应用Maxent模型对我国珍稀植物刺楸分布预测及其保护空缺分析
2022-09-08王文波胡理乐布艾佳尔闫伯前
王文波 胡理乐 布艾佳尔 闫伯前
(国家林业和草原局林草调查规划院,北京,100714) (新疆维吾尔自治区林业规划院) (北京农学院)
环境决定了物种的地理分布,例如许多研究认为,随着气候变化,物种的分布也随之发生变化[1-3],特别是位于分布边缘区的种群往往对气候变化响应的敏感性更高,其变动幅度也会更明显[4]。对于珍稀物种,其分布范围也是保护的基础信息。在大尺度上,为了获取影响物种分布的主导环境因子并预估未来环境因子变化对物种分布的影响,相关学者逐渐开始使用物种分布模型(SDM)这一研究手段[5]。SDM又被称为生态位模型(NM)或生境适宜模型,其本质是通过物种分布数据结合物种分布区域的环境因子数据映射出符合物种生存需求的潜在生境,并以此预测研究区域内物种未来的潜在分布[6]。常用的SDM分为两类,即机理性模型和概率性模型(如GARP、ENFA、bioclim、Domain等)。最大熵模型(Maxent)也属于概率性模型,与机理性模型相比,其系统相对开放,模型结构相对简单,所需参数也较少;而与其他常见的相关性模型相比,其预测的结果便于分析[7],准确度也相对较高[8-10],尤其分析广域分布且位置信息不确定的物种时具有很大的优势[11],因此被广泛应用于生物多样性评估、自然保护地设计、入侵物种管理和物种迁地保护等方面[12-14]。
刺楸(Kalopanaxseptemlobus)是五加科刺楸属单种属的落叶乔木,自然分布在我国东北、华北、长江流域等地,日本、朝鲜半岛和西伯利亚也有分布[15]。多生于阳性森林、灌木林中和林缘,水湿丰富、腐殖质较多的密林,向阳山坡,甚至岩质山地也能生长[16]。集药用、材用、园林绿化、食用等为一体,是我国重要的用材、园林观赏、药食兼用树种[17]。刺楸自然分布范围虽然较为广泛,但由于环境开发使其生境遭受严重破坏,加之过度采伐,刺楸野生种群数量急剧下降,多为零散分布,现北京、山东等地区已把刺楸列入珍稀濒危物种名录[18]。目前刺楸相关研究主要集中在其生长规律、种群生态位特征、影响生长因素、保护现状及种子催芽育苗等方面[19-22]。但在我国,有关刺楸种质资源的自然分布、群体结构、遗传变异及其保护成效评估等研究仍然较少,刺楸种质资源的保护仍不到位。在气候变化和人类活动加剧的大背景下,如何对刺楸进行保护规划成为刺楸种质资源保护及可持续利用过程中亟待解决的问题。为预测刺楸高适生区的潜在分布并提出有效的保护对策,本研究将采用Maxent模型结合ArcGIS软件对刺楸进行适宜分布区的划分,根据百分比贡献率、jackknife检验和环境限制因子探讨具体的环境因子对刺楸分布的影响,探究刺楸的生态位需求;并通过对保护地边界数据的叠加分析,获取刺楸适生区保护空缺范围,为防止刺楸种群的持续退化提供坚实的理论依据。
1 研究方法
刺楸地理分布数据基于查阅文献、检索中国植物图像库(ppbc.iplant.cn)和全球生物多样性信息平台(Global biodiversity Information Facility)、中国数字植物标本馆(https:www.cvh.ac.cn/)和中国自然保护地标本共享平台(http:www.papc.cn/html/folder/1-1.htm)(标本数据自1970年以后)等数据库选取刺楸野生分布地及其标本采集地信息共203个。筛选出有具体经纬度坐标或能精确至小地名(至少为乡镇)的点位,通过GPS确定其经纬度坐标,去除经纬度重复的数据,最终整理得到159条经纬度精确的刺楸分布点信息(图1)。
图1 本研究刺楸的分布位点
环境因子主要包括地形因子和气候因子两个方面。通过中国科学院国际科学数据服务平台(http:www.csdb.cn)获取了3个地形因子(海拔、坡向、坡度),通过世界数据库World-Clim(http:www.worldclim.org/)[23]获取了19个常用生物气候因子(bio1-bio19)[24]。上述数据的空间分辨率为1 km×1 km。
中国国界和各省行政区划矢量图源自全国地理信息资源目录服务系统(http:www.webmap.cn/),保护地边界矢量图来自中国科学院地理科学与资源研究所资源环境数据中心。
由于气候因子主要包括降水及温度两类,各气候因子之间存在一定的相关性,在进行潜在分布地模拟之前需对各环境因子进行相关性分析[25]。气候因子和地形因子之间多重共线性分析检验图层的相关性通过SPSS软件实现,若2个环境因子的相关性绝对值>0.8,则去除其中一个。最终选取11个环境因子,包括8个气候因子(bio1,bio2,bio3,bio5,bio7,bio12,bio13,bio15)和3个地形因子(alt,asp,slope)用于构建模型。
分别将ASCII格式的环境因子和csv格式的刺楸分布点经纬度信息导入Maxent 3.3.3中。取其中25%的分布点作为测试集,余下75%分布点作为训练集,重复运行100次预测,并通过AUC值来评价预测结果的精确度,AUC值为ROC曲线(受试者工作特征曲线)与横轴围成的面积[26]。该值越接近于1,则环境变量与预测的物种地理分布区之间的相关性越大,即模型预测结果越准确[27]。当AUC值在0.50~0.60时,代表预测精度极低;在0.60~0.70时,代表预测精度较差;在0.70~0.80时,代表预测精度一般;在0.80~0.90时,代表预测精度良好;在0.90~1.00时,代表预测精度非常好。同时,通过Jacknife刀切图来评价各环境因子在模拟过程中的贡献权重。
将预测结果导入ArcGIS中,输出为栅格格式,最终得到刺楸的潜在分布图。使用适生指数P形成刺楸适宜性综合评价等级,利用等分法对刺楸的潜在分布图进行重分类[28],具体是P<0.2,为不适宜区;0.2≤P<0.4,为低度适宜区;0.4≤P<0.6,为中度适宜区;0.6≤P<0.8为高度适宜区,P≥0.8为极高度适宜区。
2 结果与分析
2.1 刺楸潜在空间分布的影响因子
本研究中模型AUC值为0.912,说明模型预测可信度非常高。
基于159个刺楸的有效分布点,利用于Maxent模型预测的11个环境因子中,由刀切法检验结果可知(图2),仅使用单一环境因子时,对正规化训练增益影响最大的5个依次为最湿润月份降水(bio13)、全年降水量(bio12)、平均气温日较差(bio2)、年均温变化范围(bio7)和年平均气温(bio1);贡献率排名前5位的依次为全年降水量(bio12,29.9%)、最湿润月份降水(bio13,19.1%)、平均气温日较差(bio2,11.8%)、坡度(slo,11.2%)和年平均气温(bio1,10.7%),累计贡献率为82.7%。综合来看,影响刺楸潜在地理分布的主要环境因子依次为降水因子(降水量)、气温因子(日温差和年均温)和地形因子(坡度)。
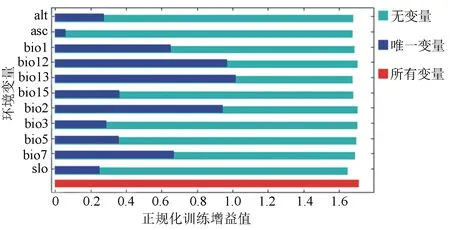
图2 刺楸环境刀切法检验结果
Maxent模型运行结果中的10%训练存在阈值通常被界定为物种存在的阈值,本研究根据环境因子对存在概率曲线的响应来判断刺楸的存在概率与环境因子之间的关系[29]。本研究选取影响力前3的环境变量进行阐述(图3),当年降水量(bio12)达到510 mm后,刺楸开始存在,随着降水的继续增加,刺楸的存在概率呈现上升趋势,直至在1 100 mm时达到峰值,其中年降水量1 050~1 490 mm时为其高适生范围;而最湿润月份降水量(bio13)大于135 mm时,刺楸开始存在,随着最湿润月份降水量的增加,刺楸的存在概率呈现上升趋势,最湿润月份降水量达到250 mm时达到峰值,其中最湿润月份降水量200~260 mm时为其高适生范围;在模型采集到的日温差(bio2)范围内,刺楸存在的概率随着日温差的增加基本呈单调下降的趋势,其中高适生范围为4~12 ℃。
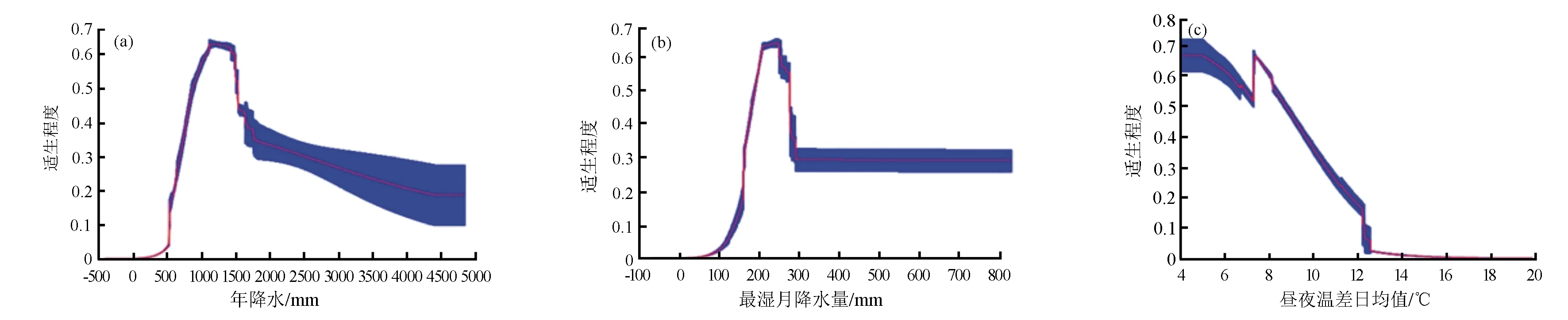
(a)年降水量;(b)最湿润月份降水量;(c)平均日气温差。图3 刺楸对主要的3个环境变量的响应曲线
2.2 刺楸的潜在空间分布
利用Maxent模型运算模拟出刺楸在现代气候条件下的潜在地理分布区(图4)。刺楸的高适宜区生境面积为24.4万km2,约占中国国土总面积的2.5%。刺楸的潜在高适宜生境区域,主要集中分布于辽东半岛山地-长白山南段、燕山山脉、鲁中山地和鲁南丘陵、太行山东南角、伏牛山地-大别山地、皖南-浙西山地、藏东南湿润性的低海拔河谷区、大巴山、岷山、邛崃山东坡、大小相岭、鄂西-川东山地、乌蒙山至武陵山脉、罗霄山脉以及桂西岑王老山-南岭一带。本研究将112°E作为刺楸潜在高适生分布区的东、西部分界线,发现两个区域潜在高适生分布区的空间格局具有一定差异,东部地区存在明显的地理隔离,尤其在东北、华北、山东以及江南一带较为明显,而在112°E经线以西,除了藏东南种群外,刺楸的潜在高适生分布区则基本连接成片。
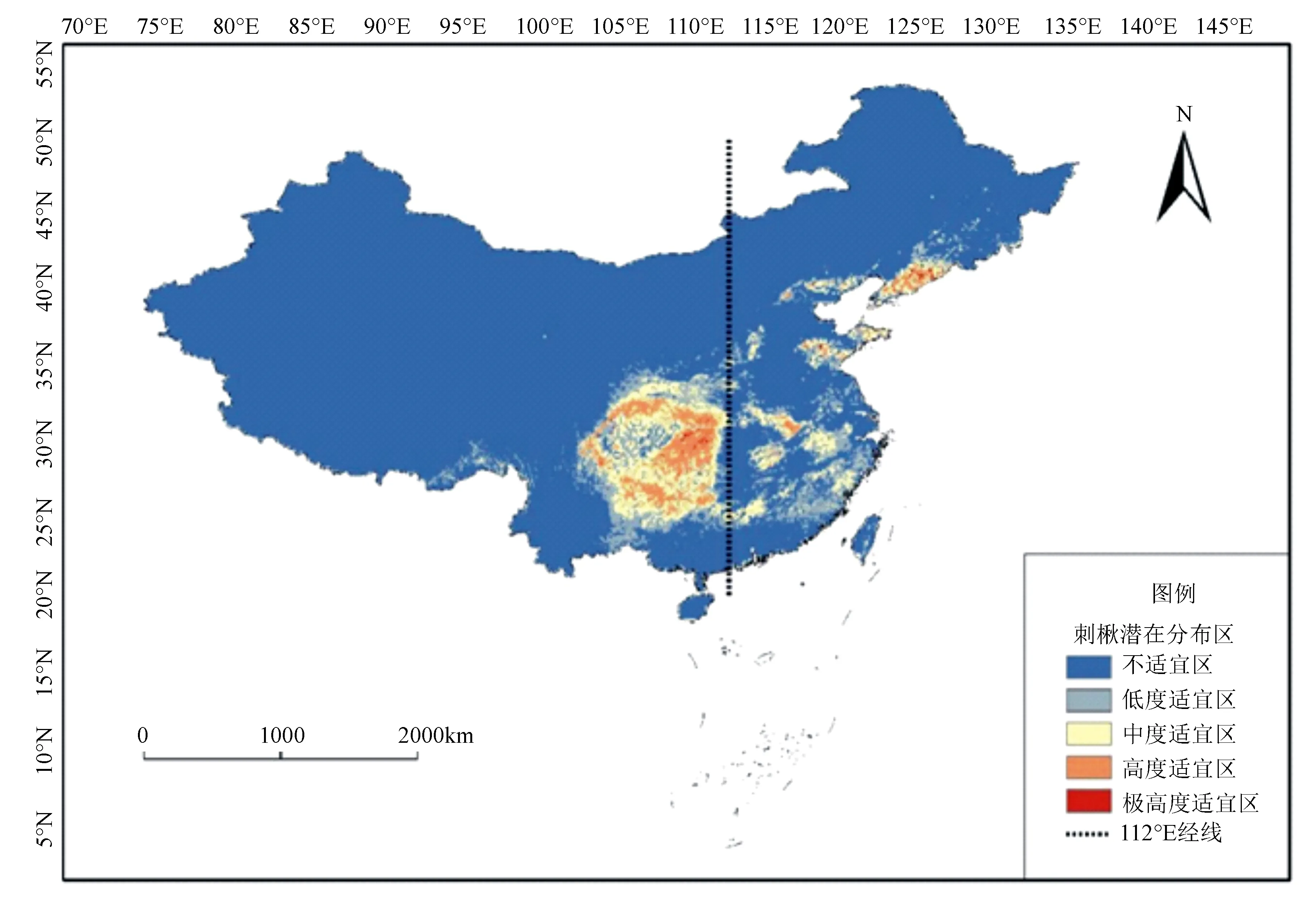
图4 刺楸在中国的潜在分布预测
2.3 刺楸保护空缺
模型模拟结果表明,当前刺楸潜在高适生区主要位于我国暖温带、北亚热带和中亚热带相对湿润的山地。本研究将保护地边界及刺楸潜在分布极高适生区进行叠加发现,刺楸极高适生区生境面积(热点区域)为9 126.8 km2,而全国仅有12.5%(1 141.7 km2)的热点区域位于各级自然保护地内,其中东部地区仅保护了6.3%(210.5 km2)的热点区域,西部地区保护了16.0%(931.2 km2)的热点区域(图5和表1)。相对而言,西部地区现有的保护地对刺楸潜在极高适生区的保护略好,而东部地区对刺楸的保护目前存在大量空缺。
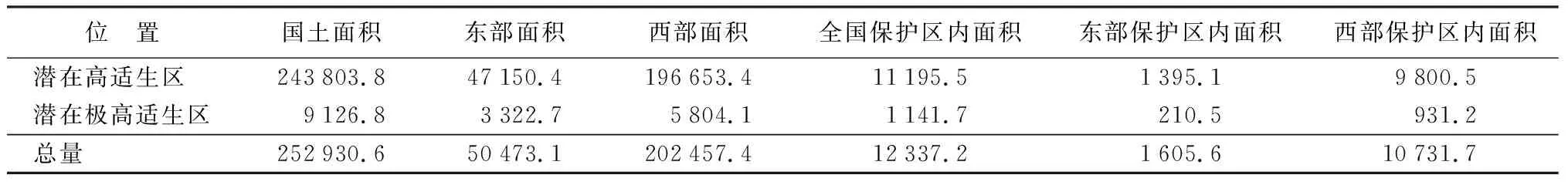
表1 刺楸潜在高适生区和潜在极高适生区的面积 km2
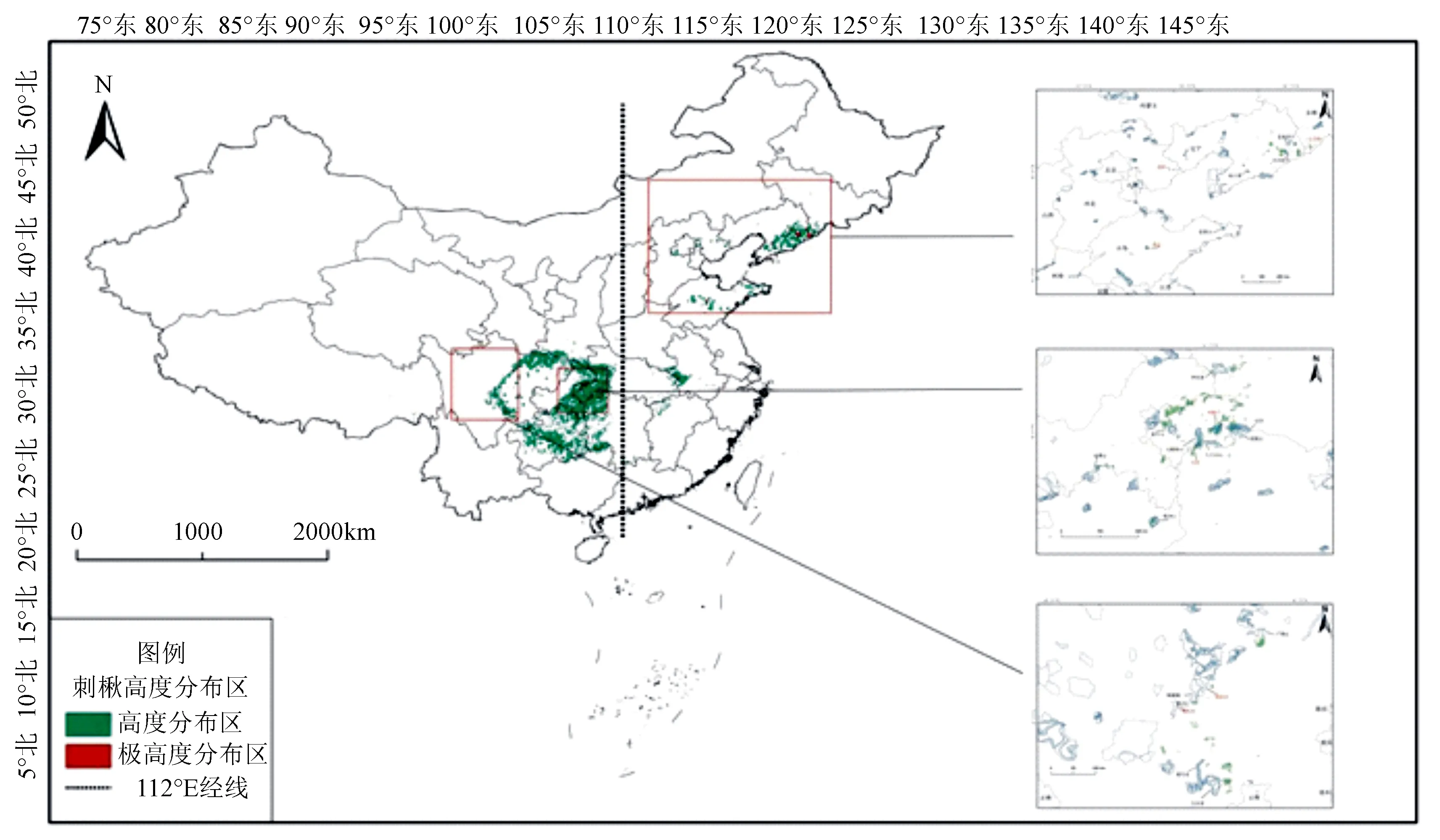
图5 刺楸在中国的保护空缺分析
3 讨论与结论
刺楸在我国分布面积广大,通过Maxent刀切法分析发现,影响其分布的气候因子主要是水分,其次是温度,刺楸对地形因子的响应相对不敏感。刺楸叶片为纸质,相对较薄,导致其不耐旱,例如在我国东北地区,广布树种色木槭、千金榆、紫椴、蒙古栎、加杨的耐旱性高于刺楸[30],并且刺楸的抗寒性弱于榆、黄檗、紫椴、水曲柳等广布物种[31]。在我国温带、亚热带森林主要受到季风气候的影响,季风带来的大量降水保证了生长季刺楸对水分的需求。刺楸对温度的适应幅度相对较宽,这也是其分布广泛的主要原因,其分布北缘的限制因子可能是低温,长期低温导致生长季偏短[32]。但Kang et al.[33]使用Maxent模型发现温度和坡向是影响韩国刺楸分布的主导环境因子,这可能是因为韩国地域相对狭小,山地垂直气候更多地影响气温变化,而坡向同时影响积温和降水。虽然刺楸落叶的特性有助于抵御冬季的严寒,但在我国南方常绿阔叶林中,这一特性可能导致其竞争性相对较弱,难以成为群落中的优势物种。同时在南亚热带难以寻觅到刺楸,有可能是刺楸的种子具有长休眠现象,其休眠因素主要是种皮不透水,种子的胚发育不全,并存在抑制物,在人工萌发的处理中需要低温层积[19],而华南地区的低温难以满足刺楸种子成熟过程。
虽然在我国刺楸的分布面积广大,但该物种却存在于多地的保护植物名录中。过去以其散生不集中的状态难以为其单独设立保护区,因此目前全国仅有一个以保护刺楸物种为目的的保护地,即于1983年设立的、现为市州级保护区的吉林省白山市八道江区三道沟刺楸林保护区,位于本研究中我国刺楸潜在分布的东北角。但在全国范围内,本研究发现仅有少量潜在极高适生热点(12.5%)位于自然保护地内,同时多数自然保护地并未将其视为主要保护对象,这不利于刺楸种群的恢复。根据现有的模型结果,本研究认为东部地区的刺楸种群亟待加强保护,特别是辽东山地的保护地空缺十分明显(如白石砬子国家级自然保护区、老秃顶子国家级自然保护区、仙人洞国家级自然保护区、文字岭县级保护区等保护区等周边地区),需要调整扩大保护地范围或设立保护小区,而东部其余地区人口压力较大,例如燕山山脉(老岭市级自然保护区、十渡风景区等)、鲁中南山地丘陵(昆嵛山国家级自然保护区、鲁山省级自然保护区等)和大别山区(天马国家级自然保护区等),可能不适合扩大保护地范围,因此需要加强保护的监管力度;相对而言,整体上西部山地人口密度较小,可以适当扩大保护区面积或保护小区以更好地囊括刺楸潜在极高适生区,如鄂西-武陵山脉(神农架国家级自然保护区、后河国家级自然保护区、星斗山国家级自然保护区、七姊妹山国家级自然保护区、壶瓶山国家级自然保护区、八大公山国家级自然保护区、梵净山国家级自然保护区,金佛山国家级自然保护区、木林子省级自然保护区、天子山省级自然保护区和大风堡省级自然保护区等保护区和周边地区)、岷山-大小相岭(千佛山国家级自然保护区、蜂桶寨国家级自然保护区、大风顶国家级自然保护区和喇叭河省级自然保护区等保护区和周边地区)以及西藏东南部(慈巴沟国家级自然保护区和巴结省级自然保护区的周边地区)(图5)。
现有的Maxent模型仅包含气候和地形因子,缺乏土壤因子的数据,与此同时刺楸的分布信息也存在更新不及时和调查遗漏的现象,此外,人类活动的干扰对刺楸现实分布的影响难以具体量化,因此,本研究依然存在着一些不确定性。另外,基于刺楸的生物学研究显示刺楸成熟晚,自花授粉不良[34]。在遗传学方面,目前我国仅有少数地区针对刺楸的遗传结构进行了分析[35],在全国尺度上,刺楸的种内遗传多样性是否成为刺楸濒危的重要因素还需要更多数据进行支撑。对于上述的不确定因素,未来需要继续加强对刺楸种群的研究,尤其是分析东部地理分隔种群的遗传差异及基因流,在微观角度上寻找刺楸分布格局变化的原因,同时从生理生态方面探究刺楸竞争能力和分布受限的具体机制,并且还需要进行刺楸高适生区环境因子的长期监测,以期获得更高质量的建模参数。
