很强的分子证据[4-5].
海洋哺乳动物(marine mammals),也称为“海兽”,是哺乳动物中进化十分独特的类群,它们由不同的陆生祖先分别从陆地重返海洋,亲缘关系较远,但为了适应海洋环境,进化出一系列趋同的表型和生理特征,是研究趋同进化的理想模型. 它们都拥有与鱼类类似的流线型体型以减少在水中游动时的阻力[6];皮下脂肪层增厚用以保持体温,减少温度散失;渗透调节能力增强用以适应高渗的海洋环境;储氧能力提高用以增加在水中的屏息时长[7];骨密度的变化用以适应不同的潜水深度[8]. 除此以外,海洋哺乳动物普遍发生了味觉和嗅觉的丧失[9];鲸类和海牛发生了毛发的缺失[10],为趋同进化的研究提供了理想的模型.
渗透调节是生境转换后海洋哺乳动物共同面临的一个最主要的挑战. 形态学研究表明,许多海洋哺乳动物的肾脏演化出复肾结构,如鲸类、鳍足类、北极熊和水獭拥有数十甚至上千个小肾,且每一个小肾在形态结构和生理作用上都相当于陆生哺乳动物的独立肾,以此增加了肾脏的表面积,提高了肾脏的排泄能力,为海洋哺乳动物形成高浓度的浓缩尿提供了前提. 其中,完全营水生生活的鲸类的肾最为特殊,呈葡萄状,小肾数目在种间差别很大,体型较大的须鲸可达6 000~7 000个[11-12],而体型较小的淡水豚类小肾一般仅有65~170个[13]. 前人研究发现鲸类尿液的渗透浓度以及Na+浓度均显著高于大小与之相近或比之大的陆生哺乳动物,如鲸类尿液中尿素浓度比牛高4倍[14],提示鲸类通过排高度浓缩的尿液以调节水盐平衡. 然而,海洋哺乳动物渗透调节的分子机制的研究才刚刚开始. 研究发现肾脏中与渗透调节相关的基因在海洋哺乳动物中出现了高表达的现象[15],进一步为海洋哺乳动物具有高效的水盐调节提供了分子证据. Xu等发现许多与渗透调节相关的基因在鲸类谱系中显著受正选择作用,如水通道蛋白2(aquaporin2,AQP2)和尿素转运蛋白(urea transporters,UTA)编码基因SLC14A2显著受到正选择,提示鲸类可能通过增强对水和尿素的转运以产生高浓缩的尿液,以此适应高渗的海洋环境,进一步证实鲸类产生浓缩尿是维持体内水平衡的一个有效机制. 水盐调节系统——肾素-血管紧张素-醛固酮系统(RAAS)中两个关键的成员管紧张素转换酶(angiotensin converting enzyme,ACE)和管紧张素原(angiotensinogen,AGT)鉴定到多个正选择位点,特别是长江江豚中多个基因受正选择,提示鲸类水盐调节系统是维持渗透调节的又一重要机制[16-17].
尿素转运蛋白(Urea transporters)是特异性转运尿素的膜转运蛋白,对机体的渗透调节起着关键性的作用[18]. 其中,UT-A是目前已克隆出的尿素转运蛋白亚家族之一,共6个成员(UT-A1至UT-A6),由同一基因SLC14A2编码并经过不同的启动子启动转录后剪接形成. 在UT-A亚家族的成员中,UT-A1、UT-A2、UT-A3和UT-A4均在肾脏中表达,且SLC14A2基因敲除后尿素转运蛋白表达缺陷的小鼠表现出严重的尿素浓缩功能缺陷,说明此基因在机体渗透调节过程中发挥了及其重要的作用[19-20]. 鲸类中该基因显著受正选择,提示鲸类通过提高尿素转运能力,从而增强了机体的水盐调节功能. 然而,该基因在其他海洋哺乳动物中的进化以及是否与鲸类一样具有趋同的排高浓缩尿液的渗透调节机制尚不清楚. 据此,本研究下载了73个哺乳动物的SLC14A2基因,运用比较基因组学方法探讨了以下科学问题:(1)渗透调节基因SLC14A2在海洋哺乳动物不同类群中是否受到相似的选择压力?(2)海洋哺乳动物间是否具有尿素转运的趋同分子机制?
1 材料与方法
1.1 SLC14A2基因序列数据
本研究在哺乳动物中选取了73个物种,涵盖了15个目,其中海洋哺乳动物包括鲸类25个代表性物种,鳍足类6个物种以及海牛类的1个物种(海牛).SLC14A2基因序列是在NCBI数据库和OrthoMaM(Orthologous Mammalian Markers,http://orthomam2.mbb.univ-montp2.fr/OrthoMaM)数据库中下载得到. 对于基因组尚未注释的物种,通过本地BLAST,最终获得了73个哺乳动物SLC14A2基因的序列. 由于本地BLAST可能会造成相邻外显子拼接处存在非3倍数碱基的插入或缺失,所以采用了一系列针对于存在移码突变的序列比对方法,运用Macse软件中的AlignSequence、Exportalignment程序、Hmmcleaner程序、Prank程序以及Gblocks程序,确定获得序列为同源序列[21-23].
1.2 选择压力分析
氨基酸的非同义替代(non-synonymous substitution rate,dN)与同义替代(synonymous substitution rate,dS)的比值ω(dN/dS)常用作选择压力(selective pressure)的检测指标(Kimura,1979). 如果ω=1为中性选择(neutral selection);ω>1为正选择(positive selection);ω<1为纯化选择(negative selection). 由于影响功能基因的选择作用往往都只是发生在一些特定的支系的特定位点上,因此,为了检测海洋哺乳动物进化过程中经历了怎样的选择压力,本研究采用了基于密码子的最大似然法(maximum likelihood,ML)的PAML4.7软件包中的codeML程序,检测特定支系或者位点上所受的选择压力[24].
首先,仅针对33种海洋哺乳动物SLC14A2基因序列的数据集,运用位点模型(site modal)中的三对模型(M7vs M8、M8 vs M8a和M1a vs M2a)检测正选择位点. 运用似然率检测法(likelihoodratio tests,LRTs)评估两个模型之间是否存在显著差异,计算2ΔL值与自由度之间的卡方分布情况,若P值<0.05,则认为两个模型之间存在显著差异. 利用经验贝叶斯(bayes empirical bayes,BEB)方法计算M8和M2a模型中的正选择位点的后验概率(Posterior probability),如果后验概率≥0.8,即认为该位点是潜在的正选择位点[25]. 另外,使用在线网站Datamonkey(http://www.datamonkey.org/)中的固定效应似然法(fixed effects likelihood,FEL)进一步检测正选择位点,设置FEL显著性水平小于0.2[26]. 同时被两种ML方法检测的位点,为强烈的正选择位点.
其次,在所有哺乳动物73个物种的SLC14A2基因序列的数据集中,选择分支模型(branch model)和支位点模型(branch-site model)进一步探究正选择作用是否局限于某些特定的进化谱系. 分支模型主要包括free ratio模型和two ratio模型. Free ratio模型允许系统发育树上每一个进化谱系具有不同的ω值,而Two ratio模型允许前景支(即所感兴趣的支系)与背景支之间存在不同的ω值. 运用LRT分别检测Free ratio和Two ratio模型与零假设one ratio模型(即假设系统发生树上所有支系具有相同的ω值)是否差异显著. 如果模型显著,则通过Free ratio检测ω>1的正选择支系,运用Two ratio模型检测前景支海洋哺乳动物是否出现加速进化. 此外,进一步运用更加严格的分支位点模型(Ma)分析,该模型允许在特定支上的每一个位点都有独立的ω值,将进化谱系分为前景枝(foreground,即感兴趣的枝系)和背景枝(background),使用LRT对备择假设Ma(正选择模型:0<ω0<1,ω1>1和ω2≥1)和零假设Ma0(中性模型:0<ω0<1,ω1=1和ω2=1)进行比较. 当成对模型(Ma vs Ma0)差异显著且ω>1时,接受正选择模型Ma,且所检测的位点P>0.8时,被认为是强烈的正选择位点. 衡量枝位点模型显著性的P值则通过多重检验错误发现率(false discovery rate,FDR)予以矫正,校正后的P<0.05时则为模型显著.
1.3 海洋哺乳动物SLC14A2基因平行/趋同位点检测
为了验证海洋哺乳动物在二次入水过程中,是否发生了趋同进化,本研究使用PAMLv4.7中Codeml程序对每个基因进行祖先序列重建. 通过对海洋哺乳动物各枝系中尿素转运蛋白氨基酸的序列分析,寻找趋同氨基酸替代,得到海洋哺乳动物支系间从祖先到现生氨基酸变化. 根据预测的祖先位点的差异,编码区基因序列的氨基酸替换事件分为平行替换和趋同替换两种模式. 如果两个进化枝具有相同的祖先氨基酸状态,并衍生出相同的现生氨基酸,便认为该位点为平行替换位点;反之,如果两个进化枝具有不同的祖先氨基酸状态,并衍生出相同的现生氨基酸,则为趋同替换位点,这两种模式都被认为是海洋哺乳动物趋同进化的证据. 考虑到偶然氨基酸替代引起的干扰,本研究在JTT-fgen氨基酸替换模型下进行统计检验,比较实际观察到的趋同位点数目与随机期望出现的趋同位点数. 若观测值大于期望值,则认为存在氨基酸趋同或平行替换;反之,则不存在[27].
1.4 蛋白质三维结构
位于关键区域的氨基酸改变可能会通过影响构型,进而对蛋白质的功能产生影响. 为深入探究这些正选择位点是否位于重要的功能域,本研究使用印太瓶鼻海豚(Tursiopsaduncus)SLC14A2基因的核苷酸序列在I-TASSER网站上(http://zhanglab.ccmb.med.umich.edu/I-TASSER/)进行了蛋白三维结构的预测,并将检测到的正选择位点用PyMOL软件(http://pymol.sourceforge.net/)定位到了其空间结构中. 使用uniprot网站(http://www.uniprot.org/)查阅获取更多功能域的信息.
2 结果与讨论
2.1 选择压力分析
首先,运用位点模型对33个海洋哺乳动物数据集进行选择压力分析,LRTs结果显示在SLC14A2基因中正选择模型M8显著优于中性模型M7和M8a(P<0.001),且ω=1.955,提示这个基因显著受正选择作用. 然后,在M8模型中,使用BEB方法在SLC14A2基因中鉴定到了9个显著受到正选择的位点(35、462、463、467、468、469、470、512、741)且后验概率均大于0.8. 位点模型中的M1a vs. M2a较M8 vs. M8a模型所得到的结果更为稳定,因此又使用前者进一步检测,结果表明M2a(ω=6.517)模型明显优于M1a模型(P<0.001),并鉴定了2个正选择的位点(35、463),这两个位点同时也被M8模型鉴定到(表1). 另外,进一步使用DataMonkey中的FEL模型鉴定了4个正选择位点(35、90、276、735). 综合以上3种ML方法,共有 2个位点(35、463)至少被两种以上ML方法鉴定到,认为是强烈的正选择位点(表1).
其次,为了检测SLC14A2基因受到的正选择是否局限于特定的进化谱系,在所有哺乳动物谱系73个物种中,LRT检测发现free ratio模型显著优于one ratio模型(P<0.001),且在鲸类的多个支系中检测到显著的正选择信号,即须鲸类的共同祖先支、瓶鼻海豚和印太瓶鼻海豚最近共同祖先以及鼠海豚和小头鼠海豚共同祖先(ω>1,P<0.001),而在其他海洋哺乳动物类群以及陆生哺乳动物中均没有鉴定出显著的正选择. 另外,以32种海洋哺乳动物作为前景支(ω=0.17211),其他陆生哺乳动物为背景支(ω=0.13012),运用two ratio模型检测发现SLC14A2基因在海洋哺乳动物支系具有显著的加速进化(P<0.001)(图1).
由于分支模型和位点模型并不能同时考虑位点和支系发生的正选择作用,因此进一步采用了更为严格的支位点模型检测选择压力. 结果在小须鲸鉴定到显著的正选择,且鉴定了2个显著的正选择位点(592、598,表1). 综合M8、M2a、FEL以及支位点模型的方法,共有4个正选择位点(35、463、592、598)被认为是强烈的正选择位点.

表1 在海洋哺乳动物中用两种ML方法检测到的正选择位点Table 1 Positively selected sites detected using two maximum likelihood(ML)methods across cetacean phylogeny
2.2 海洋哺乳动物SLC14A2基因特异氨基酸突变位点和平行/趋同进化分析
为了探究海洋哺乳动物适应高渗的海洋环境是否进化出了特异位点,对SLC14A2基因进行扫描后发现海洋哺乳动物具有11个特异的氨基酸突变位点. 其中,鲸类具有5个特异突变位点(Y14H、T398A、L455H、Q485R、A707S);鳍足类具有6个特异突变位点(G64T、S99G、T111S、K518M、A527T、S529H,图1),其中第398、707位氨基酸位于尿素转运蛋白的跨膜结构域中,提示海洋哺乳动物可能通过尿素转运蛋白特异的氨基酸突变,提高机体在高渗环境下的适应能力.
进一步检测SLC14A2基因在海洋哺乳动物类群间是否存在趋同的分子机制,重建了该基因的祖先序列,并进行非同义平行/趋同氨基酸位点检测,鉴定了6个平行替换位点在海洋哺乳动物类群间表现出显著的趋同进化信号. 如图1所示,在鲸类谱系中鉴定了2个平行替换位点(L20S、R532S),如在20位氨基酸位点处具有一个平行氨基酸替代,中华白海豚的最近共同祖先支(n)与白鲸和一角鲸最近共同祖先支(v)均为丝氨酸(S),而其他哺乳动物支系均为亮氨酸(L). 在鲸类和鳍足类两个海洋哺乳动物类群中共鉴定了4个平行替换位点(H1P、A219V、L508T、H814I),例如在第1位氨基酸位点处具有一个平行氨基酸替代,本研究中使用的鳍足类所有物种和抹香鲸(Physetercatodon)支系均为P,而其他哺乳动物物种此位点为H;在219位点处,小须鲸(Balaenopteraacutorostrata)和夏威夷僧海豹(Neomonachusschauinslandi)鉴定了1个平行氨基酸替代(A219V). 这些位点经过JTT-fgen氨基酸替换模型统计检验,观测值大于期望值,表明检测到的位点是可靠的平行/趋同氨基酸替代位点.
不同海洋哺乳动物类群之间鉴定到的平行/趋同氨基酸提示为了适应共同的高渗环境海洋哺乳动物可能进化出了趋同的尿素转运机制,其中第219和814位氨基酸位点位于尿素转运蛋白重要的跨膜结构域,因此海洋哺乳动物可能通过跨膜结构域的改变,增强了对尿素的转运能力,从而对机体的水盐平衡进行调控.
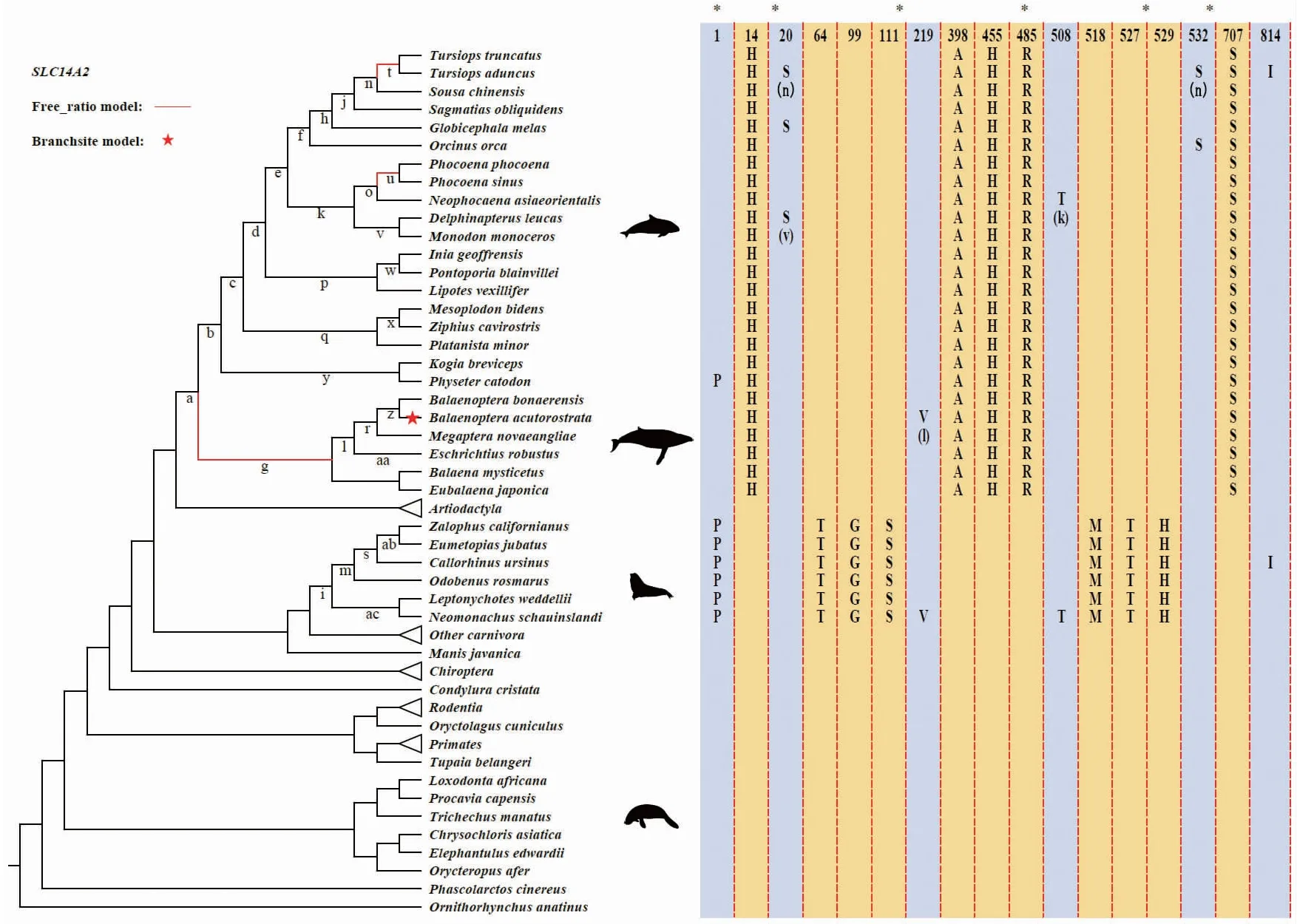
Free-ratio和支位点模型鉴定的正选择分别用红线和红五星标注. 右侧为SLC14A2基因在海洋哺乳动物中检测到的特异氨基酸位点(黄色)和平行/趋同位点(蓝色). 字母表示祖先枝. 作为外群的陆生哺乳动物包括鲸偶蹄目、食肉目、啮齿目等.图1 Free-ratio和支位点模型鉴定的海洋哺乳动物SLC14A2基因的正选择Fig.1 Evidence of positive selection across the phylogeny of marine mammals identified by free-ratio and branch-site models in SLC14A2 gene

强烈正选择位点和位于跨膜结构域的特异氨基酸位点分别用紫色和蓝色圆球表示.图2 SLC14A2蛋白质三维结构Fig.2 The three-dimensional structure of the SLC14A2 protein
2.3 SLC14A2基因三维结构分析
印太瓶鼻海豚(T.aduncus)的SLC14A2基因核苷酸序列在I-TASSER网站上(http://zhanglab.ccmb.med.umich.edu/I-TASSER/)进行了蛋白三维结构的预测,并运用PyMOL软件(http://pymol.sourceforge.net/)将强烈正选择位点和重要的特异氨基酸突变定位到3D结构中(图2). 这些位点大多位于重要的结构域或结构域附近,如第463位正选择位点位于蛋白激酶C的磷酸化位点S490附近;第35位正选择位点位于蛋白的极性氨基酸(polar amino acid)区域,此区域使得此段蛋白稳定位于膜外,同时此位点也与蛋白激酶C的磷酸化位点S26邻近[28],提示正选择位点可能与蛋白尿素转运功能相关. 另外,第398、707位特异氨基酸位点存在于蛋白重要的跨膜结构域. 跨膜结构域的改变通常与通道蛋白开放时间和转运效率密切相联系[29-30]. 因此海洋哺乳动物可能通过跨膜结构域的改变,增强了对尿素的转运能力,从而对机体的水盐平衡进行调控.
3 结论
趋同进化是进化生物学中的核心科学问题. 海洋哺乳动物不同类群是独立起源的,由不同的陆生祖先演化而来,亲缘关系相距甚远. 但是,为了适应高渗的海洋环境,海洋哺乳动物产生了趋同的渗透调节适应机制,具有很强的排盐保水能力,且能够产生高度浓缩的尿液. 先前的解剖学证据表明海洋哺乳动物具有复肾结构,通过增加肾脏表面积以提高肾脏的排泄能力[11-12],为海洋哺乳动物形成高浓度的浓缩尿提供了前提. 本研究发现尿素转运基因——SLC14A2在海洋哺乳动物发生了显著的加速进化(P<0.001),且有2个正选择位点至少被两种ML方法检测到,这两个氨基酸位点都位于蛋白激酶C的磷酸化位点附近,推测与蛋白尿素转运功能相关. 因为高渗透的内环境主要通过蛋白激酶C介导增加 UT-A1 和UT-A1A3的磷酸化,从而增强尿素的转运,且已有研究证实敲除SLC14A2基因则出现不能浓缩尿液的生理缺陷[19]. 因此本研究在海洋哺乳动物中鉴定的加速进化和正选择,提示可能增强了尿素转运能力从而维持了极高的渗透压以浓缩尿液,为海洋哺乳动物产生高浓缩尿液提供了分子证据. 另外,本研究通过对73个物种的SLC14A2基因进行比较,在鲸类和鳍足类中共鉴定出11个特异的氨基酸位点,其中2个氨基酸位于蛋白质重要的功能结构域,提示海洋哺乳动物可能通过这些氨基酸突变使尿素转运蛋白功能发生适应性的改变,从而增强尿素转运能力以适应高渗的海洋环境. 特别重要的是,在海洋哺乳动物三个类群间鉴定了6个平行/趋同进化位点,且其中两个位点位于蛋白质的跨膜结构域中,有研究表明跨膜结构域中单个氨基酸的改变即可改变通道蛋白的动力学和渗透特性[29-30]. 因此,推测海洋哺乳动物可能通过改变跨膜结构域的三维结构进而影响通道蛋白打开的时间和尿素转运的效率,进一步为海洋哺乳动物尿素转运蛋白在渗透调节中发挥重要功能以产生高度浓缩的尿液提供趋同的分子证据.
特别需要指出的是,用free-ratio模型和更严格的支位点模型鉴定的正选择均发生在鲸类谱系,可能与鲸类完全营水生生活面临较强的渗透调节压力有关. 之前生理学研究表明,鲸类尿液中尿素浓度比其近缘的陆生哺乳动物——牛高4倍[14],说明鲸类通过排高度浓缩的尿液以调节水盐平衡. 鲸类SLC14A2显著的正选择进一步证实鲸类通过增强尿素转运以产生高度浓缩的尿液. 另外,比较基因组和转录组学也在长江淡水江豚和海生江豚中分别鉴定了与尿素转运蛋白SLC14A2、ACE相关基因的特异的SNP,且发现两类江豚存在SLC14A2基因的表达差异,进一步为海生江豚产生高浓缩尿液(东亚江豚尿液:(74.96±20.06)mmol/L;长江江豚(40.7±15.78)mmol/L)提供了分子证据[17]. 先前研究已经证实水通道蛋白AQP2在鲸类中显著受正选择,AQP2基因的表达定位于肾脏的集合管,通过主动运输对水分进行重吸收,该基因显著的正选择提示鲸类可能通过增强水通道蛋白的功能而增强集合管对水的重吸收的能力,进一步为产生高度浓缩尿液提供了分子证据. 渗透调节是一个综合调控的过程,肾素—血管紧张素—醛固酮系统(RAAS)在哺乳动物水盐代谢调节中起重要作用. 该系统中关键基因血管紧张素转化酶ACE和血管紧张素原AGT在多个研究中均证实鲸类谱系显著受正选择[16-17],表明鲸类可能还通过RAAS调节系统更高效地维持鲸类的水盐平衡.
综上,本研究通过对尿素转运蛋白编码基因SLC14A2在海洋哺乳动物中的进化分析,揭示海洋哺乳动物SLC14A2基因的进化与渗透调节机制的关系. 分析结果表明海洋哺乳动物类群间SLC14A2基因的加速进化、正选择位点以及平行/趋同位点的鉴定,均为海洋哺乳动物趋同的产生高度浓缩尿液以保水提供了分子证据. 另外,正选择主要集中在鲸类进化谱系,可能与鲸类完全营水生生活面临更大的渗透调节的压力有关. 当然,SLC14A2基因在海洋哺乳动物中渗透调节的具体功能,将来还需要进一步的功能验证,以揭示该基因与渗透调节的关系.
