GABAA受体激动剂通过NF-κB信号通路抑制炎症因子的释放
2022-09-07任晓曦邓邺云刘康瑞张建亮
陈 茜, 任晓曦, 邓邺云, 刘康瑞, 张建亮
(首都医科大学基础医学院神经生物学系, 北京 100069)
帕金森病(Parkinson’s disease,PD)作为常见的神经退行性疾病,其主要的神经病理特征是患者的中脑黑质致密部的多巴胺能神经元的选择性变性与缺失[1]。目前,PD的发病机制仍不清楚,可能的致病因素有线粒体紊乱[2]、氧化应激[3]和蛋白质的错误折叠[4]等。越来越多的证据表明,在PD患者的尸检报告和PD动物模型中,大脑黑质组织中会发生小胶质细胞大量激活,且会伴随着促炎细胞因子水平的增加[5, 6]。进一步的研究证据表明,抑制小胶质细胞激活和炎症因子的释放可以挽救多巴胺能神经元的进行性丢失[7]。然而,如何抑制炎症从而减缓PD的病程仍然是一个未解的问题。
γ-氨基丁酸(γ-aminobutyric acid,GABA)通过与离子型受体GABAA和代谢型受体GABAB结合发挥作用。目前,通过研究表明,GABAA和GABAB受体在中枢神经系统的神经元和胶质细胞上都有表达[8, 9]。给予蝇蕈醇(Muscimol,GABAAR激动剂)可以激活小鼠抗原提呈细胞中的GABAA受体,从而减少细胞中TNF-α、IL-6的升高[10],提示GABA除了众所周知的离子型作用外,GABA还能发挥代谢型作用[11]。原代星型胶质细胞和小胶质细胞系中,给予Muscimol后可降低脂多糖(lipopolysaccharide,LPS)诱导的NF-κB (nuclear factor-κ-gene binding,NF-κB)中p65亚基的升高[8],后者是炎症反应的关键组成部分。此外,体外研究还证明,Muscimol可以通过抑制小胶质细胞炎症因子的释放,从而进一步保护了神经元的活性[12],但是其中具体的机制仍不清楚。
总之,研究表明,GABAAR激动剂似乎可以通过激活小胶质细胞上的GABAAR从而发挥抗炎作用。本文即验证GABAAR激动剂对炎症因子的抑制作用,并探究其背后的分子机制。
1 材料与方法
1.1 材料
DMEM培养基和胎牛血清(FBS,Gibco),脂多糖(LPS,Sigma-Aldrich),γ-氨基丁酸(GABA,Sigma),蝇蕈醇(Muscimol,Sigma-Aldrich),印防己毒素(PTX,Tocris),P65抗体(CST),Mouse TNF-α ELISA Kit(北京百智生物科技公司),Mouse IL-6 ELISA Kit(北京百智生物科技公司),双荧光素酶报告检测试剂盒(翌圣生物科技公司),乳酸脱氢酶细胞毒性检测试剂盒(碧云天),MTT(Sigma)。
1.2 细胞培养及转染
小鼠小胶质细胞(BV2细胞)或神经母细胞瘤细胞(SH-SY5Y细胞)培养于含有10%胎牛血清的DMEM培养基中,待细胞在培养箱中密度达到80% ~ 90%时进行传代,达到70% ~ 80%时用PEI进行转染。
1.3 ELISA法检测炎症因子的释放
BV2细胞培养于96孔板中,使用浓度为1~100 μmol/L GABA/Muscimol预处理30 min,加入终浓度为1 μg/mL LPS处理12 h;对于GABAAR的氯通道抑制剂Picrotoxin(PTX)在GABA处理前10 min加入到培养基中。收集细胞培养基,300 g离心10 min。收集离心后的培养基上清液,使用酶联免疫吸附测定(ELISA)试剂盒依照说明书的步骤,检测培养基中肿瘤坏死因子-α(TNF-α)和白介素-6(IL-6)水平。
1.4 细胞免疫荧光
将细胞播种于提前用多聚赖氨酸包被35 mm直径的培养皿,随后将其培养在37 ℃、5% CO2饱和湿度的条件下,12 h后对其加药处理。到达既定时间后,弃除培养基,用0.01 mol/L PBS清洗3次。随后用4%多聚甲醛和4%蔗糖的细胞固定液固定15 min,再用3%牛血清白蛋白和0.1% Triton X-100细胞封闭液封闭15 min。用0.01 mol/L PBS清洗细胞5遍,用相应的抗体进行孵育染色,随后使用Leica TCS SP5进行成像。
1.5 双荧光素酶检测
BV2细胞培养于24孔板中,将100 ng报告基因质粒pNF-κB-Luc和10 ng pRL-TK质粒用PEI共转染入BV2细胞。转染48 h后收取细胞,使用双荧光素酶检测试剂盒依照说明书的步骤,测定细胞的荧光素酶活性。
1.6 MTT法检测细胞活力
将细胞接种于96孔板中。待细胞长至孔板的40% ~ 60%时,进行细胞转染操作。转染后4 ~ 6 h进行细胞换液,18 h后弃去孔板中培养基,并向每孔加入10 μL 5 mg/mL的MTT溶液,37 ℃继续培养4 h。4 h后弃去液体,向每孔加入100 μL DMSO,室温避光孵育10 min。用酶标仪仪器,选择490 nm波长进行检测。
1.7 乳酸脱氢酶(lactate dehydrogenase,LDH)释放法检测细胞毒性
将细胞接种于96孔板中,待细胞密度于40% ~ 60%时进行细胞加药处理。其中,加药处理到达既定时间后,取细胞上清液置于新96孔板中,用LDH细胞毒性检测试剂盒(碧云天)测定细胞的毒性。
1.8 统计学方法
2 结果
2.1 在BV2细胞中,脂多糖(LPS)刺激促进炎症因子的释放并会对SH-SY5Y细胞造成损伤
之前有文献[13]报道,LPS刺激会造成小胶质细胞的炎症因子的释放。因此,本文使用了小鼠小胶质细胞系BV2细胞,将BV2细胞接种到96孔板中,培养16~24 h,加入不同浓度的LPS处理12 h,并收集培养基上清。用ELISA法检测其炎症因子的释放情况。结果显示:与无处理对照组相比,TNF-α和IL-6的释放水平与LPS的处理浓度有剂量依赖关系,与无处理组相比,浓度为10-3~ 100μg/mL的LPS均促进了TNF-α和IL-6的释放(P<0.01,Fig.1A-B)。
LPS刺激后小胶质细胞会激活,激活的小胶质细胞会释放炎症因子,不断增多的炎症因子会刺激周围的神经元,进而导致神经元的死亡。为了进一步观察使用经LPS刺激后的BV2培养基与SH-SY5Y神经母细胞瘤细胞共培养,进一步查看SH-SY5Y细胞的存活情况。LDH检测的结果表明:SY5Y细胞的损伤情况与LPS使用的浓度有剂量依赖关系,浓度为10-2~ 100μg/mL的LPS刺激SYSY细胞后均减少了细胞的活性(P<0.001)。MTT检测结果表明:浓度为10-2~ 100μg/mL的LPS刺激SYSY细胞后,均使细胞的数量减少(P<0.001,Fig.1C-D)。
为了排除这种损伤增加是BV2细胞死亡导致的,本文用LDH和MTT法检测了同样剂量、时间范围的LPS对BV2细胞数量和活力的影响。结果显示,本文中所使用的LPS的剂量、时间范围,对BV2细胞未造成毒性(P>0.05,Fig.1E-H)。
为了进一步排除SH-SY5Y细胞的损伤不是因为LPS对其造成的直接伤害,使用LDH和MTT法进一步检测。结果显示:同上述相同剂量LPS刺激后,SH-SY5Y细胞的数量和活力无明显损伤(P>0.05,Fig.1I-J)。
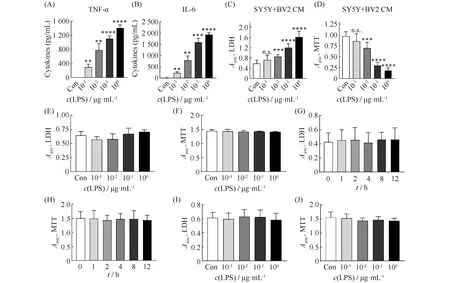
Fig.1 LPS treatment could promote the release of TNF-α and IL-6 in BV2 cells and cause damage to SH-SY5Y cells (A-B) BV2 cells were treated with 10-3-100 μg/mL LPS for 12 hours. The release of TNF-α and IL-6 was detected by ELSIA. Values are presented as mean ± SD. One-way ANOVA followed by Tukey’s post hoc test. **, P<0.01; ***, P<0.001; ****, P<0.0001 compared with control (n= 3 independent experiments for each group). (C-D)SH-SY5Y cells were treated with conditioned medium from BV2 cells (BV2 CM) stimulated by LPS 10-3-100 μg/mL, and cytotoxicity and cell viability was detected with the LDH and MTT assays. Values are presented as mean ± SD. One-way ANOVA followed by Tukey’s post hoc test. ***, P<0.001; ****, P<0.0001 n.s., not significant, compared with control (n= 6 independent experiments for each group). (E-H) BV2 cells were treated with 10-3-100 μg/mL LPS for 12 hours. Cytotoxicity and cell viability was detected with the LDH and MTT assays. n.s., not significant. Values are presented as mean ± SD. One-way ANOVA followed by Tukey’s post hoc test. n.s., not significant, compared with control (n= 12 independent experiments for each group). (I-J) SH-SY5Y cells were treated with 10-3-100 μg/mL LPS for 12 hours. Cytotoxicity and cell viability were evaluated with the LDH and MTT assays. Values are presented as mean ± SD. One-way ANOVA followed by Tukey’s post hoc test. n.s., not significant, compared with control (n= 9 independent experiments for each group)
上述结果表明,经LPS刺激BV2细胞后所产生的炎症因子可以损伤SH-SY5Y细胞。
2.2 γ-氨基丁酸降低LPS导致的BV2细胞炎症因子的释放,从而保护SH-SY5Y细胞活性
先前有文献报道,GABA可以抑制LPS诱导的小胶质细胞的激活,从而进一步抑制炎症因子的释放。为了确证GABA是否可以降低BV2培养基中炎症因子的水平,用不同浓度的GABA(1、10、50、100 μmol/L)预处理细胞30 min,进一步使用LPS诱导炎症因子的释放。ELISA结果显示:与溶剂对照组相比,GABA可以剂量依赖方式降低炎症因子的表达。其中,100 μmol/L的GABA显著降低TNF-α和IL-6的释放(P<0.001,Fig. 2A-B)。
GABA预处理是否能保护神经细胞免受小胶质细胞激活诱导的死亡呢,为此本文进一步用GABA预处理后的BV2培养基继续培养SH-SY5Y细胞12 h。用MTT和LDH法对SH-SY5Y细胞进行细胞数量和活性的检测。结果显示,与对照组相比,仅加入1 μg/mL的LPS处理的BV2培养基使SH-SY5Y细胞损伤显著(P<0.001);与LPS组相比,GABA预处理能降低BV2对SH-SY5Y细胞的损伤,且随着GABA浓度的增强,SH-SY5Y细胞数量逐渐增加,细胞活力逐渐增强(P<0.05,Fig. 2C-D)。

Fig.2 The pretreatment of GABA could reduce the release of TNF-α and IL-6 caused by LPS in BV2 cells, thus protecting the activity of SH-SY5Y cells (A-B). BV2 cells were pretreated with 0, 1, 5, 10, 50,100 μmol/L GABA for 30 minutes and then 1 μg/mL LPS was added. The release of TNF-α and IL-6 was detected by ELSIA. Values are presented as mean ± SD. One-way ANOVA followed by Tukey’s post hoc test. *, P<0.05; **, P<0.01; ***, P<0.001, n.s, not significant, compared with the group treated with LPS (n= 3 independent experiments for each group) (C-D). SH-SY5Y cells were treated with conditioned medium from BV2 cells (BV2 CM) co-treated with 0, 1, 5, 10, 50,100 μmol/L GABA and LPS, cytotoxicity and cell viability was evaluated with the LDH and MTT assays. Values are presented as mean ± SD. One-way ANOVA followed by Tukey’s post hoc test. ####,P<0.0001;compared with the control group, ***, P<0.001; ****, P<0.0001; n.s, not significant, compared with the group treated with LPS (n= 9 independent experiments for each group)
这些数据表明,GABA能够降低LPS诱导的TNF-α和IL-6的释放,同时也可以进一步减缓对SH-SY5Y细胞的损伤。
2.3 γ-氨基丁酸通过GABAAR,发挥抑制炎症的作用
之前有文献表明,在外周系统中,GABA可以通过激活GABAAR发挥抑制炎症的作用。那么,在中枢神经系统中,GABA是否能通过激活GABAAR从而发挥抗炎作用呢?在BV2细胞中,进一步使用GABA与GABAAR的拮抗剂PTX共处理。结果显示,PTX可以拮抗GABA介导的抑制炎症的作用(P<0.05,Fig.3A-B)。为了进一步确定GABA是通过GABAAR而发挥作用的,使用了GABAAR的特异性激动剂Muscimol预处理BV2细胞。结果显示,Muscimol能以剂量依赖的方式降低炎症因子的释放。其中,与LPS处理组相比,浓度为10 μg/mL的muscimol显著减少了TNF-α和IL-6的释放(P<0.001,P<0.01,Fig. 3C-D)。以上结果说明,GABA通过激活GABAAR发挥抑制炎症的作用。
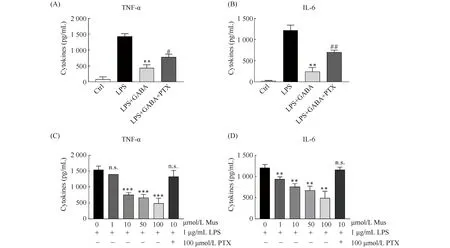
Fig.3 GABA inhibits inflammation through GABAAR (A-B) BV2 cells were pretreated with 100 μmol/L GABA or GABA+PTX for 30 minutes and then treated with 1 μg/mL LPS for 12 hours. The release of TNF-α and IL-6 was detected by ELISA. Values are presented as mean ± SD. One-way ANOVA followed by Tukey’s post hoc test. **, P<0.01, compared with the group treated with LPS, #, P<0.05, ##, P<0.01, compared with the group treated with LPS+GABA (n= 3 independent experiments for each group). (C-D) BV2 cells were pretreated with 0, 1, 10, 50, 100 μmol/L Muscimol or 10 μmol/L Muscimol and 100 μmol/L PTX for 30 minutes and then added 1 μg/mL LPS for 12 hours. The release of TNF-α and IL-6 was detected by ELSIA. ANOVA followed by Tukey’s post hoc test. **, P<0.01; ***, P<0.001, n.s., not significant, compared with control (n= 3 independent experiments for each group)
2.4 GABAAR的激动剂可以通过抑制NF-κB入核,从而抑制炎症因子的释放
为了解析GABA及GABAAR的激动剂抑制炎症因子释放的分子机制。有文献[14]表明,NF-κB只有进入细胞核后才能行驶转录功能而生成炎症因子。那么,GABAAR的激动剂是否通过抑制NF-κB的入核,从而抑制炎症因子的释放呢?为了验证这一假设,本文通过免疫荧光检验了Muscimol预处理后p65亚基的核定位。结果显示,LPS刺激1 h后,p65核内分布升高,而Muscimol预处理可显著抑制LPS导致的P65核移位(P<0.0001)。继而使用了GABAAR的抑制剂PTX预处理细胞。结果显示,PTX反转了P65核移位现象(P= 0.1122,Fig. 4A)。上述结果确证了Muscimol可以抑制LPS介导的p65核移位。
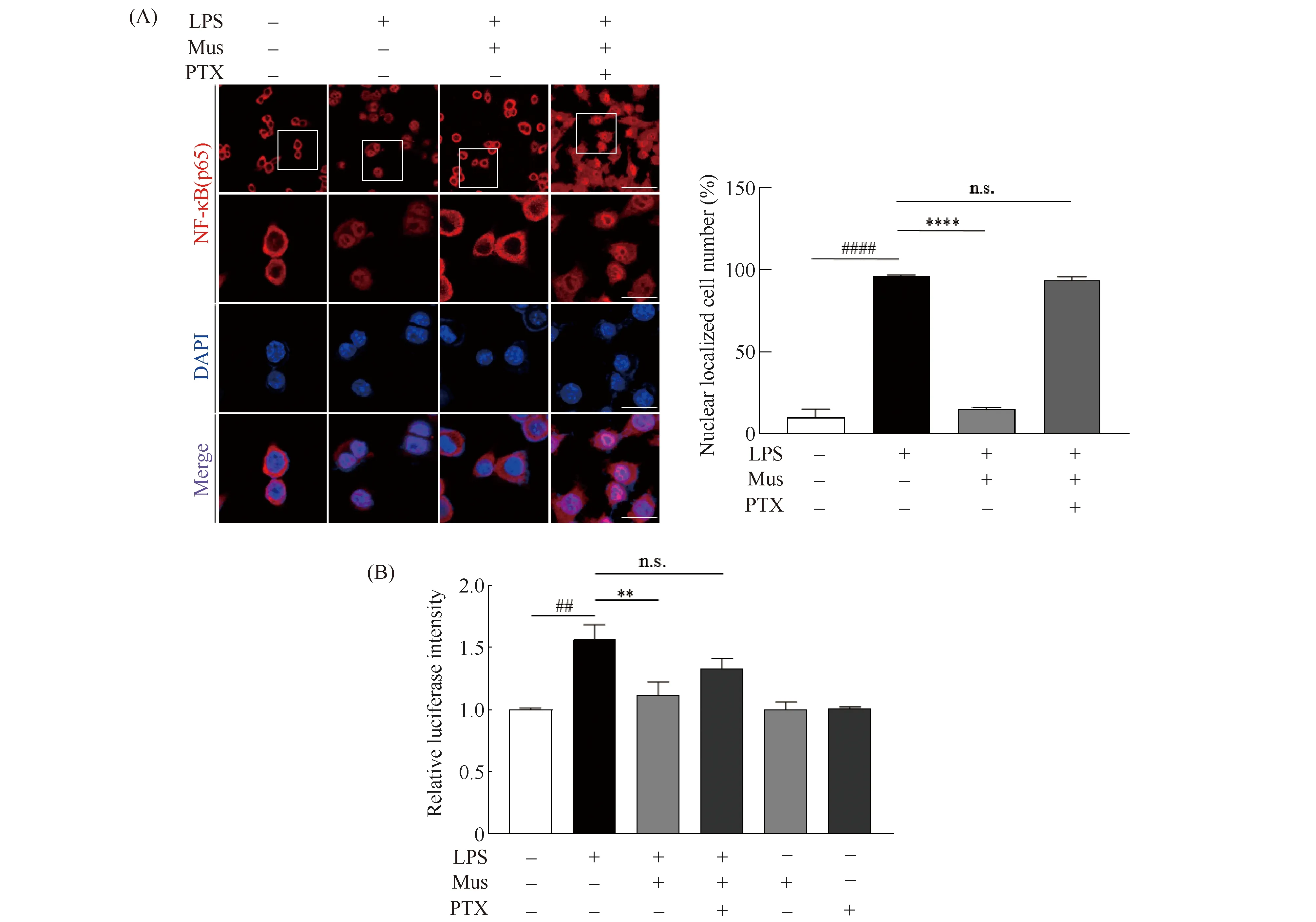
Fig.4 GABAAR agonists could inhibit the nuclear translocation of NF-κB (A) The subcellular localization of p65 was evaluated using an anti-p65 antibody. Bar= 75 mm; Bar (magnification)= 25 mm. Comparisons between the control and other group were made based on the statistical analysis of the cells with nuclear localization of p65 counted in three random fields, ####, P<0.0001, compared with control, ****, P<0.0001, n.s., not significant, compared with the group treated with LPS (n= 6). (B) The relative luciferase intensity was determined 6 hours after LPS stimulation, with or without pre-treatment of muscimol. ##, P<0.01, compared with control, **, P<0.01, n.s., not significant, compared with the group treated with LPS (n= 3). ANOVA followed by Tukey’s post hoc test
基于以上数据,我们提出问题Muscimol的预处理是否就抑制了p65的转录活性呢?为了回答这一问题,本文使用了pNF-κB-luc报告基因质粒,pNF-κB-Luc是以pGL6质粒为模板,可以用于检测NF-κB的转录活性。同时使用CMV-Renilla质粒作为对照组,将pNF-κB-luc和CMV-Renilla以1∶10的比例共转染入BV2细胞,通过荧光素酶检测验证NF-κB的转录活性。结果显示,Muscimol预处理可以显著降低LPS刺激诱导的荧光素信号(P<0.01);进一步使用PTX预处理细胞,结果显示,PTX抑制了荧光素信号的增高(P= 0.0552,Fig. 4B)。以上结果共同说明,GABAAR的激动剂可以抑制NF-κB的入核,进而降低其转录活性,从而抑制炎症因子的生成,最终发挥抑制炎症的作用。
3 讨论
帕金森病(PD)作为第2常见的神经退行性疾病,严重影响人们的身心健康。临床上,PD患者有严重的运动障碍和中脑SN区大量多巴胺能神经元丢失的特性[15]。目前,PD病因尚不清楚。越来越多的证据表明,神经炎症在帕金森病的发生发展中发挥重要作用。当神经炎症发生时,以小胶质细胞为主的免疫细胞过度激活,释放促炎因子,导致周围神经元退化[16-18]。LPS是革兰氏阴性菌细胞壁的组成部分,可引起炎症反应[19]。有研究表明,在大鼠SN中注射LPS可诱发PD症状[20, 21]。小胶质细胞是中枢神经系统中的免疫细胞,是神经炎症反应的主要参与者。有研究报道,PD患者和PD模型动物的SN中存在大量异常激活的小胶质细胞[22, 23]。因此,抑制小胶质细胞的过度激活被认为是防治PD的潜在策略。
GABA能信号与免疫调节过程密切相关。因此,GABA可能是调节神经炎症,进而缓解PD进程的潜在治疗靶点。之前的研究表明,在人类的小胶质细胞中,GABAAR激动剂可以显著减少小胶质细胞TNF-α和IL-6的释放水平[8]。Liu等[12]研究结果表明,在小鼠的PD模型中,GABAAR的激动剂改善了小胶质细胞的过度活化。在我们的结果中也同样观察到,在BV2细胞系中,GABA通过激活GABAAR发挥抑制炎症的作用,也可以进一步保护神经细胞免受损伤。
机制方面:NF-κB是一种经典的炎症途径,参与细胞炎症和许多神经系统疾病。有研究报道,NF-κB信号通路的激活是由细胞外刺激引起的。这些刺激被细胞膜上的受体识别,并传递到细胞中,通过信号级联,激活IκB。当IκB被激活会进一步发生磷酸化进而降解,此时没有IκB的抑制作用,NF-κB复合物就会暴露其入核信号从而转录相关的炎症因子[14]。实验中,通过细胞免疫荧光和双荧光素酶检测也验证了,Muscimol通过激活GABAAR,明显抑制了LPS介导的P65核移位及进一步的转录,从而降低了炎症因子的产生。
尽管本研究中提出GABA-GABAAR-NF-κB信号通路发挥抗炎效应的可能机制,但是GABAAR激活后是如何进一步激活NF-κB信号通路的呢,以及GABAAR是离子型受体,具体是如何发挥代谢作用呢,以上这些问题仍需进一步研究。总之,本文确证了GABA可以抑制炎症因子的生成,从而起到神经保护作用,并初步探索了该作用背后的分子机制。该研究可以为研发GABA-GABAAR-NF-κB通路的靶向新药提供重要的理论依据。针对GABAAR的治疗也有望成为PD防治的新策略。
