黄连NRAMP3基因的克隆与生物信息学分析*
2022-08-26王文斌刘义飞胡志刚
王文斌 ,刘义飞 ,刘 迪 ,胡志刚 **,汪 波
(1. 湖北中医药大学药学院 武汉 430065;2. 湖北省药品监督检验研究院 武汉 430075)
中药黄连(Coptidis Rhizoma)来源于毛茛科植物黄连Coptis chinensisFranch.、三角叶黄连Coptis deltoideaC.Y.Cheng et Hsiao 或云连Coptis teetaWall.的干燥根茎[1],是我国传统中药材,生长于海拔500-2000 m间的山地林中或山谷阴处,主要分布在我国湖北、重庆、四川、陕西南部、贵州、湖南等地,其中湖北利川和重庆石柱是我国黄连的主产区,约占黄连总产量的80%以上。植物可以从土壤中吸收各种养分和必需元素来维持生长和发育。然而,一些有毒物质(如镉)也会积累,这可能会对植物的生长发育、生理和生化特性产生不利影响。镉作为有毒的重金属之一,可以降低植物体内酶的活性及光合强度,造成代谢紊乱,改变细胞膜透性,阻碍根系生长,抑制其对水分和养分的吸收[2],最终影响作物的产量和品质。此外,还会导致植物细胞内活性氧的产生与清除系统的平衡遭到破坏,甚至导致植物死亡[3-4]。在一项研究中表明,114 批次的黄连样本中,仅有20份样本镉含量在黄连镉限量值的正常范围内,超标率为82.45%,镉超标情况严重[5]。因此,镉污染可能对黄连造成很大的损害,甚至危害消费者的健康。随着现代工业、城市活动和交通的快速发展,黄连中的镉污染已经成为影响其出口的重要因素。因此,研究黄连对镉的吸收和运输机理对其抗镉和减镉具有重要意义。
为了将重金属的毒性降到最低,植物形成了一个完整的机制来控制金属的吸收、运输和积累分布。在重金属胁迫下,一些植物会在根部积累金属离子,限制根到地上部的运输,从而减轻地上部分的毒害。植物细胞通过金属螯合蛋白来螯合金属离子,并通过金属膜转运蛋白将金属离子运输到液泡或从细胞中运送出去,从而表现出对这些重金属的抵抗能力。自然耐药相关巨噬细胞蛋白(NRAMP)是一类内膜转运蛋白,通常有10-12 个跨膜结构域(transmembrane domain,TMD),1-2个糖基化位点和1个转运蛋白特征的结构域[6-7]。NRAMP最先在小鼠身上发现[8],并已被确认为细菌、真菌、昆虫、植物和哺乳动物中的一种二价金属转运蛋白[9-10]。
植物中的NRAMP 蛋白参与多种二价阳离子的运输,如 Fe2+、Mn2+、Cu2+、Pb2+和 Cd2+[11,12]。NRAMP 蛋白在拟南芥[13-15]、水稻[16-18]、东南景天[19-20]、大豆[12]、可可豆[21]和甘蓝型油菜[22]中的作用已被分析。拟南芥中有6 个NRAMP 蛋白家族成员,通过系统发育分析,将其分为两个亚家族,AtNRAMP1 和 AtNRAMP6 属于第 I 组,AtNRAMP2、AtNRAMP3、AtNRAMP4、AtNRAMP5 蛋白属 于 第 II 组[23],并 且 AtNRAMP1、AtNRAMP3、AtNRAMP4 可以转运 Fe,Mn 和 Cd[15,24],而 AtNRAMP6只转运Cd[25]。OsNramp1 可能参与镉在植物根木质部的吸收,从而影响镉向地上部的转运[18],而OsNramp5蛋白在水稻吸收锰和镉过程中起着至关重要的作用[16-17,26-27]。东南景天SaNRAMP1 定位于质膜可以运输 Zn、Mn 和 Cd[19],SaNRAMP6 直接参与镉的吸收,可以提高镉的吸收速率和积累量[20]。可可中TcNRAMP3和TcNRAMP5 可以转运生长必需的二价金属阳离子(Fe、Mn)和 Cd2+,敲除TcNRAMP5 有望获得低 Cd 品种可可[21]。多种植物的NRAMP 蛋白在Cd转运中的作用已有研究。然而,NRAMP 蛋白家族的功能和镉在黄连中的运输机制仍不明确。
本研究在黄连参考基因组[28]数据库中鉴定了黄连CcNRAMP3基因,通过TA 克隆该蛋白的编码基因,并通过生物信息学分析黄连NRAMP3 蛋白的理化性质、亚细胞定位、跨膜结构域、系统进化关系等,对基因功能以及编码蛋白的性质作出初步的判断和预测,为研究黄连体内重金属镉转运机制以及CcNRAMP3基因的功能作用提供参考依据。
1 材料、试剂与仪器
1.1 材料
黄连植株采自湖北省神农架林区,经肖凌博士鉴定为毛茛科植物味连(Coptis chinensis),移栽培养在湖北省药品监督检验研究院分子生物实验室光照培养箱中,设置光照培养箱温度为19±2℃;湿度为60%;光强1000-1500 Lx;光照时间14 h/天。实验材料系霍格兰营养液培养3 周长势良好的黄连叶片,洗净后液氮冷冻,置于-80℃冰箱备用。
1.2 试剂与仪器
HLingene 琼脂糖凝胶回收/PCR 产物纯化试剂盒购于上海惠凌生物技术有限公司。植物总RNA 提取试剂盒购于天根生化科技(北京)有限公司。pEASY®-Blunt 克隆载体和Trans1-T1 大肠杆菌感受态细胞购自北京全式金生物科技有限公司。IPTG 溶液(50 mg·mL-1)、X-Gal 溶液(20 mg·mL-1)、Amp 溶液(100 mg·mL-1)购自索莱宝生物科技有限公司。高效率PCR 酶KOD Dash 购于东洋纺(上海)生物科技有限公司。引物合成及测序由生工生物工程(上海)股份有限公司完成。100×改良型霍格兰营养液和Thermo First cDNA Synthesis Kit 反转录试剂盒购于武汉莱博金服科技有限公司
T100 型PCR 扩增仪(美国Bio-Rad);冷冻高速离心机(Thermo Fisher Scientific);Nano drop-2000 超微量分光光度计(Thermo Fisher Scientific);DYY-10C 电泳仪(北京六一);Gel Doc XR+凝胶成像系统(美国Bio-Rad 公司);LWB-26 双列六孔电热恒温水浴锅(上海龙跃);光照培养箱(上海博讯)。
2 方法
2.1 获取目的基因
以拟南芥、水稻的NRAMP基因序列为参考序列,比对黄连基因组,通过blast 比对寻找黄连NRAMP家族基因(e 值≤e-10),并且将它们在Pfam 蛋白质家族数据库(http://pfam.xfam.org/search)中进行检验以确保其包含NRAMP保守结构域(PF01566),得到黄连NRAMP3基因。
2.2 黄连叶片RNA的提取与cDNA的合成
黄连叶片用清水冲洗洗净,滤纸吸干,并用液氮研磨成粉末,使用天根植物总RNA 提取试剂盒提取总RNA。使用琼脂糖凝胶电泳和超微量分光光度计检测提取的RNA 质量和浓度。然后以提取的总RNA 为模板,采用RNA 反转录试剂盒反转录合成cDNA,50℃水浴20 min,85℃水浴10 min,灭活逆转录酶,储存在-20℃冰箱中备用。
2.3 引物扩增与黄连NRAMP3基因的克隆
依据已测得的黄连基因组数据,查找该基因上下游1500 bp 长度,分别设计扩增引物(表1:上游引物3L-F 与3L-R;下游引物3R-F 与3R-R)将目的序列片段扩增,以确定黄连NRAMP3基因编码序列。根据获得的黄连NRAMP3基因序列,设计特异性扩增引物NRAMP3-F/R。以反转录得到的黄连叶片cDNA 为模板进行PCR 扩增,获得NRAMP3的完整编码序列。PCR 反应体系为:1μL KOD Dash,5μL 10 × Buffer for KOD Dash,5μL 2 mM dNTPs,3μL cDNA,34μL ddH2O,1μL NRAMP3-F Primer(10μmol·L-1),1μL NRAMP3-R Primer(10μmol·L-1),共 50μL。PCR 反应程序为:95℃ 5 min;95℃ 30 s,57℃ 30 s,74℃ 1 min30 s,反应进行32个循环;74℃ 10 min。
PCR 产物经切胶回收后,纯化产物与pEASY-Blunt 克隆载体连接并转化至Trans1-T1 感受态细胞,随机挑选阳性克隆经菌液PCR 扩增后,送生工生物工程(上海)股份有限公司测序验证,引物信息表见表1。

表1 引物信息表
2.4 黄连NRAMP3基因的生物信息学分析
对黄连NRAMP3基因测序完成后,利用TBtools 软件(版本号为v1.0971)[22]查找黄连NRAMP3 基因的开放阅读框并翻译成氨基酸序列,在NCBI 数据库中对氨基酸序列进行Blast 比对;利用GSDS 在线工具预测NRAMP3的基因结构;利用ExPASy 在线软件分析NRAMP3编码氨基酸序列的组成和理化性质;利用ProtScale 在线工具进行蛋白疏水性分析;利用ProtComp 在线软件对NRAMP3的氨基酸序列进行亚细胞定位;利用SignalP 进行蛋白信号肽预测;利用TMHMM 通过隐马尔可夫模型来预测蛋白跨膜区域判断其是否为跨膜蛋白;利用SPOMA分析NRAMP3的蛋白质二级结构并进行预测,以了解局部空间结构;利用SWISS-MODEL 预测CcNRAMP3蛋白的三级结构;使用MEGA 6.05 软件对CcNRAMP3序列进行多重比对分析,采用Neighbor-Joining 法构建蛋白质系统发育树,Bootstrap 统计检验次数为500,其他参数设置为默认参数。
2.5 黄连NRAMP3基因表达分析
取3年生黄连的须根、根茎、叶柄、叶片材料,依据RNA 提取试剂盒的操作步骤,提取总RNA,Thermo First cDNA Synthesis Kit 反转录试剂盒进行基因的定量表达分析。以黄连actin作为内参基因,检测NRAMP3基因在黄连各组织部分的表达情况(引物见表1)。使用PikoReal 实时定量PCR 仪。每个样品设置3 次重复。数据采用2--△△CT法方法进行处理。
3 结果
3.1 黄连NRAMP3基因的克隆
根据黄连基因组中找到的黄连NRAMP3基因组序列上下游1500bp 设计的引物将NRAMP3序列进行扩增,以确定NRAMP基因序列的首尾端正确序列,并以此为依据设计引物进行DNA扩增,确定该基因的序列全长2771 bp。通过GSDS 网站在线分析基因结构,发现黄连NRAMP3具有4 个外显子和3 个内含子(图1)。

图1 黄连NRAMP3的基因结构分析
依据天根植物RNA 提取试剂盒说明书要求,选取新鲜洁净黄连叶片提取总RNA,将凝胶电泳条带清晰、吸光度与浓度均满足实验要求的RNA 作为模板RNA。用反转录试剂盒将RNA 样品合成cDNA 第一条链。以黄连叶片cDNA 为模板,利用设计的引物扩增了约1500 bp 的条带,与预期结果长度一致(图2)。将该片段切胶回收后与pEASY-Blunt 载体连接,转化至大肠杆菌感受态细胞,挑选阳性克隆的进行菌液PCR,最后将序列提交至GenBank,基因登录号为KAF9619594.1。

图2 黄连CcNRAMP3基因电泳图
用CodonCode Aligner 软件拼接测序结果的峰图,得到1617 bp 的cDNA 序列,经TBtools 软件分析发现其为一个完整的开放阅读框,共编码538 个氨基酸。将该氨基酸序列进行blast 比对,结果表明,与GenBank 已报道的其他植物的NRAMP3蛋白序列具有较高的相似性,将该基因命名为CcNRAMP3。利用Pfam 分析氨基酸序列的保守结构域,发现CcNRAMP3蛋白的第98-461位氨基酸残基是典型的NRAMP结构域(PF01566)。
3.2 CcNRAMP3 基因编码蛋白的序列分析
3.2.1 理化特性分析
利用ExPASy数据库的ProtParam工具(https://web.expasy.org/protparam/)分析黄连NRAMP3基因转化的蛋白序列,发现CcNRAMP3蛋白包含538个氨基酸,分子量为58.93 kD,等电点为5.46,带负电荷的残基(Asp+Glu)为44,带正电荷的残基(Arg+Lys)为37,含有20 种氨基酸,异亮氨酸含量最高,占比12.6%,半胱氨酸含量最低,占比1.1%,分子式为C2714H4267N667O753S20。蛋白质不稳定系数为38.29,属于稳定蛋白质。
使用EXPASY的Protscale在线工具预测NRAMP3蛋白的亲水性和疏水性(图3),发现在第8个氨基酸位置,最低峰值是-3.278,在第511 个氨基酸位置,最高峰值是3.178,平均亲水性为0.496,这表明该蛋白是疏水性蛋白。

图3 CcNRAMP3蛋白的亲疏水性预测结果
3.2.2CcNRAMP3蛋白信号肽、亚细胞定位、跨膜结构域预测
SignalP-5.0 Server软件分析结果显示,含有Sec信号肽(Sec/SPI)的可能性为0.0017,不含有信号肽(Other)的可能性为0.9983,预测CcNRAMP3蛋白不含信号肽序列,不属于分泌型蛋白。使用ProtComp 9.0对CcNRAMP3的亚细胞定位进行预测,预测结果表明CcNRAMP3蛋白质定位于液泡膜上。利用TMHMM Server v. 2.0 预测CcNRAMP3蛋白的跨膜结构域(transmembrane domain,TMD),对应的氨基酸残基区域分别为 78-98、113-135、156-178、188-210、217-234、258-280、301-323、356-378、398-415、430-449、461-483、493-515。结果表明,CcNRAMP3蛋白含有12个跨膜区域,具有膜蛋白的重要特征(图4)。

图4 黄连NRAMP3蛋白跨膜结构域分析
3.2.3CcNRAMP3蛋白二级结构预测
经 PRABI 网站的 SOPMA 程序对CcNRAMP3蛋白二级结构进行分析,预测发现CcNRAMP3蛋白主要由α 螺旋(Alpha helix,Hh)、延伸链(Extended strand,Ee)、β 转角(Beta turn,Tt)和无规则卷曲(Random coi,Cc)四部分结构组成(图5),Alpha helix 有304 个氨基酸,占比56.51%、Extended strand 有 64 个氨基酸,占比11.90%、Beta turn 有19 个氨基酸,占比3.53%、Random coi 有151 个氨基酸,占比28.07%。α-螺旋与无规则卷曲是多肽链的主要结构元件。Alpha helix、Extended strand 和Random coil 贯穿于整个氨基酸链,Beta turn只有一点点,散布在Helix附近。

图5 黄连NRAMP3蛋白二级结构图
3.2.4 三维建模
对CcNRAMP3蛋白的三级结构预测分析显示,CcNRAMP3蛋白与葡萄球菌锰转运突变体MntH 的晶体结构(二价金属阳离子转运体MntH)具有37%的序列相似性,以该蛋白(SMTL ID:5m8a.1.A)为模板,通过同源建模构建CcNRAMP3蛋白的三级结构,用于建模的氨基酸残基范围为70-519位,占编码氨基酸总数的83%,蛋白三级结构如图6。
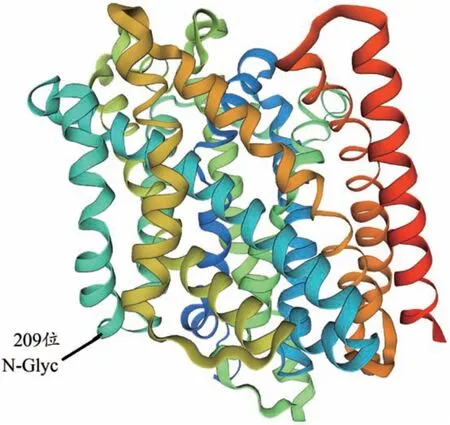
图6 黄连NRAMP3蛋白的三级结构
3.3 多序列比对分析
为研究黄连CcNRAMP3蛋白在物种中的进化位置和亲缘关系,利用MEGA 6.05 对水稻(Oryza sativa)、拟 南 芥(Arabidopsis thaliana)、玉 米(Zea mays)、芝 麻(Sesamum indicum)、烟 草(Nicotiana tabacum)、莲(Nelumbo nucifera)等植物的NRAMP3蛋白序列进行进化树分析比较。进化关系表明(图7),黄 连CcNRAMP3与 博 落 回McNRAMP3(Macleaya cordata, accession no. OVA01006.1)、莲(Nelumbonucifera, accession no. XP_010271562.1)亲缘关系最近 ,与 烟 草(Nicotiana tabacum)、芝 麻(Sesamum indicum)、中 华 猕 猴 桃(Actinidia chinensisvar.chinensis)、枣(Ziziphus jujuba)、梅(Prunus mume)等聚为一个大类。

图7 黄连NRAMP3蛋白的多序列比对分析
3.4 NRAMP3基因表达情况
利用天根植物RNA 提取试剂盒提取水培黄连的须根、根及根茎、叶柄、叶片,采用qPCR 方法分析CcNRAMPs家族在不同组织中的表达水平。结果表明,在未受镉胁迫时,CcNRAMP3基因在根、茎、叶中都有表达,但是不同器官中的表达具有特异性,在黄连地上部分含量比地下部分含量高。在黄连遭受镉胁迫时,在须根表达量显著增加后逐渐下降,叶柄与根茎中的表达量下降一度被抑制,而叶片部位的表达量有较大变化。依据结果初步判断CcNRAMP3基因可能参与Cd胁迫响应,并且与黄连积累Cd的特性相关,并且该基因在根部对Cd胁迫的响应最为敏感。
4 讨论
黄连药用历史悠久,药用始载于《神农本草经》,一名“王连”。称其“味苦,寒,无毒。治热气,目痛,眦伤,肠澼,腹痛,下利,泣出,明目,妇人阴中肿痛。”此后成为历代医家常用的苦寒降火,清热燥湿要药。但随着工业发展,其生长环境遭到严重破坏,尤其是在种植过程中重金属污染、化肥、农药和生长调节剂滥用等,野生黄连的资源越来越少。黄连药用部位为根茎,而其须根和根茎却为镉富集的主要部位,镉富集程度明显大于茎和叶[29],无公害种植黄连的要求显得尤为重要[30]。由于镉不是植物生长所必需的金属元素,在植物中不认为其有专用的转运通路或蛋白[3],故明确黄连中镉及其螯合物相关转运蛋白的性质,对了解黄连体内镉的积累机制,尤其是黄连响应镉胁迫的分子机制方面有着重要的意义,有助于揭示黄连体内镉调节网络,也可以为黄连抗镉育种提供基因资源。
NRAMP家族基因在进化过程中高度保守,广泛分布于哺乳动物、酵母和高等植物中,并参与了Zn、Fe、Mn、Cd 等二价金属。OsNRAMP1定位于质膜,被认为参与了细胞对 Cd 的吸收;OsNRAMP5将 Fe、Mn 和 Cd从外源溶液转运到根细胞[31]。 有趣的是,与OsNRAMP5相近的同源物种大麦(Hordeum vulgare)转运蛋白HvNRAMP5也定位在质膜上,但只运输Cd 和Mn 离子,而不转运Fe[31],两者具有不同的吸收系统。AtNRAMP3定位于液泡膜上,可以将存储在液泡中的金属离子释放到液泡膜附近的细胞质中[33]。小金海棠MxNRAMP1定位于质膜,MxNRAMP3定位于液泡膜[34],表达在酵母中可以转运镉,而不能转运其他金属。

图8 黄连NRAMP3在镉胁迫下的表达情况
本研究采用TA 克隆从黄连中克隆出了CcNRAMP3的编码基因,对CcNRAMP3跨膜结构域的预测结果表明,CcNRAMP3蛋白含有12 个可能的跨膜区域。通过对黄连CcNRAMP3基因编码的蛋白序列进行分析发现,在第98-461 氨基酸残基处含有一个NRAMP蛋白结构域,并在第209-212氨基酸残基处存在一个N-糖基化位点。亚细胞定位预测结果表明,CcNRAMP3蛋白定位于液泡膜上。以上结果说明,黄连CcNRAMP3具有NRAMP蛋白家族的重要特征,是一个典型的NRAMP蛋白家族成员。序列相似性比对和系统进化树分析表明,黄连CcNRAMP3蛋白与博落回、莲、烟草等植物的NRAMP3蛋白亲缘关系最近,序列相似性高,表明CcNRAMP3可能与这些蛋白具有相似功能,可以转运诸如镉等常见金属离子。与拟南芥NRAMP家族基因的AtNRAMP3和AtNRAMP4在蛋白水平上具有较高的相似性。AtNRAMP3与AtNRAMP4蛋白均定位在液泡膜上[14],这与CcNRAMP3一致。AtNRAMP3主 要 转 运 Fe、Mn 和 Cd 等 元 素 ,而AtNRAMP4除了转运Fe、Mn和Cd以外还能转运Zn,这表明黄连CcNRAMP3基因可能也具有Cd2+的吸收、转运功能。
为了研究CcNRAMP3基因在黄连内的分布及受Cd 胁迫后的表达情况,本实验利用qPCR 技术分析CcNRAMP3基因的时空表达模式。结果表明,CcNRAMP3基因在黄连各组织均有表达,且在叶片表达量最高。在受到Cd胁迫时,CcNRAMP3基因在须根中首先大量表达而后逐渐下降,这与植物吸收金属的吸收转运机制相似。整体上讲在须根中的表达情况是急剧升高而后逐渐降低的过程,这可能是黄连应对Cd 胁迫的一种调控方式,降低CcNRAMP3表达量,从而降低对Cd的吸收,抑或黄连吸收镉的量超过了自身耐受阈值,其自身防御系统被激发(植株死亡)而导致NRAMP基因的表达量下降。结合CcNRAMP3定位在液泡上,以上推测其与重金属Cd2+转运相关。
随着工业的发展,重金属污染对生物造成了严重威胁,根据其生物有效性浓度和受体暴露敏感性,重金属具有潜在的毒性。大量研究表明植物必需金属与非必需金属存在竞争吸收,如Zn2+/Cd2+。在这些金属中,那些类似于离子价态营养素的元素(如Hg、Pb和Cd)被认为构成了更大的威胁,因为它们可以通过现有的金属吸收通道被植物吸收和利用,如何有效地减轻重金属污染的问题已经引起了全世界的关注。本研究首次克隆出了CcNRAMP3基因,通过生物信息学分析及序列比对分析,确定该基因的理化性质及结构,结合其在黄连植物体内表达情况及在镉胁迫下的表达情况,认为该基因可能与黄连Cd 转运吸收有关,同时为后续抗性植物的筛选奠定了基础。未来可考虑应用基因工程技术将抗性基因或决定镉超富集的基因导入理想植物体内,进行基因重组和表达,或者通过抑制转运Cd的基因,从而抑制黄连吸附重金属的效率,从而降低黄连植株中的镉含量,真正意义上促进中药材的无公害种植。
