Role of insulin in pancreatic microcirculatory oxygen profile and bioenergetics
2022-09-16BingWeiLiYuanLiXuZhangSunJingFuBingWangXiaoYanZhangXueTingLiuQinWangAiLingLiMingMingLiu
INTRODUCTION
The concept of pancreatic islet microcirculation, which is currently under the spotlight[1,2], is responsible for coupling metabolic demands with glucose distribution and oxygen delivery in a manner involving microvascular endothelium-dependent vasodilation. Emerging evidence, including ours,indicates that the integrated pancreatic microcirculation in islets is necessary to maintain endocrine function and is involved in the pathogenesis of diabetes, partially through impairment of microcirculatory blood perfusion[3,4].
As part of a highly specialized microvascular system[5], pancreatic islets are richly vascularized and have 5-10-fold higher blood flow than the exocrine pancreas[6]. Pancreatic islet microvascular endothelial cells (IMECs) are therefore responsible for maintaining substance exchange and contribute to the dynamic regulation of glucose metabolism. Malfunction of IMECs is not only the culprit of deteriorated pancreatic islet microvascular blood perfusion and oxygen supply but also a victim of imbalanced energetic homeostasis.
Metabolic abnormalities in glucose are generally related to alterations in energy metabolism,especially at the onset of diabetes. The main organelle of IMECs responsible for energetic homeostasis is the mitochondrion, which plays a critical role in IMEC survival and death by regulating ATP synthesis through glucose metabolism, ROS generation, and apoptosis[7,8]. Furthermore, the metabolic profile of IMECs links the microcirculatory phenomena to the occurrence of pathological phenotypes. It is therefore important and rational to investigate the metabolic states of IMECs to clarify the microcirculatory pathological changes that occur under glucotoxicity.
Several reports have suggested the involvement of microcirculatory endothelial dysfunction in diabetes. However, knowledge surrounding the bioenergetics of IMECs related to insufficient microcirculatory oxygen is rather limited. We have established a new microcirculatory monitoring approach that integrates pancreatic microcirculatory partial oxygen pressure (PO
), relative hemoglobin (rHb) and hemoglobin oxygen saturation (SO
) using a multimodal device[9]. Thus, the purpose of this study was to describe the integrated pancreatic microcirculatory oxygen under glucotoxicity and to determine the impact of insulin on the microcirculatory oxygen and bioenergetic profile of IMECs.
MATERIALS AND METHODS
Animals
BALB/c mice were obtained from the Institute of Laboratory Animal Science, Chinese Academy of Medical Sciences (CAMS). The mice were bred and housed at 22 ± 1 °C with 55%-65% humidity under a 12 h:12 h light:dark cycle. The mice were randomly divided into three groups, including a type 1 diabetes mellitus (T1DM) model group, an insulin-treated group and a nondiabetic control group (all
= 3). T1DM was established by intraperitoneal administration of streptozotocin (STZ, 40 mg/kg) into the mice for five consecutive days. A level of blood glucose higher than 200 mg/dL was considered to indicate diabetes. Insulin was subcutaneously injected (1.5 IU/day) into the mice in the insulin-treated group to maintain the blood glucose within the normal range[10]. The animal experiments in this study were permitted by the Laboratory Animal Welfare and Ethics Committee, Institute of Microcirculation,CAMS (China).
The next day, I cannot help myself-I am drawn8 at the same time to that spot near the fence. Am I crazy for hoping she will come again? Of course. But in here, I cling to any tiny scrap9 of hope. She has given me hope and I must hold tightly to it.
Measurements of the microcirculatory oxygen profile
To assess pancreatic microcirculatory oxygen, we employed a multimodal auxiliary microcirculatory monitoring system established with a fiber-optic oxygen sensor (Precision Sensing, Regensburg,Germany) and an Oxygen to See device (LEA Medizintechnik, Giessen, Germany) to determine the SO
,rHb, and PO
. After anesthesia, the pancreas was gently exposed by a median abdominal incision, and the microcirculatory oxygen profile, including SO
, rHb, and PO
, was subsequently captured. These parameters were measured at three random sites of the pancreas in each mouse.
Establishment of the three-dimensional framework of the microcirculatory oxygen profile
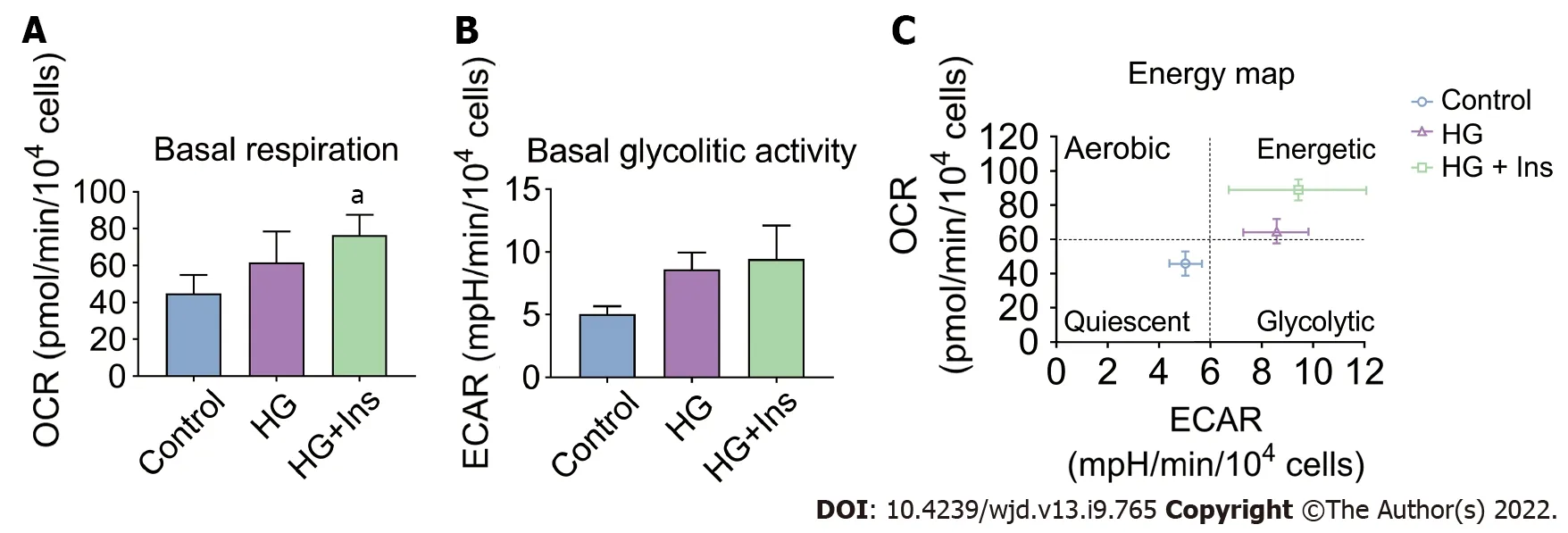
Insulin-treated IMECs exhibited significantly increased basal respiration in comparison with glucotoxicity-exposed IMECs (
< 0.05, Figure 4A). However, there were no significant differences in the basal glycolytic activity among the groups (all
> 0.05, Figure 4B). Moreover, an energy map was plotted based on the basal respiration and glycolytic activity in the IMECs. IMECs in the control group were in the quiescent quadrant (lower left). Glucotoxicity-exposed IMECs shifted to the energetic quadrant (upper right), reflecting increased mitochondrial activity. Insulin-administered IMECs were located in the right upper energetic quadrant, revealing more energetic metabolism (Figure 4C),suggesting that insulin increased the glycolytic activity of glucotoxicity-exposed IMECs when needed.
It was the afternoon of December 24, the day before Christmas; and as the newest hygienist in our office, I had to work. The only thing that brightened my day was the beautifully decorated Christmas tree in our waiting room and a gift sent to me by a fellow I was dating—a dozen long-stemmed red roses.
Video recording of the microcirculatory oxygen profile
ScreenToGif (version 2.19.3) was used to capture the dynamic 3-D framework. Each video was recorded in an MPEG4 file. The bitrate was 2000 Kbps with a 1920 × 1080 resolution ratio.
Transmission electron microscopy
Ultrastructural changes in the pancreatic islet microvasculature were examined using transmission electron microscopy (TEM). Fresh pancreatic tissue was fixed in 3% glutaraldehyde and 1% osmic acid and then passed through a graded series of dehydration and embedding solutions. Ultrathin sections(70 nm) were made after resin polymerization and uranyl acetate/Lead citrate staining. The samples were examined using a JEM-1400Plus transmission electron microscope (JEOL Ltd., Tokyo, Japan). The mitochondrial ultrastructure of IMECs was assessed.
Cell culture
The IMECs were purchased from ATCC (MS1, Manassas, VA, United States) and routinely cultured in DMEM supplemented with 10% FBS, 5.6 mmol/L glucose, 2% HEPES and 100 U of penicillin–streptomycin (Gibco, Carlsbad, CA, United States). After IMECs grew to confluence, the cells were divided into three groups, the control, glucotoxicity-exposed (HG, 25 mmol/L glucose), and insulin-treated (HG + Ins, 25 mmol/L glucose plus 10
mol/L insulin[10]) groups, and were treated for 24 h (
= 4 each group).
Bioenergetics assay
To investigate the effects of the high concentration of glucose with or without insulin on the bioenergetics of IMECs, an Extracellular Flux Analyzer (XFe24, Seahorse Bioscience, Billerica, MA, United States) was used to detect the real-time changes in energy pathways by directly probing the oxygen consumption rate (OCR) and extracellular acidification rate (ECAR). Briefly, IMECs were seeded in XFe24 culture plates at 1 × 10
cells per well. The cells were treated according to the abovementioned grouping for 24 h in DMEM with 0.5% FBS. The medium was subsequently replaced by Seahorse XF assay medium, and the cells were incubated without CO
for 1 h until detection.
For the mitochondrial stress test, mitochondrial function was monitored along with the sequential addition of oligomycin, carbonyl cyanide 4-(trifluoromethoxy) phenylhydrazone (FCCP) and rotenone/antimycin A (all 0.5 μM). Multiple respiratory indexes, including baseline metabolic functions(basal respiration, proton leak, ATP-linked respiration, non-mitochondrial respiration, and oxidative metabolism) and metabolic stress responses (maximum respiratory capacity [MRC] and spare respiratory capacity [SRC]), were analyzed and compared. In addition, an ECAR value was probed to indicate the basal glycolytic function when 10 mmol/L glucose was preadded into the medium before any pharmacological intervention. The ECAR-associated glycolytic capacity was subsequently reached after the injection of oligomycin. In this study, the values of both OCR and ECAR were normalized to 10
cells.
The woman who lies here was so remarkably18 stingy, that during her life she would get up in the night and mew, that her neighbors might think she kept a cat
Statistical analysis
SPSS software 21.0 (IBM, Armonk, NY) was used to perform the statistical analyses. The data are shown as the mean ± standard error of the mean. The data sets were subjected to one-way ANOVA and post hoc multiple comparisons. A
value under 0.05 was considered to indicate statistical significance. In addition, the correlation between the microcirculatory oxygen profile and bioenergetics of IMECs was analyzed by Pearson's method.
RESULTS
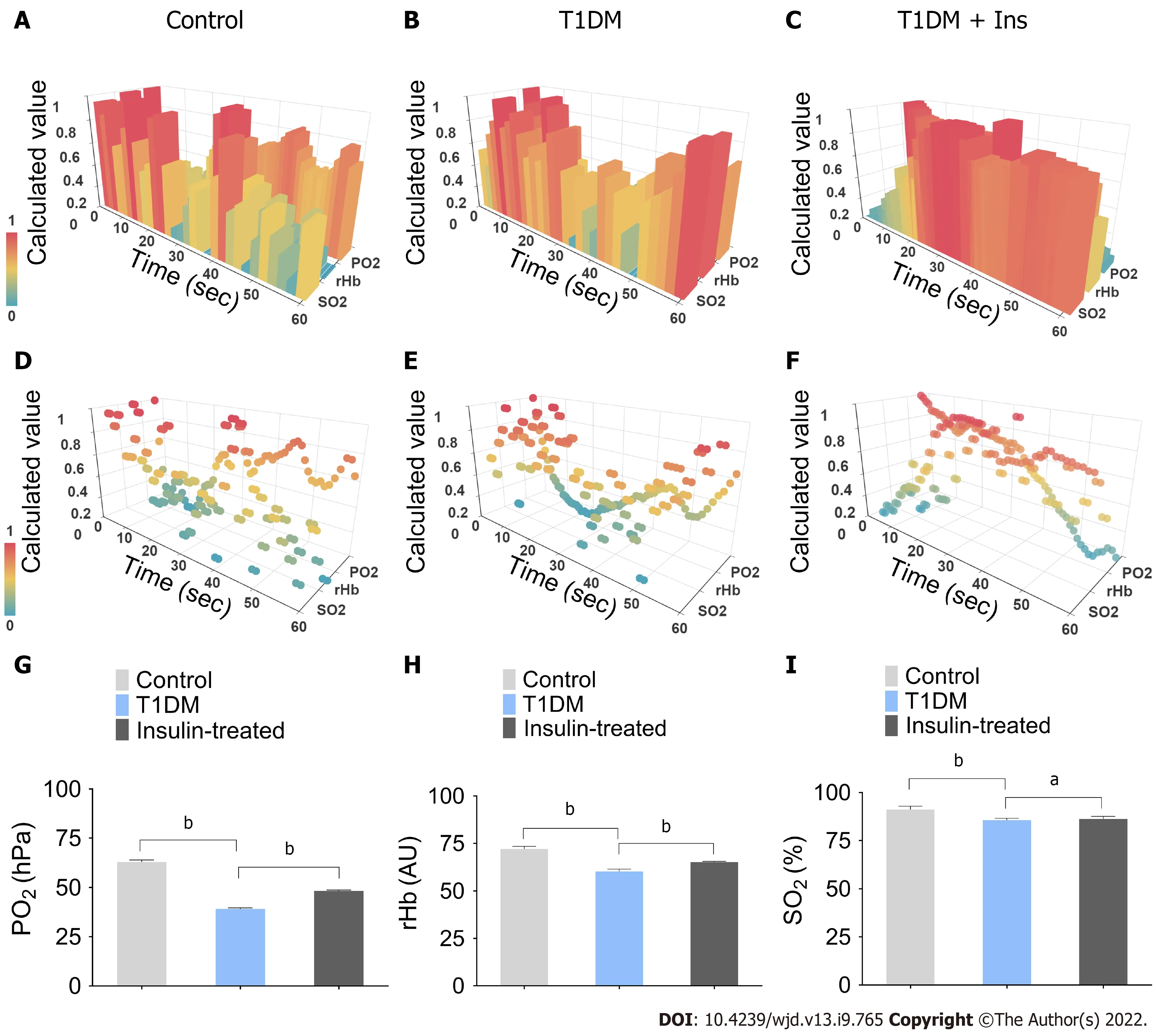
Insulin ameliorates the decrease in the integrated microcirculatory oxygen profile
In this study, the efficiency of STZ to induce T1DM mice model was 100%. To analyze the integrated microcirculatory oxygen profile of islet microcirculation, the preprocessed raw data were imported into the common microcirculatory framework. The oscillation of the microcirculatory oxygen profile is shown in histograms of the 3-D module (Figure 1A-C), and the distribution pattern of the microcirculatory oxygen profile was indicated using a scatter plot (Figure 1D-F, Videos 1-6). Loss of microcirculatory oxygen was observed in T1DM mice, which exhibited a significant decrease in PO
, rHb, and SO
compared with nondiabetic controls. Additionally, insulin administration ameliorated the STZ-induced decreases in these microcirculatory oxygen parameters (Figure 1G-I).
One day his queen presented him with a baby daughter as beautiful as the dawn, and the king himself was so happy and delighted that, for a whole week, he forgot to hunt, and spent the time in public and private rejoicing
But by this time night had come, and there was no light in the house, and the woman was in too much hurry to get to her treasures, to go and look for one
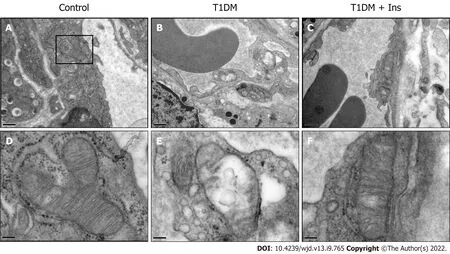
Insulin improves the mitochondrial ultrastructural abnormalities in IMECs in T1DM mice
Given that microvessels are responsible for distributing oxygen, we sought to determine whether the mitochondrial ultrastructure of IMECs changes in T1DM mice. TEM revealed the normal architecture of IMECs in the nondiabetic control group, with ovoid nuclei and well-arranged mitochondria in the cytoplasm. In contrast, mice with T1DM showed a narrowed or closed lumen with a contorted and thickened basement membrane, nuclear disaggregation, and mitochondrial swelling, suggesting an impaired ultrastructure of mitochondria in IMECs. The mitochondria in insulin-treated IMECs were restored, with a thin basement membrane, wide capillary lumen, and well-arranged parallel cristae(Figure 2). These data confirm the protective effect of insulin in the microcirculation of T1DM mice.
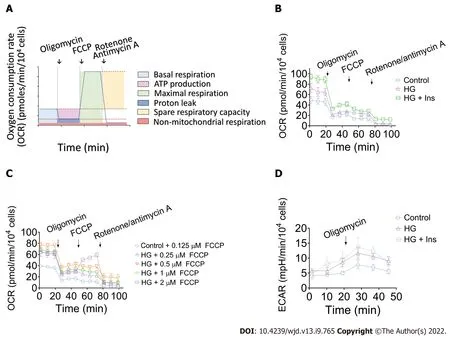
Effects of glucotoxicity and insulin administration on OCR and ECAR
The tight integration between endothelial metabolism and microcirculatory oxygen transport begs for integrative analysis that spans the cellular scale. We then performed real-time analysis of OCR and ECAR to determine energetic metabolic features in IMECs. The OCR of the IMECs was determined before and after receiving interventions of oligomycin, FCCP, and rotenone/antimycin A. A schematic of the real-time analysis of the IMEC OCR is depicted in Figure 3A. Our data revealed comparable mitochondrial maximal respiration in control, glucotoxicity-exposed, and insulin-treated IMECs, which was not induced after the injection of FCCP (Figure 3B).
The unique role of mitochondria in endothelial cells implies that a cell-regulatory function other than their energy-providing function is dominant[28]. Our previous study indicated that the microvascular blood perfusion of pancreatic islets was significantly decreased in T1DM mice but was partially restored after the administration of insulin[4]. Negative correlations were observed between the microcirculatory oxygen profile and metabolic indexes in the control group. In addition, a relatively low level of mitochondrial metabolism was detected in glycolysis-addicted IMECs, suggesting that glucotoxicity broke the negative correlation due to decreases in microcirculatory perfusion and the oxygen profile.
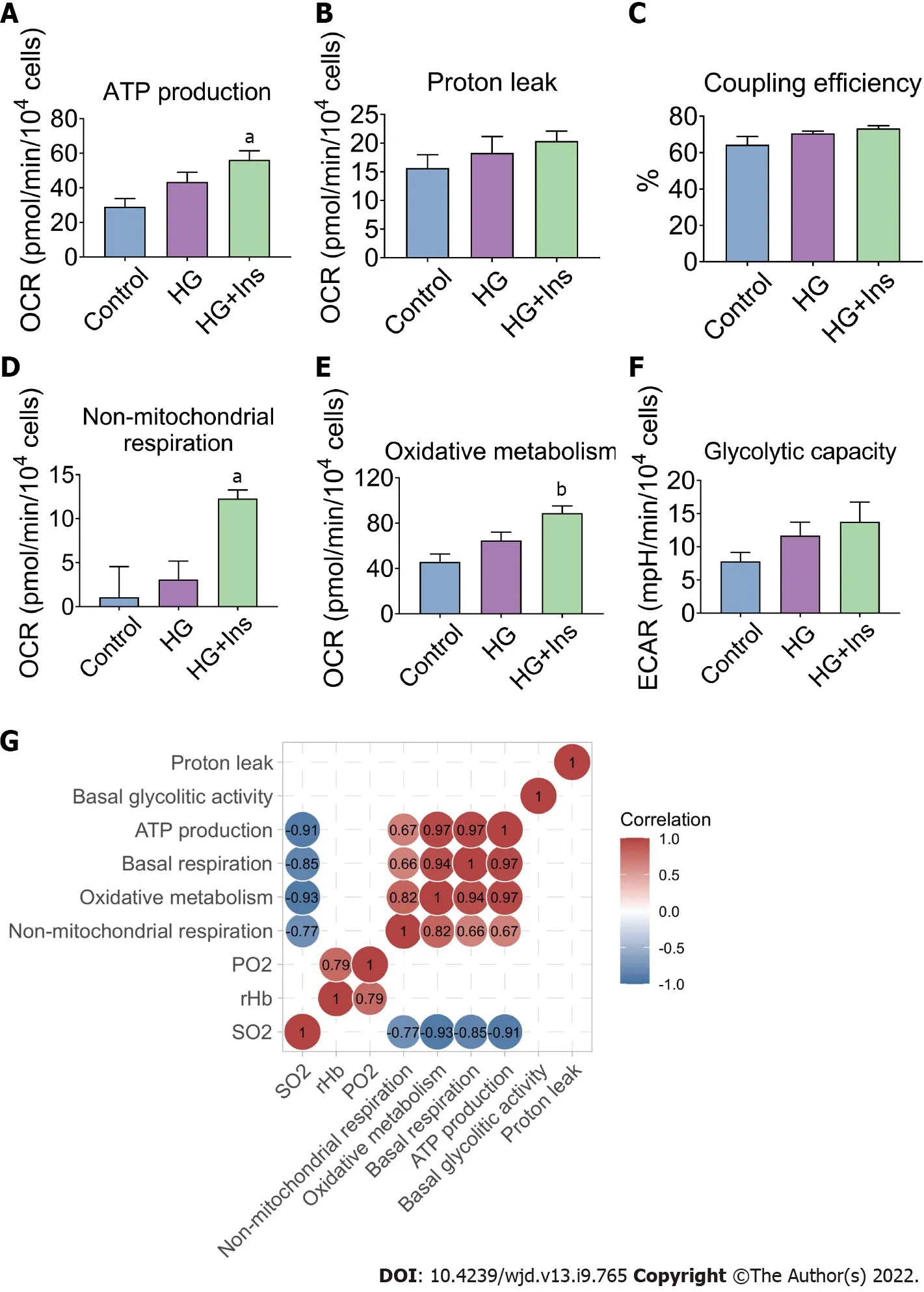
The basal respiration of mitochondria and non-mitochondrial respiration constitute oxidative metabolism in IMECs. Specific mitochondrial and non-mitochondrial functions were subsequently analyzed. ATP production, non-mitochondrial respiration, and oxidative metabolism were significantly increased in insulin-treated IMECs (
< 0.05, Figure 5A, D and E). However, proton leak (Figure 5B),coupling efficiency (Figure 5C), and endothelial glycolytic capacity (Figure 5F) were comparable among the groups.
Around seven o’clock that evening, however, my day took a turn for the better. I was in the middle of making another pot of vanilla9 hazelnut decaf when the customer from the day before drove up to the window. But instead of ordering anything, he handed me a single pink rose and a little note. He said that not too many people take the time to care about others and he was glad there were still people like me in the world. I was speechless and very touched; I hadn t thought that I had done anything incredible. After a moment, I came to my senses and thanked him. He told me I was welcome and with a friendly wave he drove away.
Insulin administration increases basal respiration and glycolytic capacity
Python (ver 3.7.4) and Apache ECharts (ver 4.2.0-rc.2) were used to generate a three-dimensional framework to visualize the pancreatic microcirculatory oxygen profile. In the 3-D framework, the X-, Y-,and Z-axes represented the time course, microcirculatory oxygen variables, and calculated values of the microcirculatory oxygen profile, respectively. The outliers were adjusted by the box plot algorithm.Additionally, the least common multiple algorithm was used to adjust the time granularity. Min-max normalization was conducted to transform the dimensions of multiple parameters.
Insulin administration activates oxidative metabolism and alleviates glucotoxicity-induced microcirculatory failure in IMECs
Subsequently, to determine whether FCCP concentration is responsible for the evaluation of the MRC, the IMECs were incubated with different concentrations of FCCP (0.125, 0.25, 0.5, 1, and 2 μM) in the control and HG groups. Surprisingly, none of the OCR values exceeded the corresponding basal OCR after the FCCP injections, suggesting that the IMECs do not have SRC (Figure 3C). The ECAR values were simultaneously measured to reflect the glycolytic activity of IMECs. After 0.5 μM oligomycin injection, the glycolytic capacity was recorded as the peak value of ECAR (Figure 3D).
The correlation between the microcirculatory oxygen profile and bioenergetics of IMECs was then analyzed. Significant negative correlations between microcirculatory SO
and bioenergetic parameters(ATP production, basal respiration, oxidative metabolism, and non-mitochondrial respiration) were found by Pearson’s correlation analysis (Figure 5G). These lines of evidence further confirmed that glucotoxicity in IMECs was related to pancreatic islet microcirculatory failure that could be reversed by insulin administration.
DISCUSSION
Insulin administration ameliorated the glucotoxicity-induced decreases in microcirculatory oxygen parameters (PO
, rHb, and SO
) and improved the mitochondrial ultrastructural abnormalities in IMECs. Insulin-treated IMECs exhibited significantly greater basal respiration than glucotoxicityexposed IMECs. An energy map revealed increased energetic metabolism in insulin-treated IMECs,with significantly increased ATP production, non-mitochondrial respiration, and oxidative metabolism.Significant negative correlations were revealed between microcirculatory SO
and bioenergetic parameters.
Considering islet β cells, rather than IMECs, are specific target of STZ. Therefore, STZ-involved T1DM animal models are useful in elucidating the mechanisms of diabetic microvascular endothelial pathogenesis. IMECs are the key determinants in pancreatic islet microcirculation homeostasis. Blood perfusion and oxygen transport requires the coordinated communication of mitochondria with metabolic demands, which is influenced by a variety of factors (including hypoxia)[12]. Coinciding with the impairment in the microcirculatory oxygen profile, pathological alterations in mitochondrial ultrastructure and other subcellular structures have been observed in IMECs of T1DM mice. Earlier studies have reported that defects in mitochondrial function correlate with mal-matching adenosine triphosphate generation[13,14], which interferes with the bioenergetics of pancreatic islet microcirculation. Treatment with insulin during glucotoxicity exposure resulted in restoration of the ultrastructure of IMECs. Thus, our data suggest that insulin can improve the functional status of pancreatic islet microcirculation.
Metabolic capacity is important for energy regulation and the maintenance of cell survival[15]. In parallel with damage to the ultrastructure of IMECs, biogenetic mechanisms act during glucotoxicity exposure to compensate for the decreased blood perfusion and oxygen distribution. IMECs supplied with insulin increase their basal respiration and ATP production and switch to energetic adaptation.Mitochondria are important organelles for ATP production[16]. Dysfunction of mitochondria is one of the key determinants in the pathogenesis of diabetes[17].
Then the third son said, Father, let me also show you something; and, as it was beginning to rain, he drew his sword and swung it in cross cuts above his head, so that not a drop fell on him, and the rain fell heavier and heavier, till at last it was coming down like a waterspout, but he swung his sword faster and faster, and kept as dry as if he were under cover
One week after graduation Rose died peacefully in her sleep. Over 2,000 college students attended her funeral in tribute to the wonderful woman who taught by example that it s never too late to be all you can possibly be. When you finish reading this, please send this peaceful word of advice to your friends and family, they ll really enjoy it! These words have been passed along in loving memory of ROSE.
Unexpectedly, our results indicated that maximal respiration of the mitochondria was not induced after injection of FCCP. Multiple factors are associated with FCCP-induced maximal respiration of mitochondria[18]. Therefore, to exclude the effect of FCCP concentration, we subsequently tested five FCCP concentrations, but none caused the basal OCR value to be exceeded, suggesting that the IMECs do not have SRC. In addition to the organ- and tissue-specific nature of microvascular endothelial cells[19], one of the possible explanations is that IMECs generate more than 85% of their ATP through glycolysis[20], which does not require an excessive number of mitochondria to obtain energy.
Furthermore, the increased OCR was associated with non-mitochondrial respiration, suggesting the existence of extensive ROS signaling caused by increased enzymatic activity of nitric oxide synthases,NADPH oxidase, and other oxygenases[21,22]. Although the glycolytic metabolism of endothelial cells is a protective strategy against oxidative stress[14], insulin can increase ROS production
activation of non-mitochondrial respiration
. The excessive ROS levels and increased oxidative stress may lead to mitochondrial dysfunction[23] and endothelial dysfunction[24]. In this energy-demanding process,quiescent endothelial cells divide and migrate to form new vessels[25], and excessive ROS synthesis inhibits angiogenesis by inducing excessive ROS synthesis[26].
In vain the Giant showed me his menagerie, which was entirely composed of children who would not work! Nothing did me any good, and at last I was reduced to drawing water for the dyeing of the wools, and even over that I was so slow that this morning the Giant flew into a rage and changed me into a gazelle
Similar to basal respiration, the basal glycolytic activity increases when insulin is present, although no significant difference was noted. An
study indicated that insulin, in the context of high glucose,significantly activates oxidative metabolism other than glycolysis in IMECs, although endothelial cells are considered “glycolysis addicted”[27]. The OCR measurements for oxidative metabolism can be divided into three components, including OCR associated with ATP production, proton leak, and nonmitochondrial respiration; the first two indicators together constitute the basal respiration of the mitochondria. Increased ATP production-associated OCR was found in IMECs after insulin treatment,suggesting that mitochondrial energy metabolism participates in the regulatory effects of insulin on microvascular endothelial mitochondrial injury.
29. No, a living creature is dearer to me than all the treasures in the world: His refusal of this offer shows how Rumpelstiltskin prefers a person for his prize than riches. It also implies that his intention was to win the baby from the beginning of his participation in the story. He was happy to accept the maid s necklace and ring before, but he took those so he could reduce her to agreeing to the horrible arrangement they made previously. Return to place in story.
The current study is the first report on the relationship between pancreatic microcirculatory oxygen profile and microvascular endothelial mitochondrial metabolism. However, there are still several limitations. First, the sample size of mice in each group was limited. Although pancreatic microcirculatory oxygen profile was measured at three random sites of the pancreas in each mouse, large sample size is preferred to ensure the data are representative. Second, in an interdependent functional relationship with β cells, IMECs are involved not only in the delivery of oxygen, but affect adult β cell function and promote β cell proliferation
vasoactive substances. However, the phenotypic and functional crosstalk between IMECs and islet β cells are not involved in our study.
CONCLUSION
In conclusion, glucotoxicity deteriorates the integrated pancreatic microcirculatory oxygen profile and bioenergetics, but this deterioration can be reversed by insulin administration.
ARTICLE HIGHLIGHTS
Research background
The pancreatic islet microcirculation adapts its metabolism to cope with limited oxygen availability and nutrient delivery. In diabetes, the balance between oxygen delivery and consumption is impaired.Insulin has been proven to exert complex actions promoting the maintenance of homeostasis of the pancreas under glucotoxicity.
Research motivation
the Beijing Municipal Natural Science Foundation, No. 7212068; the National Natural Science Foundation of China, No. 81900747.
Research objectives
To test the hypothesis that insulin administration can improve the integrated pancreatic microcirculatory oxygen profile and bioenergetics.
Research methods
A three-dimensional framework was generated to visualize the pancreatic microcirculatory oxygen profile. The microcirculatory partial oxygen pressure (PO
), relative hemoglobin (rHb) and hemoglobin oxygen saturation (SO
) were evaluated in nondiabetic, type 1 diabetes mellitus (T1DM), and insulintreated mice. An Extracellular Flux Analyzer was used to detect the real-time changes in bioenergetics by measuring the oxygen consumption rate and extracellular acidification rate in islet microvascular endothelial cells (IMECs).
Research results
The influence of microcirculatory disturbance on the development of diabetes mellitus has been highlighted over decades[11]. However, the current data associated with pancreatic microcirculatory oxygen profiles are deficient. Here, we used a computer algorithm-based common microcirculatory framework to reveal integrated pancreatic microcirculatory oxygen profiles among groups. The existence of microcirculatory hypoxia in T1DM was noted.
Research conclusions
Glucotoxicity deteriorates the integrated pancreatic microcirculatory oxygen profile and bioenergetics,but this deterioration can be reversed by insulin administration.
Research perspectives
Our understanding of the physiology and pathology of the pancreas islet microvascular endothelial cell in T1DM has been continually enhanced with the advancement of microcirculatory technology in parallel with rapidly developing bioenergetics that allows us to increase resolution and precision in our investigations.
ACKNOWLEDGEMENTS
We thank Xi-Nan Duan for discussions about the computational algorithms.
FOOTNOTES
Liu MM designed the experiments; Li BW, Li Y, Zhang X, Fu SJ, Wang B, Zhang XY, Liu XT,Wang Q and Li AL performed the experiments; Li BW, Li Y and Liu MM analyzed the data; Li BW and Liu MM wrote the manuscript; Liu MM made critical revisions to the article for important intellectual content; All authors discussed the results and approved the final version of the manuscript.
We tried to provide new insight into the relationship between pancreatic microcirculatory oxygen profile and microvascular endothelial mitochondrial metabolism.
So they went home, and Little Two-eyes again left her little dish untouched, and Little One-eye could not tell her mother why she would not eat, and said as an excuse, I was so sleepy out-of-doors
The study was reviewed and approved by the Institutional Review Board at the Institute of Microcirculation, Chinese Academy of Medical Sciences & Peking Union Medical College.
All animal experiments conformed to the internationally accepted principles for the care and use of laboratory animals, and approved by the Institutional Animal Care and Use Committee at the Institute of Microcirculation, Chinese Academy of Medical Sciences & Peking Union Medical College.
The authors declare no conflict of interest.
No additional data are available.
The authors have read the ARRIVE guidelines, and the manuscript was prepared and revised according to the ARRIVE guidelines.
Full of remorse26 and sorrow, he begged for her forgiveness, and Helena, only too pleased to have got him back again, did not, you may be sure, keep him waiting very long for her pardon, and so they were married and returned to Helena s castle, where they are no doubt still sitting happily together under the lime-tree
Bill was in actual fact, not so much a man of the world, as a man of Basford and Hartshill, but here was a life lived in an extraordinary richness of simplicity, a wealth of minute experience and also, I always felt, a whole ocean of experience that only Bill knew of, with which he sat at night in his back room alone, and unpacked secretly like a cut glass set, and repacked again before morning. I felt that there was more to Bill than he ever let on.
This article is an open-access article that was selected by an in-house editor and fully peer-reviewed by external reviewers. It is distributed in accordance with the Creative Commons Attribution NonCommercial (CC BYNC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited and the use is noncommercial. See: https://creativecommons.org/Licenses/by-nc/4.0/
China
Bing-Wei Li 0000-0002-0373-446X; Yuan Li 0000-0002-0065-9038; Xu Zhang 0000-0003-4499-2331; Sun-Jing Fu 0000-0002-2064-0537; Bing Wang 0000-0002-7287-0852; Xiao-Yan Zhang 0000-0002-4725-1849; Xue-Ting Liu 0000-0002-4281-4462; Qin Wang 0000-0003-4575-0457; Ai-Ling Li 0000-0003-4937-4700; Ming-Ming Liu 0000-0002-6750-5068.
Chang KL
A
Chang KL
1 Dybala MP, Gebien LR, Reyna ME, Yu Y, Hara M. Implications of Integrated Pancreatic Microcirculation: Crosstalk between Endocrine and Exocrine Compartments.
2020; 69: 2566-2574 [PMID: 33148810 DOI:10.2337/db20-0810]
2 Dybala MP, Kuznetsov A, Motobu M, Hendren-Santiago BK, Philipson LH, Chervonsky AV, Hara M. Integrated Pancreatic Blood Flow: Bidirectional Microcirculation Between Endocrine and Exocrine Pancreas.
2020; 69:1439-1450 [PMID: 32198213 DOI: 10.2337/db19-1034]
3 Liu M, Zhang X, Li A, Wang B, Li B, Liu S, Li H, Xiu R. Insulin treatment restores islet microvascular vasomotion function in diabetic mice.
2017; 9: 958-971 [PMID: 27976498 DOI: 10.1111/1753-0407.12516]
4 Li Y, Li B, Wang B, Liu M, Zhang X, Li A, Zhang J, Zhang H, Xiu R. Integrated pancreatic microcirculatory profiles of streptozotocin-induced and insulin-administrated type 1 diabetes mellitus.
2021; 28: e12691 [PMID:33655585 DOI: 10.1111/micc.12691]
5 Muratore M, Santos C, Rorsman P. The vascular architecture of the pancreatic islets: A homage to August Krogh.
2021; 252: 110846 [PMID: 33197561 DOI: 10.1016/j.cbpa.2020.110846]
6 Jansson L, Carlsson PO. Pancreatic Blood Flow with Special Emphasis on Blood Perfusion of the Islets of Langerhans.
2019; 9: 799-837 [PMID: 30892693 DOI: 10.1002/cphy.c160050]
7 Wai T, Langer T. Mitochondrial Dynamics and Metabolic Regulation.
2016; 27: 105-117[PMID: 26754340 DOI: 10.1016/j.tem.2015.12.001]
8 Bertero E, Maack C. Calcium Signaling and Reactive Oxygen Species in Mitochondria.
2018; 122: 1460-1478[PMID: 29748369 DOI: 10.1161/CIRCRESAHA.118.310082]
9 Li Y, Song X, Liu M, Wang B, Zhang J, Li A, Zhang H, Xiu R. Multimodal Device and Computer Algorithm-Based Monitoring of Pancreatic Microcirculation Profiles In Vivo.
2020; 49: 1075-1082 [PMID: 32769856 DOI:10.1097/MPA.0000000000001627]
10 Wang B, Zhang X, Liu M, Li Y, Zhang J, Li A, Zhang H, Xiu R. Insulin protects against type 1 diabetes mellitus-induced ultrastructural abnormalities of pancreatic islet microcirculation.
2020; 69: 381-390 [PMID: 32648910 DOI: 10.1093/jmicro/dfaa036]
11 Strain WD, Paldánius PM. Diabetes, cardiovascular disease and the microcirculation.
2018; 17: 57[PMID: 29669543 DOI: 10.1186/s12933-018-0703-2]
12 Leone TC, Kelly DP. Transcriptional control of cardiac fuel metabolism and mitochondrial function.
2011; 76: 175-182 [PMID: 22096028 DOI: 10.1101/sqb.2011.76.011965]
13 Wong BW, Marsch E, Treps L, Baes M, Carmeliet P. Endothelial cell metabolism in health and disease: impact of hypoxia.
2017; 36: 2187-2203 [PMID: 28637793 DOI: 10.15252/embj.201696150]
14 Li X, Sun X, Carmeliet P. Hallmarks of Endothelial Cell Metabolism in Health and Disease.
2019; 30: 414-433[PMID: 31484054 DOI: 10.1016/j.cmet.2019.08.011]
15 Yetkin-Arik B, Vogels IMC, Nowak-Sliwinska P, Weiss A, Houtkooper RH, Van Noorden CJF, Klaassen I, Schlingemann RO. The role of glycolysis and mitochondrial respiration in the formation and functioning of endothelial tip cells during angiogenesis.
2019; 9: 12608 [PMID: 31471554 DOI: 10.1038/s41598-019-48676-2]
16 Caja S, Enríquez JA. Mitochondria in endothelial cells: Sensors and integrators of environmental cues.
2017;12: 821-827 [PMID: 28448943 DOI: 10.1016/j.redox.2017.04.021]
17 Pinti MV, Fink GK, Hathaway QA, Durr AJ, Kunovac A, Hollander JM. Mitochondrial dysfunction in type 2 diabetes mellitus: an organ-based analysis.
2019; 316: E268-E285 [PMID: 30601700 DOI:10.1152/ajpendo.00314.2018]
18 Rellick SL, Hu H, Simpkins JW, Ren X. Evaluation of Bioenergetic Function in Cerebral Vascular Endothelial Cells.
2016 [PMID: 27911398 DOI: 10.3791/54847]
19 Minami T, Muramatsu M, Kume T. Organ/Tissue-Specific Vascular Endothelial Cell Heterogeneity in Health and Disease.
2019; 42: 1609-1619 [PMID: 31582649 DOI: 10.1248/bpb.b19-00531]
20 De Bock K, Georgiadou M, Schoors S, Kuchnio A, Wong BW, Cantelmo AR, Quaegebeur A, Ghesquière B,Cauwenberghs S, Eelen G, Phng LK, Betz I, Tembuyser B, Brepoels K, Welti J, Geudens I, Segura I, Cruys B, Bifari F,Decimo I, Blanco R, Wyns S, Vangindertael J, Rocha S, Collins RT, Munck S, Daelemans D, Imamura H, Devlieger R,Rider M, Van Veldhoven PP, Schuit F, Bartrons R, Hofkens J, Fraisl P, Telang S, Deberardinis RJ, Schoonjans L, Vinckier S, Chesney J, Gerhardt H, Dewerchin M, Carmeliet P. Role of PFKFB3-driven glycolysis in vessel sprouting.
2013;154: 651-663 [PMID: 23911327 DOI: 10.1016/j.cell.2013.06.037]
21 Nickel A, Kohlhaas M, Maack C. Mitochondrial reactive oxygen species production and elimination.
2014; 73: 26-33 [PMID: 24657720 DOI: 10.1016/j.yjmcc.2014.03.011]
22 Lyra-Leite DM, Andres AM, Cho N, Petersen AP, Ariyasinghe NR, Kim SS, Gottlieb RA, McCain ML. Matrix-guided control of mitochondrial function in cardiac myocytes.
2019; 97: 281-295 [PMID: 31401347 DOI:10.1016/j.actbio.2019.08.007]
23 Vandekeere S, Dubois C, Kalucka J, Sullivan MR, García-Caballero M, Goveia J, Chen R, Diehl FF, Bar-Lev L, Souffreau J, Pircher A, Kumar S, Vinckier S, Hirabayashi Y, Furuya S, Schoonjans L, Eelen G, Ghesquière B, Keshet E, Li X, Vander Heiden MG, Dewerchin M, Carmeliet P. Serine Synthesis
PHGDH Is Essential for Heme Production in Endothelial Cells.
2018; 28: 573-587.e13 [PMID: 30017355 DOI: 10.1016/j.cmet.2018.06.009]
24 Kalucka J, Bierhansl L, Conchinha NV, Missiaen R, Elia I, Brüning U, Scheinok S, Treps L, Cantelmo AR, Dubois C, de Zeeuw P, Goveia J, Zecchin A, Taverna F, Morales-Rodriguez F, Brajic A, Conradi LC, Schoors S, Harjes U, Vriens K,Pilz GA, Chen R, Cubbon R, Thienpont B, Cruys B, Wong BW, Ghesquière B, Dewerchin M, De Bock K, Sagaert X,Jessberger S, Jones EAV, Gallez B, Lambrechts D, Mazzone M, Eelen G, Li X, Fendt SM, Carmeliet P. Quiescent Endothelial Cells Upregulate Fatty Acid β-Oxidation for Vasculoprotection
Redox Homeostasis.
2018; 28:881-894.e13 [PMID: 30146488 DOI: 10.1016/j.cmet.2018.07.016]
25 Carmeliet P, Jain RK. Molecular mechanisms and clinical applications of angiogenesis.
2011; 473: 298-307[PMID: 21593862 DOI: 10.1038/nature10144]
26 Zhuo W, Song X, Zhou H, Luo Y. Arginine deiminase modulates endothelial tip cells
excessive synthesis of reactive oxygen species.
2011; 39: 1376-1381, suppl 2 p following 1382 [PMID: 21936818 DOI:10.1042/BST0391376]
27 Pircher A, Treps L, Bodrug N, Carmeliet P. Endothelial cell metabolism: A novel player in atherosclerosis?
2016; 253: 247-257 [PMID: 27594537 DOI: 10.1016/j.atherosclerosis.2016.08.011]
28 Kadlec AO, Beyer AM, Ait-Aissa K, Gutterman DD. Mitochondrial signaling in the vascular endothelium: beyond reactive oxygen species.
2016; 111: 26 [PMID: 26992928 DOI: 10.1007/s00395-016-0546-5]
杂志排行

World Journal of Diabetes的其它文章
- Different nutrient compositions in diet and taking hypoglycemic drugs can modulate gut microbial flora
- Mapping the global research landscape on insulin resistance:Visualization and bibliometric analysis
- Relationship between age of pregnant women with gestational diabetes mellitus and mode of delivery and neonatal Apgar score
- Hyperglycemia and reduced adiposity of streptozotocin-induced diabetic mice are not alleviated by oral benzylamine supplementation
- Effectiveness and safety of COVID-19 vaccines in patients with diabetes as a factor for vaccine hesitancy
- COVID-19 associated diabetes mellitus: A review