基于50K SNP 芯片信息的枣庄黑盖猪群体遗传结构分析
2022-09-14王余北肖莲梅何帅涵孙厚伟朱绍伟李肖产舒恒郭雷李久庆方美英
王余北,肖莲梅,何帅涵,孙厚伟,朱绍伟,李肖,产舒恒,郭雷,李久庆,方美英*
(1.中国农业大学动物科学技术学院,北京 100193;2.山东福藤食品有限公司,山东滕州 277599;3.滕州市畜牧渔业事业发展中心,山东滕州 277599;4.枣庄市畜牧渔业事业发展中心,山东枣庄 277000)
枣庄黑盖猪是我国优良的华北型地方猪种,蕴含独特的种质基因,是我国宝贵的遗传资源。但随着外来商业猪种的引入,枣庄黑盖猪的养殖数量越来越少,以至于到90 年代末,枣庄黑盖猪只零星散见于山东部分偏僻村庄中。2004 年在山东省相关部门、企业与科研单位的支持下,山东福藤食品有限公司着手开展枣庄黑盖猪的保种工作,对其进行了抢救性保护与提纯复壮。但在开展实际工作的过程中遇到了2 个方面问题:一方面,由于枣庄黑盖猪最初的核心群主要从一些散在农户手中购买,具体遗传背景未知,给配种计划的安排带来不少困难;另一方面,传统的畜禽遗传学研究中,遗传参数评估主要依赖于育种过程中的表型数据与系谱记录,而枣庄黑盖猪育种工作开展时间短,再加上实际生产过程中常发生系谱记录与表型数据错误、缺失等情况,难以通过传统的遗传学方法对枣庄黑盖猪群体进行遗传参数评估。因此,从分子层面研究枣庄黑盖猪的群体遗传结构,明确群体遗传背景,对于枣庄黑盖猪遗传资源的保护和发掘利用具有重要意义。
近年来随着高通量测序技术的发展和成本的降低,在猪、鸡等畜禽中均推出高效的商业化SNP 芯片。SNP 芯片具有密度高、分型准确性高、可自动化等优势,在医药、生物、农业等领域的全基因组研究中被充分利用。目前利用SNP 标记分析群体遗传结构的生物信息学方法较为成熟,使得分析未知遗传背景群体的遗传结构成为了可能。本研究使用猪全基因组SNP 芯片对枣庄黑盖猪核心群体进行遗传结构分析,为群体的种质资源保护与发掘利用工作提供研究基础。
1 材料与方法
1.1 实验材料 实验样本采集于山东省福藤食品有限公司枣庄黑盖猪原种猪场,所用芯片为纽勤生物科技(上海)有限公司的50K SNP 芯片。本研究共采集200 头枣庄黑盖猪核心群,其中公猪12 头,母猪188 头。采样部位为耳组织,用消毒的耳号钳剪下少许耳组织放入5 mL 离心管中后,迅速放入-20℃冰箱中保存备用。
1.2 基因型测定 所有组织样品均浸泡在75% 的酒精中,经冲洗晾干,进行DNA 提取并质控后,使用NEOGEN GGP-Porcine 50K 芯片进行基因分型。
1.3 SNP 质量控制 使用PLINKv1.90 软件对检测到的SNP 进行质控,质控条件为:①删除检出率低于90% 的SNP 标记(PLINK 参数:--geno 0.1);②删除基因型缺失率高于10% 的个体(PLINK 参数:--mind 0.1);③删除次等位基因频率(Minor Allele Frequency,MAF)小 于5% 的SNP标记(PLINK 参数:--maf 0.05);④删除性染色体上的位点(PLINK参数:--chr 0-18)。后续使用质控后的SNP 进行分析。

2 结果
2.1 SNP 质量控制 本研究使用NEOGEN 公司的GGP-Porcine 50K 芯片对枣庄黑盖猪基因组SNP 进行基因分型,芯片上共检测到50 697 个SNP 位点,删除位于性染色体上的位点后剩余48 228 个位点,其中检出率低于90%的SNP 标记有367 个,次等位基因频率小于5% 的SNP 标记有3 691 个,整体质控后共剩余44 170 个SNP 标记用于后续分析。
2.2 群体遗传结构与相关遗传参数分析 为研究枣庄黑盖猪的群体遗传结构,首先使用质控后的200 头枣庄黑盖猪群体的SNP 数据进行系统发生树分析,图1 为使用最大似然法(ML)构建的系统发生树,图中最外圈个体名带黑点为公猪个体,其余为母猪个体。结合系统发生树和KING 软件对公猪血统的推断结果,将枣庄黑盖猪12 头公猪确定为6 个血统,各血统内公猪信息如表1 所示。
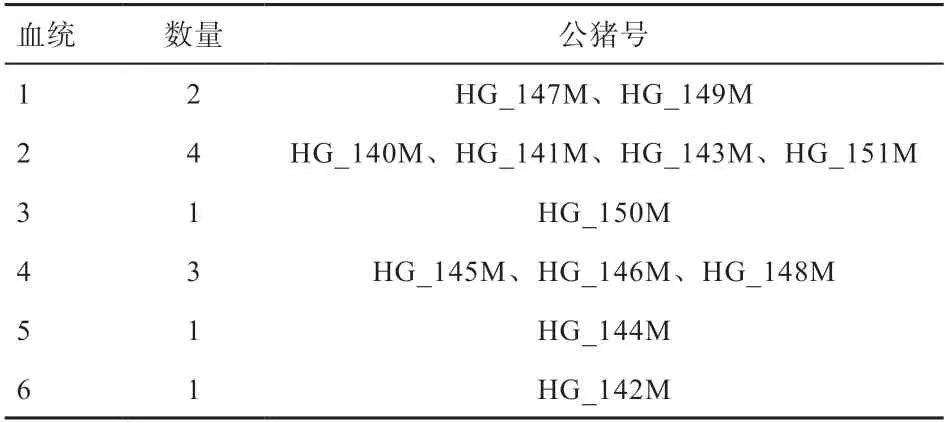
表1 枣庄黑盖猪公猪血统表
结合公猪血统信息与系统发生树(图1)结果,将188 头母猪划分进6 个公猪血统中,形成6 个母猪家系,图中最外圈分别用数字1~6 来表示各血统/家系。图2为结合血统与家系信息使用PLINK 软件进行的群体主成分分析,并使用R 绘制第一主成分和第二主成分的分析图,其中不同血统/家系个体用不同的形状代表,主成分分析中可看出枣庄黑盖猪群体虽然在图中的分布并不均匀,但大部分同血统/家系内的样本聚在一起,说明同血统/家系内的个体关系更近,血统与家系的划分较合适。
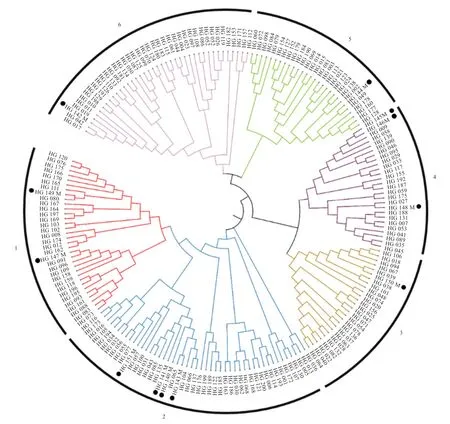
图1 枣庄黑盖猪群体系统发生树

图2 枣庄盖猪各血统/家系主成分分析(PCA)
为了对群体中的亲缘关系进行分析,研究使用群体基因组血缘同源(IBD)信息进行分析,群体内200 个个体共产生19 900 个关系对,其分析结果表明群体平均亲缘系数为0.089 0±0.109 6,其中90.95%的个体对间亲缘关系较远(亲缘系数<0.25),7.89% 的个体对间亲缘关系较近(亲缘系数0.25~0.50),其余1.16%的个体对间亲缘关系很近(亲缘系数>0.25)。各家系的平均亲缘系数如表2 所示。
研究使用PLINK 软件与R 包detectRUNS 对枣庄黑盖猪群体的杂合度与近交系数等遗传参数进行分析,分析结果表明枣庄黑盖猪群体的平均观测杂合率为0.389 0,平均期望杂合率为0.378 9,平均近交系数为0.111 7±0.072 3。其中各家系平均近交系数如表2 所示。
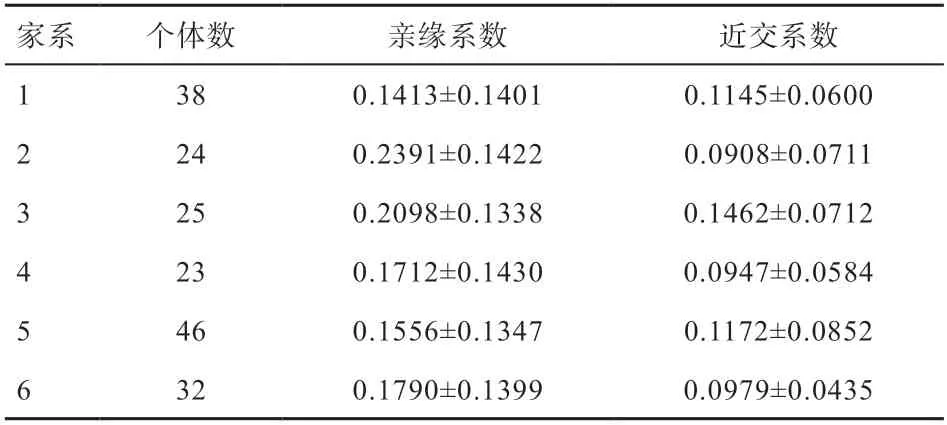
表2 枣庄黑盖猪不同家系亲缘系数与近交系数
3 讨 论
本研究使用枣庄黑盖猪的基因组信息对其遗传结构进行了系统发生树、主成分分析和群体IBD 等系统分析。系统发生树是一种被认为具有共同祖先的各物种间演化关系的树状图,是一种亲缘分支分类方法。本研究中由系统发生树结果可以看出枣庄黑盖猪群体在系统进化树上聚合成几个独立的大分支,根据系统发生树结果和群体亲缘关系推断结果将枣庄黑盖猪公猪群体分为6 个血统,并进一步将母猪群体划分进这6 个公猪血统中。根据血统与家系信息,进一步对枣庄黑盖猪群体进行了主成分分析,主成分分析中枣庄黑盖猪群体虽然分布并不均匀,但大多数同血统/家系内的样本聚在一起,说明同血统/家系内的个体关系更近,证明研究对枣庄黑盖猪群体血统与家系的划分较合适,可为后续育种工作提供研究基础,而有些样本在图中分布较散可能是在其他主成分上密集分布。本研究使用群体IBD 信息进行了枣庄黑盖猪的亲缘系数估计,与王继英等人在2019 年分析的结果相比,本研究中枣庄黑盖猪群体的平均亲缘系数有一定程度的降低,说明近几年枣庄黑盖猪群体在配种时已注意选用亲缘关系较远的个体进行交配,避免了近交衰退的发生。综上所述,研究发现虽然枣庄黑盖猪群体的亲缘系数不高,处于较低水平,且与几年前相比亲缘系数已有所下降,但家系内的亲缘系数仍相对较高,因此在后续选配工作中应继续尽量避免同一血统公猪与同一家系母猪个体交配,以避免群体近交系数过快增长,造成群体近交衰退的发生。
群体杂合度是评估一个群体遗传多样性的关键指标,观测杂合度是指在这个群体内实际观测到的杂合个体数占样本总体的比例,期望杂合度是指样本中任意一个个体某一位点杂合的期望概率,群体杂合度可以反映一个群体的遗传多样性,本研究中枣庄黑盖猪群体的平均观测杂合度为0.389 0,平均期望杂合度为0.378 9,群体平均观测杂合度高于平均期望杂合度,一方面说明枣庄黑盖猪品种在形成过程中可能在一定程度上混入了其他品种猪的血统,另一方面也说明枣庄黑盖猪群体内遗传多样性较高,具有较高的纯化选育潜力。
近交是指由较近亲缘关系个体进行交配,而近交程度的升高会使基因组上有害基因纯合子增多,产生近交衰退而降低生产性能。目前有研究表明根据ROH片段可以对个体的近交系数进行有效评估,且基于ROH 计算的近交系数是更准确评估个体近交水平的一种方法。本研究中使用ROH 计算了枣庄黑盖猪群体的近交系数,群体平均近交系数为0.111 7±0.072 3,与其他同样使用ROH 来评估近交系数的猪种相比,枣庄黑盖猪的近交系数低于梅山猪、沙乌头猪、法系大白猪、加系大白猪,略高于枫泾猪。整体来看,枣庄黑盖猪群体的近交系数处于较低水平,说明近些年该群体存在较低程度的近交,表明在枣庄黑盖猪的选育工作中未进行近交改良或较大程度的人工干预,群体遗传多样性良好,具有后续进行分子选育的良好基础。
4 结 论
本研究基于猪50K SNP 芯片信息对枣庄黑盖猪200 头核心群体的遗传结构进行了分析。研究表明枣庄黑盖猪公猪群体可分为6 个血统,群体亲缘系数与近交系数均处于较低水平,群体的平均观测杂合度高于平均期望杂合度,具有较高的遗传多样性,但家系内亲缘系数要高于群体亲缘系数,在选配时应尽量避免同一血统公猪与同一家系母猪交配。本研究从分子层面揭示了枣庄黑盖猪的种群遗传结构和遗传多样性,可为后续的种群保种及选育利用工作提供基础。
