夜间不同LED 补光时段对番茄幼苗生长生理指标的影响
2022-08-27刘志强朱新红刘勇鹏
刘志强,朱新红,刘勇鹏,王 清,李 春,张 婵,姜 俊
(1.驻马店市农业科学院 河南驻马店 463000; 2.漯河市农业科学院 河南漯河 462000;3.河南农业大学园艺学院 郑州 450002)
番茄是全球栽培最普遍的一种蔬菜,也是我国北方设施栽培的重要蔬菜之一,番茄果实风味独特且营养、药用价值高。研究发现,光在植物生长和发育过程中具有重要作用,如提供能量、传递信号、影响形态生长及光合作用等。番茄是一种喜光、喜温、短日照的作物,设施内光环境变化对番茄生产有很大的影响。当前冬春季低温弱光、寡照、连阴(霾)等恶劣天气条件已成为限制设施番茄生产的重要制约因子。有研究发现,采用人工补光技术可以改善和调节设施内环境,提高作物产量和品质。李晓慧等研究发现,补光处理45 d 后,番茄幼苗中的可溶性糖和蛋白质含量显著高于对照;闫晓花等研究发现,补光可以显著提高根系活力,提高黄瓜幼苗质量;杨振荣等研究发现,经过补光处理,可以降低香蕉假植苗中丙二醛含量,提高香蕉假植苗中POD、SOD 活性。此外,也有研究发现不同光周期对植物生长的影响不同。佘新等研究发现,利用红光进行夜间暗期打断处理,可抑制番茄及黄瓜幼苗茎徒长,提高壮苗指数;还有研究表明,不同光周期对不同植物叶绿素含量和光合作用的影响差别较大,暗期多次打断处理可使番茄幼苗叶绿素含量显著增加,光合速率显著提高。
众所周知,育苗是番茄生产中重要的技术环节,直接关系到番茄后期的生长发育。LED 具有节能高效的显著特征,在番茄育苗期采用LED 补光能够改善温室内光照条件,明显促进了植株器官的生长,提高壮苗指数,极大减轻了弱光造成的危害,促进了幼苗的生长发育,进而达到培育壮苗的目的。前人对夜间补光的研究大多集中在夜间延时补光时间长短、光质配比、光照强度上。夜间延时补光只是改变了光周期,少有涉及打破暗期及时长的研究。笔者利用LED 光照,进行不同时间段夜间补光,比较其形态指标、光合指标、品质特性和抗氧化酶活性,明确了夜间补光对植物幼苗生长发育的影响,以期为温室番茄育苗中选择合理夜间补光时段及补光技术的推广应用提供参考依据。
1 材料与方法
1.1 材料及设备
试验材料:番茄品种Money Maker。
广东三雄极光股份有限公司生产L100 cm×W80 cm 的LED 白色灯板,白色光板由全光谱LED等组合获取(光谱图见图1),使用智能开关定时器。

图1 光谱图(白光)
1.2 试验设计
试验于2020 年8—10 月在河南省郑州市河南农业大学2 号楼人工气候室中进行。番茄种子晾晒、选种、消毒后进行浸种催芽,待种子萌发后播种于∶=1∶1 的基质中,每穴1 粒,待长到3 叶1 心时,选取生长一致的幼苗放置于5 种不同时间的夜间补光条件下处理,按照随机区组设计,每个处理60 株番茄幼苗,3 次重复。光质为白光。为避免资源浪费,在前人研究的基础上,光照强度统一为90 μmol·m·s。采用定时器设置5 种不同的补光时间,见图2,分别为:夜间不补光(CK);白天正常照射结束后隔1.5 h 补光3 h(T1);白天正常照射结束后每隔3 h 补光1 h(T2);白天正常照射结束后隔4.5 h 后补光3 h(T3);白天正常照射前进行补光3 h(T4)。气候室白天温度设定为25~27 ℃,夜间温度17~20 ℃,湿度60%~80%。每隔3~4 d 浇灌1 次Hoagland 营养液,为保证光照的均匀性,每隔1 d 变动植株位置。

图2 补光时间
1.3 测量项目及方法
1.3.1 生长形态指标测定 光处理7、14、21、28 d
后分别测定株高、茎粗、叶片数。用直尺测株高(茎基部至生长点的长度);用游标卡尺测量茎粗(茎基部1 cm 处);用观察法统计叶片数。
1.3.2 生理生化指标测定 28 d 时测植株叶片中的叶绿素、光合指标、可溶性糖、可溶性蛋白、脯氨酸、MDA 含量和抗氧化酶(SOD、POD 和CAT)活性,同时测定植株根系活力和壮苗指数。壮苗指数=(茎粗/株高)×全株总干质量,参考季方等方法计算。叶绿素含量采用∶=1∶1 溶液提取;叶片的净光合速率、气孔导度、胞间CO浓度和蒸腾速率均用LI-6400XT 便携式光合仪来测定;采用TTC 法测定其根系活力;采用蒽酮比色法测定可溶性糖含量;用考马斯亮蓝法测定蛋白质含量;参照植物生理生化实验原理与技术测定脯氨酸含量;参照Huo 等的方法测定SOD、POD 和CAT 活性,采用分光光度法测定MDA 含量。
试验随机取样,每次取样15 株,3 次重复。
1.4 数据处理
采用Microsoft Excel 2016 和SPSS 24 软件对数据进行分析,用最小显著差数法(LSD)进行显著性差异分析。
2 结果与分析
2.1 夜间不同补光时段对番茄幼苗生长形态指标的影响
由图3 可知,从14 d 开始,不同处理的番茄幼苗株高均高于对照,且定植时间越长,补光处理的幼苗株高与对照的差别越显著。定植14 d 和21 d后,T1、T2、T3 处理之间幼苗株高差异不显著,但均显著高于T4 处理和CK;28 d 时,以T2 处理株高最高,其中T2 和T3 处理间无显著性差异,但二者均显著高于T1、T4 处理和CK。番茄幼苗定植28 d时,不同补光方式下幼苗的株高为T2>T3>T1>T4,但T2 与T3 仅在定植7 d 时差异显著,在定植后期差异不显著。由此可知,T2、T3 处理对增加番茄株高效果相对较好,其中以T2 效果最佳。
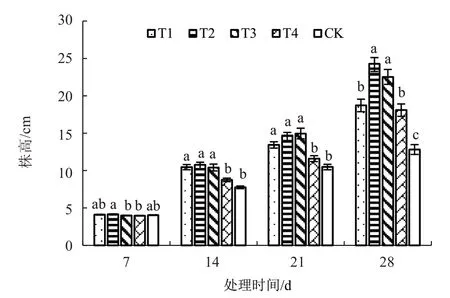
图3 夜间不同补光时段对番茄幼苗株高的影响
由图4 可知,在番茄幼苗定植14 d 后,夜间不同时段补光开始对番茄幼苗茎粗有一定影响;14 d和21 d 时,夜间不同时段补光处理番茄茎粗均显著高于对照,但各处理间无显著差异;定植28 d 时,以T3 最粗,T2 和T3 处理茎粗均显著高于其他处理,但二者之间无显著性差异。定植28 d 时,各处理幼苗茎粗为T3>T2>T1>T4。由此可知,T3、T2 处理对增加番茄茎粗效果较好,以T3 处理效果最佳。

图4 夜间不同补光时段对番茄幼苗茎粗的影响
由图5 可知,在定植后7 d 时,不同补光处理间及各处理与对照间叶片数没有显著性差异。处理后期,夜间任何补光处理均使幼苗叶片数显著高于对照,且T2 与T3 处理间差异不显著。14 d 时各补光处理叶片数为T2>T1>T3>T4,前三者间差异不显著,但T2 显著高于T4;21 d 时各补光处理叶片数为T3>T2>T4>T1,各处理间差异不显著;28 d时叶片数为T3>T2>T4>T1,T3 显著高于T1。由此可知,在幼苗定植初期14 d 时,T2 处理促进番茄幼苗叶片数的增加效果较好,后随着定植时间延长,T3 效果优于T2。
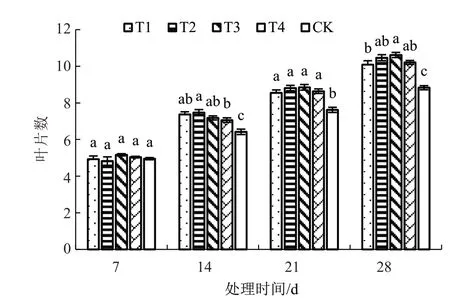
图5 夜间不同补光时段对番茄幼苗叶片数的影响
综上可知,夜间不同补光时段处理在番茄幼苗形态指标上,以T2 和T3 补光处理对增加番茄幼苗株高、茎粗、叶片数效果相对较好。其中T2 对增加株高效果最好,而T3 对增加茎粗效果最佳,在叶片数上,定植后期T3 处理优于T2 处理。整体说明T3 处理(白天正常照射结束后延时4.5 h)补光效果最佳,且补光连续性与否对增加番茄幼苗的株高、茎粗、叶片数差异不显著,而白天正常照射结束后延时1.5 h 补光或者延时9 h 再补光,对增加番茄幼苗株高和茎粗的效果相对较差。
2.2 夜间不同补光时段对番茄幼苗壮苗指数和根系活力的影响
由表1 可知,不同处理番茄壮苗指数和根系活力均显著高于对照,但不同补光处理间存在一定差异。各处理壮苗指数表现为T3>T2>T4=T1>CK,T3 壮苗指数最大,且显著高于其他处理,相比对照提高54.55%,其次是T2 处理,比对照提高27.27%,此外,T1、T2 和T4 处理间无显著差异。各处理根系活力大小为T3>T2>T4>T1>CK。除T1、T4 间无显著差异外,其他处理间均存在显著性差异。其中T3 处理相比对照提高45.22%,T2 处理相比对照提高23.69%。综上可知,进行夜间补光的番茄幼苗,在壮苗指数和根系活力指标上均比不补光高,整体以T3 处理苗子长势最强,其次是T2 处理。
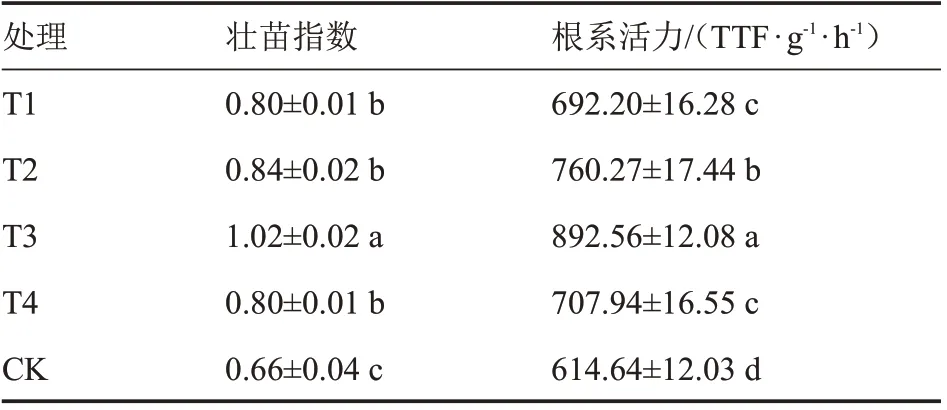
表1 夜间不同补光时段对番茄幼苗壮苗指数和根系活力的影响
2.3 夜间不同补光时段对番茄幼苗叶绿素含量的影响
由表2 可知,不同补光时段对番茄叶片的光合色素含量有一定影响。T3 处理叶绿素a 含量显著高于其他处理;T2 和T4 处理叶绿素a 含量则无显著差异,但均显著高于T1 处理和CK;T1处理和CK 则无显著性差异。T3 处理叶绿素b含量显著高于其他处理,其他处理之间叶绿素b 含量无显著性差异。T3 处理类胡萝卜素含量最高,显著高于其他处理,其次是T2 处理,T1、T4和CK 之间无显著性差异。各处理之间叶绿素(a/b)无显著性差异,但T3 处理含量仍相对较高。叶绿素(a+b)含量变化趋势和叶绿素a 相同。综上可知,不同补光时期处理均能有效增强番茄叶片中光合色素的含量,其中以T3 处理效果最佳。

表2 夜间不同补光时段对番茄幼苗叶绿素含量的影响
2.4 夜间不同补光时段对番茄幼苗光合特性的影响
由表3 可以看出,不同补光处理对番茄幼苗的光合特性产生了一定影响。蒸腾速率的变化趋势与净光合速率一致,T3 处理净光合速率和蒸腾速率均最高,显著高于其他处理和对照,分别比CK提高43.76%和122.57%。气孔导度和胞间CO浓度变化趋势基本一致,T3 处理气孔导度和胞间CO浓度均与对照差异不显著,分别增加13.33%和1.77%,但二者均显著高于其他3 组处理。综合4 个指标,T3 处理下,番茄幼苗的光合特征值均最高。除此之外,T4 和CK 处理的蒸腾速率和净光合速率差异不显著,但均显著低于其他处理;T1、T2 间气孔导度差异不显著,且显著大于T4;T1、T2、T4 胞间CO浓度差异不显著。综上可知,夜间不同补光时段处理在番茄光合特性指标上,以T2 和T3 补光处理对增加番茄净光合速率及蒸腾速率相对较好,其中T3 处理净光合速率、气孔导度、蒸腾速率均高于其他处理及对照,表明白天正常照射结束隔4.5 h 后,连续3 h 的补光处理最有利于提高番茄幼苗的光合效率。

表3 夜间不同补光时段对番茄幼苗光合特性的影响
2.5 夜间不同补光时段对番茄幼苗叶片中可溶性糖、可溶性蛋白和脯氨酸含量的影响
由表4 可知,不同夜间补光时段番茄幼苗可溶性糖含量为T3>T2>T4>T1>CK。T3 处理可溶性糖含量最高,与T2 差异不显著,但比T1、T4 和CK 显著提高21.91%、14.51%、35.41%,T1 可溶性糖含量与T4、CK 差异不显著。不同补光处理下番茄幼苗中可溶性蛋白含量为T3>T2>T1>T4>CK,所有处理均显著高于对照,其中T3 处理可溶性蛋白含量(,后同)最高,为18.66 mg·g,对照可溶性蛋白含量最低,为14.66 mg·g。与对照相比,T3 处理可溶性蛋白含量显著提高27.28%,T1、T2、T4 处理之间无显著差异。这表明T3 处理最有利于增加番茄幼苗中的可溶性蛋白含量。不同补光处理下番茄幼苗中脯氨酸含量为T3>T2>T4>T1>CK,脯氨酸含量变化趋势与可溶性蛋白含量基本一致,T3 处理下番茄幼苗中脯氨酸含量最高。
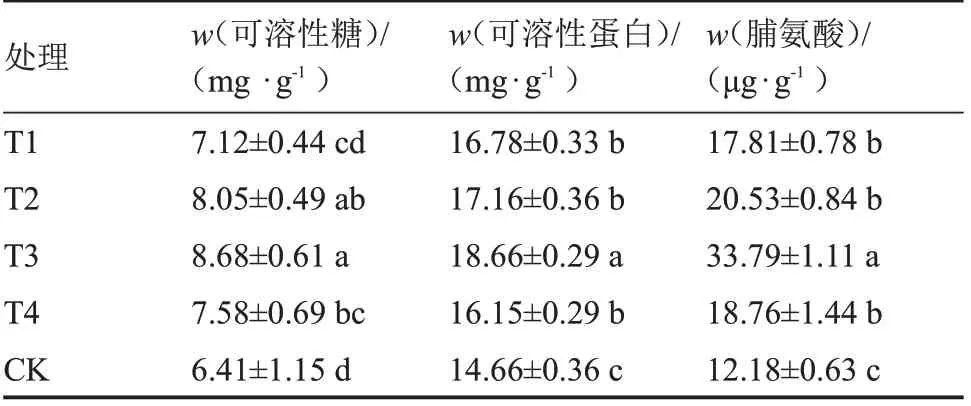
表4 夜间不同补光时段对番茄幼苗叶片中可溶性糖、可溶性蛋白和脯氨酸含量的影响
2.6 夜间不同补光时段对番茄幼苗叶片抗氧化酶活性及MDA含量的影响
由表5 可知,不同补光处理下番茄幼苗叶片POD 活性为T3>T2>T1>T4>CK,T3 的POD 活性显著高于其他处理和对照,T2 显著高于对照,T1、T2、T4 间差异不显著,T1、T4 与对照相比差异不显著。不同补光处理下CAT 活性为T2>T1>T3>T4>CK,T2 的CAT 活性最高,各处理CAT 酶活性均显著高于CK,但不同处理间差异不显著。不同补光处理下SOD 活性为T3>T4>T2>CK>T1,T3 处理的SOD 活性最高,与T4 差异不显著,但显著高于T1、T2 和CK;T1、T2、T4 与CK 差异不显著。不同处理下MDA 活性为T4>CK>T1>T2>T3,T4 处理的MDA 含量最高,显著高于其他处理;T1、T2 与CK 之间差异不显著;T3 处理的MDA 含量最低,显著低于CK。

表5 夜间不同补光时段对番茄幼苗叶片中抗氧化酶活性及MDA 含量的影响
3 讨论与结论
番茄生产中关键环节是育苗,设施内因覆盖材料、季节性导致的低温寡照等因素,光照时长远低于露地,严重制约着温室内冬春番茄的生长发育。随着现代农业的快速高效发展,为满足市场的需求,集约化、标准化、规模化、智能化的蔬菜育苗工厂已逐步走向市场,而LED 光源作为新型节能光源,具有光谱广泛、耗能小、发热少和寿命长等优点,已经迅速应用到蔬菜育苗产业中,为设施蔬菜优质种苗培育产生了积极的影响。试验研究了夜间温室条件下不同时段LED 补光对番茄幼苗生长发育的影响,结果表明,不同补光方式对设施中番茄幼苗的形态指标、壮苗指数、根系活力、光合特性及可溶蛋白、可溶性糖含量等不同指标影响不尽相同。
在番茄幼苗形态指标上,笔者研究发现T2 和T3 补光处理对番茄幼苗株高、茎粗、叶片数的效果相对较好,显著高于对照,且补光连续性与否对影响番茄幼苗形态指标差异不显著。这与陈丽丽等研究表明经过暗期打断处理,菜心的株高、茎粗、叶片数等高于对照,以及佘新认为暗期每隔2 h 补充5 min 红光光照,黄瓜幼苗的茎粗显著高于对照的结果相一致。
在番茄幼苗的壮苗指数和根系活力上,有研究发现植物根系强壮程度直接影响其对水分和营养物质的吸收能力。本试验结果表明,夜间不同时间补光均可提高番茄幼苗的壮苗指数和根系活力,且T3(白天正常光照4.5 h 后进行3.0 h 暗期光打破)处理显著高于其他时段补光处理。这与李晓慧等在19:00—23:00 补光处理显著提高了番茄幼苗根系活力等研究结果一致。
有研究表明光合速率代表植物吸收二氧化碳的能力,二者呈正相关;蒸腾速率反映根系对水分的吸收能力,直接影响光合作用;气孔导度表示气孔张开的程度,直接影响植物的光合作用和蒸腾作用。本研究发现,不同补光处理对番茄幼苗光合特性的影响不尽相同,其中T3 处理的幼苗净光合速率、蒸腾速率、叶绿素含量均显著高于其他处理和对照,这与段齐珍等认为LED 补光能够提高设施中黄瓜叶绿素a/b 含量、蒸腾速率及番茄幼苗叶绿素含量,以及佘新等认为红光暗期多点打断提高番茄幼叶的叶绿素含量和光合速率的结果相似。本研究结果也显示夜间T3 补光处理下番茄幼苗胞间CO浓度、气孔导度虽显著高于其他处理,比对照也有所增加,但差异不显著,其他时段补光则均显著低于对照,这与陈小玲等认为补光提高了“人参榕”的气孔导度不一致,可能是由设置的光照度、光质存在差异造成的。
不同补光处理对番茄幼苗叶片中可溶性糖、可溶性蛋白、脯氨酸、MDA 含量及保护酶(SOD、POD、CAT)活性影响的研究结果显示,T3 处理下番茄幼苗的可溶性糖、可溶性蛋白质、脯氨酸含量、过氧化物酶活性、超氧化物歧化酶活性均显著高于其他处理及对照,过氧化氢酶活性显著高于对照,丙二醛含量低于其他处理并显著低于对照,表明白天结束光照后延时4.5 h 进行夜间补光能最大程度地缓解膜脂过氧化,使幼苗具有较强抗性,提高其抗逆性。这与刘磊等发现对洋葱幼苗叶片进行暗期红光打断处理可提高洋葱幼苗叶片的可溶性蛋白质含量和POD 活性及前人研究在设施中番茄幼苗、黄瓜幼苗、香蕉假植苗补光处理时,可显著提高补光下的植物可溶性糖、可溶性蛋白质、脯氨酸含量及过氧化物酶、过氧化氢酶、超氧化物歧化酶活性,同时显著降低丙二醛含量的结果相一致,但涉及物质积累在幼苗不同器官中分配率的差异,还有待进一步研究。
光环境作为植物生长最为密切的因素之一,通过对番茄植株延长光照周期,可以明显提高作物光照积累量,不仅可以有效促进番茄开花,利于番茄提早上市,而且还能显著提高番茄第一穗果单果质量和单株产量,对干物质分配、开花和抗病等产生影响。伴随着LED 植物补光灯的快速发展,利用光调控技术来培育壮苗逐渐成为一项经济有效、节能环保且简便的新方法,而研究不同作物在不同生长期筛选出适宜的补光时段也将是未来LED补光技术在设施蔬菜生产应用上的重要研究内容。
综上所述,白天正常光照4.5 h 后进行3.0 h 暗期补光是对设施番茄幼苗生长发育最为有利的夜间补光时间段,最能提高番茄幼苗的根系活力、光合能力和保护酶活性,有利于幼苗可溶性蛋白、可溶性糖及脯氨酸等物质的积累。
