大鼠延髓头端腹内侧区内G蛋白偶联雌激素受体与多种神经元的免疫荧光共定位关系
2022-09-07戎伟芳
孟 立 戎伟芳
(上海交通大学医学院基础医学院,上海 200025)
痛觉信息传递和阿片类药物镇痛存在明显的性别差异,女性对某些以疼痛为主要症状的疾病较男性更易感[1-2]。雌激素被认为可能参与痛觉的调控,但具体机制不清楚。G蛋白偶联雌激素受体(G protein-coupled estrogen receptor,GPER)能够介导雌激素的快速非基因组效应[3-4],有研究表明其在痛觉传导通路中发挥作用[5-7]。延髓头端腹内侧区(rostral ventromedial medulla, RVM)由中缝大核(nucleus raphe magnus, NRM)、网状巨细胞核(nucleus reticularis gigantocellularis, Gi)及邻近的网状结构构成[8-9],在痛觉的下行调控和阿片类药物镇痛中起重要作用[10-11],但其神经元构成和局部环路尚不清楚。本研究对大鼠的RVM区域GPER和5-羟色胺(5-hydroxytryptamine,5-HT)能神经元标志物色氨酸羟化酶(tryptophan hydroxylase,TPH)、儿茶酚胺能神经元标志物酪氨酸羟化酶(tyrosine hydroxylase, TH)、神经元型一氧化氮合 酶(neuronal nitric oxide synthase, nNOS)、胆碱能神经元标志物胆碱乙酰转移酶(choline acetyltransferase, ChAT)以及脑啡肽(enkephalin,ENK)进行双重免疫组织化学显色,旨在探究GPER在RVM的表达特征以及与上述多种神经元的共定位关系,从而了解雌激素影响痛觉下行调控通路和阿片镇痛效应可能的结构和分子基础。
1 材料和方法
1.1 实验动物
6周龄雄性SD大鼠,由上海交通大学医学院实验动物科学部提供。动物饲养室内保持25°C恒温,光照时间为早8∶00点至晚8∶00点,通风良好,水食充足。实验操作均符合上海交通大学动物伦理相关规定。
1.2 主要试剂
兔 源GRER抗 体(LifeSpan BioSciences Inc.,LS-A4272);山羊源ChAT抗体(Millipore, AB144P);鼠源Enkephalin抗体(Abcam, ab150346);山羊源nNOS抗体(Abcam, ab1376);鼠源TH抗体(Immunostar,22941);鼠源TPH抗体(Sigma, T0678);驴抗兔Alexa 488(Invitrogen, A21206);驴抗羊Alexa 568(Invitrogen,A11057);羊抗兔 Alexa 568(Invitrogen, A11036);羊抗 鼠Alexa 488(Invitrogen, A11029);羊抗鼠Alexa 568(Invitrogen, A11031);羊抗兔 Alexa 488(Invitrogen,A11034)。
1.3 免疫荧光组织化学显色检测RVM内GPER与多种神经元的共定位关系
以60 mg/kg的剂量腹腔注射戊巴比妥麻醉大鼠,将动物固定在解剖板上,剪开胸腔暴露心,将连接灌流泵的注射针头从心尖刺入,灌注生理盐水的同时剪开右心耳。血液冲洗干净后,灌注预冷的4%多聚甲醛溶液。灌流结束后,取出脑组织,置于4%多聚甲醛溶液中后固定过夜。更换预冷的30%蔗糖,放置于4°C使组织脱水,当组织沉底后取出组织,切除大脑和小脑等多余组织,水平放置在盛有OCT的方形包埋盒中,放入干冰速冻,-80℃保存。常规冰冻切片,厚度30 µm。24孔板中加入预冷的0.05 mol/L磷酸盐缓冲液(PBS),将切下的脑片依次放入孔内,每孔6片。从RVM核心位置挑选脑片行常规免疫荧光组织化学显色。加入封闭液(0.05 mol/L PBS+10% 羊血清/驴血清+1%Triton X-100),每孔400 µL,放置在摇床上慢摇,室温1 h。弃封闭液,加入含有相应浓度的一抗抗体稀释液(表 1),每孔300 µL,放置在摇床上慢摇,室温孵育2 h后置于4°C过夜。加入相应的二抗和DAPI(1∶1 000)稀释液,放置在摇床上避光慢摇2 h。将每孔组织切片转移到盛有ddH2O的6 cm培养皿中,用毛刷将切片铺展在载玻片上。待脑片晾干后,加适量封片剂,盖上盖玻片,使用荧光显微镜拍片或放入4°C冰箱暂存。

表1 一抗的稀释比及对应的二抗
2 结果
2.1 RVM脑区GPER与TPH无共定位
本课题组前期研究了RVM内GPER与5-HT的共定位关系,显示RVM内有丰富的5-HT和GPER免疫活性,而5-HT阳性神经元不表达GPER。GPER与TPH双标记免疫荧光显色结果显示,GPER-ir和TPH-ir神经元在NRM内大量存在,两者交错分布(图1A~C,见封二),高倍镜下可见在GPER-ir胞体周围存在TPH-ir纤维(图1D~F,见封二),说明GPER-ir神经元与5-HT能神经元是2个独立的神经元亚群,但两者之间可能存在突触联系。
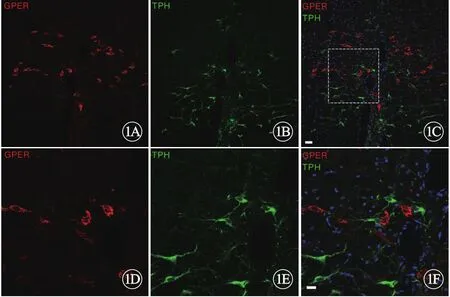
图1 RVM脑区GPER免疫反应阳性神经元不表达TPH。A~C:大鼠RVM脑区GPER与TPH免疫荧光共显色代表图,标尺=40 µm;D~F:大鼠RVM脑区GPER与TPH免疫荧光共显色代表图,标尺=20 µm。红色荧光:GPER;绿色荧光:TPH;蓝色荧光:DAPI.
2.2 RVM脑区GPER与ChAT没有共表达
胆碱能神经元在脑干有大量分布,这些神经元向其他脑区发出投射纤维,组成脑干胆碱能系统。大鼠的RVM脑区组织GPER和ChAT免疫荧光显色结果显示,ChAT在RVM的核心区域NRM没有明显的阳性表达(图2A~C,见封二),ChATir神经元主要分布在外侧巨细胞旁核(nucleus paragigantocellularis lateralis, LPGi)(图2D~F,见封二),与GPER没有共定位。

图2 RVM脑区ChAT与GPER无共表达。A~C:WT大鼠NRM脑区GPER与ChAT免疫荧光共显色代表图,标尺=40 µm;D~F:WT大鼠PGi脑区GPER与ChAT免疫荧光共显色代表图,标尺=40 µm。绿色荧光:GPER;红色荧光:ChAT;蓝色荧光:DAPI.
2.3 RVM脑区绝大多数GPER免疫反应阳性神经元不表达nNOS免疫活性
nNOS是一氧化氮合酶的一个亚型,能在短时间内产生NO。NO是一种气态递质,通过与细胞内受体结合触发多种信号通路。大鼠RVM脑区组织GPER和nNOS双重免疫荧光显色结果显示,nNOS-ir神经元主要分布在RVM偏上方的Giα区域,而GPER-ir则主要位于NRM区域。GPER免疫反应强阳性神经元不表达nNOS-ir,但少量GPER免疫反应弱阳性神经元表达nNOS-ir(图3A~G,见封二)。

图3 RVM脑区多数GPER免疫反应阳性神经元不表达nNOS免疫活性。A~C:大鼠RVM脑区GPER与nNOS免疫荧光共显色代表图,标尺=40 µm。D~F:图C中虚线框内区域放大图,红色荧光:nNOS; 绿色荧光:GPER;蓝色荧光:DAPI;D:大鼠RVM脑区GPER-ir神经元与nNOS-ir神经元分布模式图;绿色:GPER,红色:nNOS,黄色:GPER/nNOS。图中箭头所指为GPER免疫反应弱阳性神经元共表达nNOS免疫活性.
2.4 GPER与TH在PGi有少量共表达
为明确GPER-ir神经元是否可能释放儿茶酚胺类递质,利用免疫荧光方法检测RVM脑区GPER与TH的共表达情况。结果显示,RVM核心区NRM内未见TH-ir神经元分布,但可见少量TH-ir纤维(图4A~C,见封二)。TH免疫活性多表达在LPGi区域(图4D~F,见封二),仅在少数TH-ir神经元检测到较弱的GPER免疫活性(图4G~4I,见封二)。

图4 GPER与TH在LPGi有少量共表达。A~C:大鼠NRM脑区GPER与TH免疫荧光共显色代表图,标尺=40 µm;D~F:WT大鼠LPGi脑区GPER与TH免疫荧光共显色代表图,标尺=40 µm;G~I: 图F中虚线框内区域放大图,标尺=20 µm;红色荧光:TH;绿色荧光:GPER;蓝色荧光:DAPI。图中箭头所指为GPER免疫反应弱阳性神经元表达TH免疫活性.
2.5 RVM脑区GPER阳性神经元周围有大量ENK阳性纤维分布
ENK是一种内源性阿片肽,在疼痛调节中发挥重要功能。对ENK与GPER的免疫荧光显色结果显示, RVM内有密集的ENK免疫反应(ENK-ir)阳性纤维,一些ENK-ir纤维紧邻GPER-ir神经元胞体周围(图5A~C,见封二),这提示两者之间可能存在功能上的联系。在RVM内未观察到ENK免疫反应阳性神经元胞体,这可能是因为肽类物质在胞体内合成后通过轴浆运输到末梢释放,使得胞体内ENK的浓度不足以呈现免疫反应阳性形态。
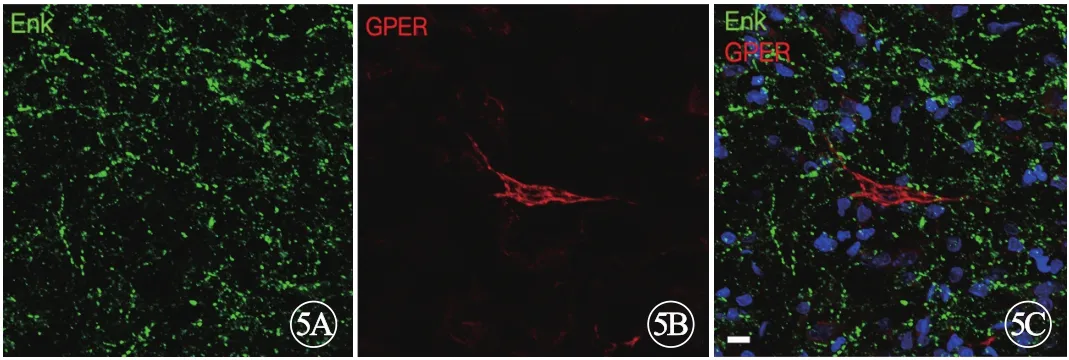
图5 RVM脑区GPER免疫反应阳性神经元不表达ENK免疫活性,标尺=10 µm。A~C:WT大鼠RVM脑区GPER与ENK免疫荧光共显色代表图; 绿色荧光:ENK;红色荧光:GPER;蓝色荧光:DAPI.
3 讨论
脑干下行痛觉调控通路对脊髓背角的感觉信息传递发挥强大的下行调节作用。RVM作为下行调节通路中最后一个功能性核团[12],在痛觉调节中发挥重要作用。RVM脑区内ON细胞响应伤害刺激表现出放电增强,OFF细胞表现放电减少[13],分别促进和抑制伤害性信息传递[14],但目前仍缺乏直接证据。
雌激素参与生殖、神经、内分泌和免疫等系统的调节,与肿瘤、认知功能障碍等多种疾病的发展密切相关[15-16]。然而,无论是在生理还是病理条件下,雌激素随周期变化的特性使得对其疼痛调节作用潜在机制的研究更加困难。RVM内表达芳香化酶,可以局部合成雌激素[17]。课题组前期工作表明,GPER-ir神经元在RVM大量存在,同时表达GABA能神经元标志物GAD67和µ型阿片受体,在在体电生理实验中表现出ON细胞的电活动特征。
5-HT能神经元主要在NRM中分布,其向脊髓的投射是脊髓内5-HT神经末梢的主要来源。研究表明,鞘内注射5-HT可产生明显的镇痛作用[18],而选择性激活5-HT神经元可诱导持续疼痛敏化[19],表明RVM内5-HT系统对疼痛的调节十分复杂。本研究结果显示,GPER-ir神经元与5-HT能神经元交错分布,5-HT能纤维在GPER-ir神经元胞体附近分布。推测GPER-ir神经元与5-HT能神经元之间存在相互作用,影响脊髓内5-HT的释放,以及GPER-ir神经元的活动。
胆碱能神经元在RVM中主要分布在巨细胞旁核(PGi),与文献报道相一致[20],可介导抗伤害感受作用[21]。NO通过与细胞内受体结合触发多种信号通路参与神经系统的重要功能[22]。nNOS在抗伤害感受中发挥重要作用[23]。RVM中有丰富神经元表达nNOS[24]。本研究结果显示GPER的表达有强阳性和弱阳性,这可能是由抗体的非特异性引起,不能排除极少的弱阳性GPER与nNOS、TH共表达,而RVM向脊髓的投射几乎没有nNOS阳性和TH阳性神经元[24],提示可能存在少量GPER阳性神经元在局部发挥作用。
ENK-ir神经元在延髓腹侧和腹外侧的网状结构中有大量分布,并且部分向脊髓投射[25]。RVM内 GABA和ENK双重阳性的神经元可能符合OFF细胞的特征,在下行调控中发挥抑制作用[26]。本研究结果显示RVM区域内有大量ENK-ir纤维分布,一些ENK-ir纤维紧邻GPER-ir神经元胞体,提示GABA和ENK双重阳性的神经元可能通过局部释放ENK作用于GPER-ir神经元上的阿片受体,抑制GPER-ir神经元的活动从而发挥镇痛作用。
综上所述,RVM内GPER特异性表达在非5-HT能神经元,5-HT能纤维以及ENK-ir纤维在GPER免疫反应阳性神经元邻近分布,提示可能有突触联系的存在。笔者将进一步采用电生理方法来探究其功能联系,为研究雌激素调节痛觉和阿片镇痛效应的结构和分子基础提供新思路。
