基于树冠形态特征因子的云南红豆杉单木生物量模型拟合
2022-08-26欧建德欧家琳康永武
欧建德 欧家琳 康永武
(1. 明溪县林业局,福建 三明 365200;2. 福建理工学校,福建 福州 350000;3. 沙县林业局,福建 三明 365500)
森林生物量是森林生态系统中最基础的数量特征[1],是影响并决定着森林经营决策的基础。单木生物量模型是估算森林生物量的基础[2],提高单木生物量模型的预测精度一直是林业科技的重点与难点。模型选择与优化[1−2]、自变量的选取[3]是提高生物量模型预测精度的主要途径。鉴于当前植物生物量模型的建立大多基于生长性状[4−6]变量进行,未见导入总体树冠结构特征自变量。考虑到树冠结构改变植物光合作用、同化能力[7−8],影响生物量积累,选取导入树冠特征作为自变量,以提高生物量模型的预测精度十分重要。
云南红豆杉(Taxus yunnanensis)(以下简称红豆杉)的紫杉醇半合成前体10−脱乙酰巴卡亭Ⅲ(10−DAB)累积含量高[9],是紫杉烷原料林的主栽树种和珍贵用材树种,建立并优化单木生物量模型,精准预测原料林生物量(产量)十分必要。当前原料林研究多关注遗传规律[10−11]、培育[12−13]、采收季节[14−15]等,尚未见单木生物量模型报道。冠幅、冠形率和树冠率是树冠形态的二维指标体系[7,16−18],亦是调控树冠形态结构的指标性状[7,16−18]。因此,以福建省明溪县3年生云南红豆杉幼林为研究对象,在生长性状(地径、树高)自变量基础上,全面导入冠幅、冠形率、树冠率(树冠形态结构)等自变量,运用逐步回归和独立样本检验的方法,旨在建立、评价并优化红豆杉系列单木生物量模型,明确理想的树冠形态结构与调控措施,以期为紫杉烷原料林经营提供技术与理论支持。
1 研究区概况
研究地在福建省西北部的明溪县(26 °8′~26 °39 ′ N、117 °4 ′~118 °47 ′ E),主要以低山丘陵地貌为主,地处武夷山脉南面,气候属中亚热带海洋性季风气候,年均温18.1 ℃,年均降水量1786 mm,5—6月最多,全年平均蒸发量1364 mm,年均日照时数1750.7 h,无霜期283 d,年均相对湿度81%[19]。供试材料取自明溪县王桥的3年生云南红豆杉原料林林分,株行距25 cm×25 cm,林分郁闭度0.7,平均地径1.2 cm,平均树高97 cm。海拔300 m,平均坡度5°左右,山地红壤,立地肥沃。
2 研究方法
2.1 性状测定
2019年12 月在原料林基地内,随机布设1 m×1 m的样方8个,逐株标志并测量红豆杉地径、树高、枝下高、东西和南北冠幅等,分别精确至0.1、1、1、1 cm,共计128株植株。采用全挖法,挖取样方内全部植株,带回实验室,测定枝叶生物量、茎干生物量、根系生物量。生物量测定参考文献[20]的方法执行。样木基本情况见表1。

表1 样本基本情况Table 1 Basic information of samples
树冠形态特征及生物量按以下公式计算 :



2.2 生物量数学模型的拟合与优选
地径、树高、冠幅均匀分布,按照典型抽样的方法抽取,选择80%(103棵)红豆杉样本数据作为建模数据。生物量模型表达式如下:
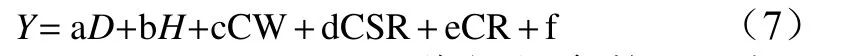
式中:a、b、c、d、e、f是待解的参数;Y表示枝叶生物量(Ly)、茎干生物量(Sy),地上生物量(Ay),总生物量(Ty);D表示地径,H表示苗高,CW表示冠幅,CSR表示冠形率,CR表示树冠率。
采用逐步回归方法拟合数学模型,根据拟合模型判定系数(R2)的大小,筛选出最优数学模型[1,23]。
2.3 优选模型检验
采用独立样本检验评价模型预测水平和精度,即以剩余的20%(25棵)红豆杉样本数据作为检验数据,利用系列优选生物量模型,分别计算出检验数据的生物量预测值后[1,23],对预测值与实测值进行配对样本t检验[1,23]、计算平均偏差(ME)、平均绝对偏差(MAE)、平均相对偏差(MPE)、平均相对偏差绝对值(MAPE)等系列偏差指标,估算预测模型精度(P)评价模型预测能力[1,24−26];偏差检验与模型预测精度估算按参考文献[24−26]的方法执行。
2.4 数据处理
采用Excel 2003软件进行数据预处理与统计;采用SPSS 21.0软件中逐步回归方法拟合数学模型,配对样本t检验方法检验预测值、实测值间差异。
3 结果与分析
3.1 单木枝叶生物量模型拟合与比较
红豆杉单木枝叶生物量模型的逐步回归拟合结果见表2。在模型的拟合过程中,变量D、H、CW、CSR依次进入,始终未出现CR变量,说明CR单独作用不显著。在拟合模型1~3过程,模型参数均通过参数t检验(P<0.01),说明D、H、CW对枝叶生物量均有显著直接作用。模型调整判定系数(R2)随变量D、H、CW的依次进入而持续增大,导入CW变量可提高拟合效果。在模型3基础上导入CSR变量,导致模型4中H变量未通过参数t检验(P=0.158>0.05),为此继续逐步回归拟合得模型5,模型5的系列参数(常量、D、CW、CSR)均通过参数t检验(P<0.01)。由表2可知,模型5中所有参数均通过t检验且R2最大(0.812),为最优模型;模型中CW、CSR变量系数分别为1.181、18.372,均大于0,表明冠幅和冠形率有显著促进枝叶生物量效应。结果(表2)还显示,模型2为导入树冠形态特征因子前(以下简称导入前)最优模型(R2最大为0.762)。
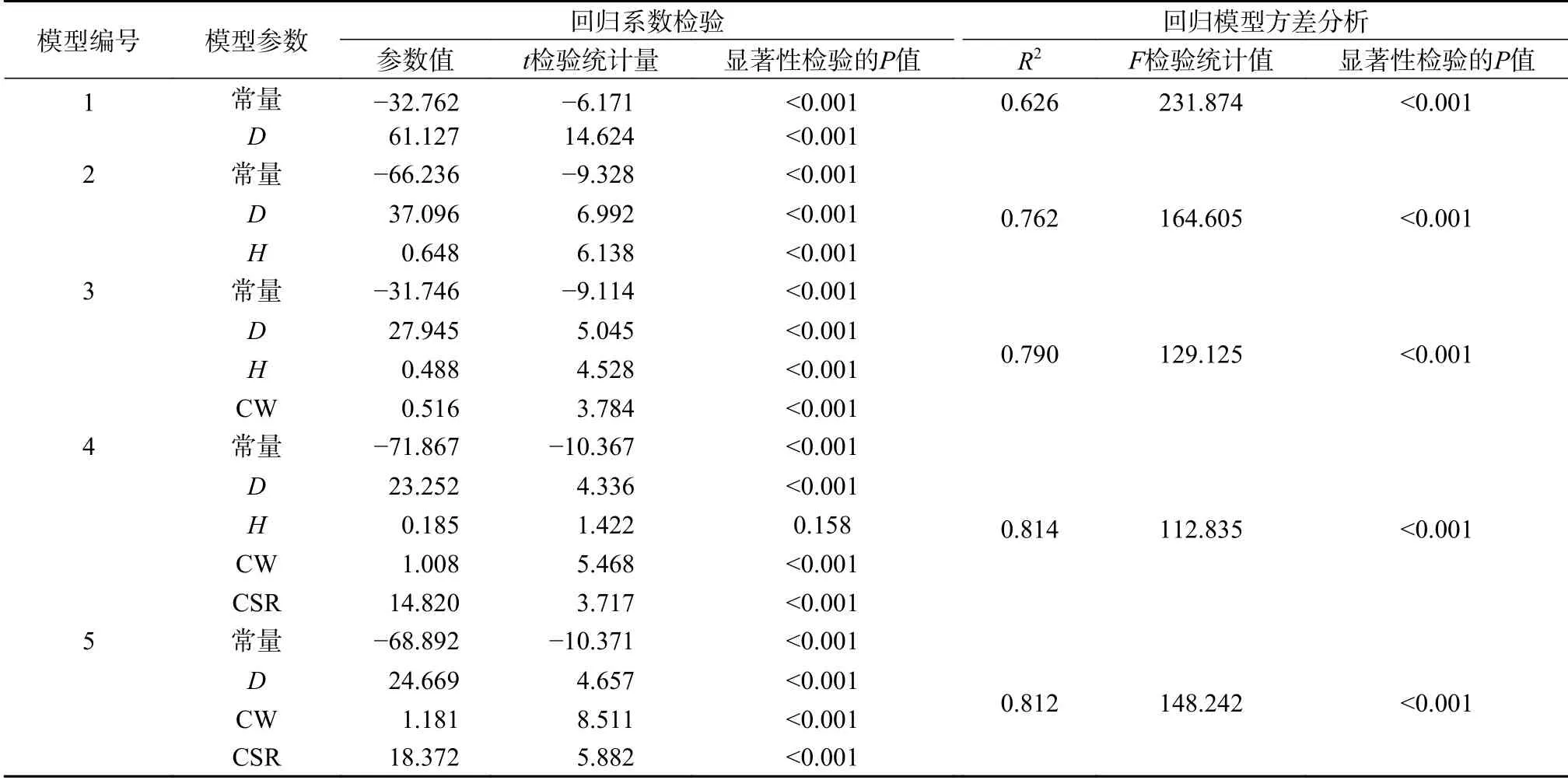
表2 单木枝叶生物量数学模型拟合结果Table 2 Fitting results of the single tree branches and leaves biomass mathematical model of T. yunnanensis
为分析导入树冠形态特征因子前后(以下简称导入前后)系列生物量模型的预测效果,采取独立样本检验方法,对系列导入树冠形态特征前后的2种最优模型(R2最大且模型参数均通过t检验)进行配对t检验、偏差检验和预测精度估算比较,结果见表3。

表3 优化的单木红豆杉生物量模型检验结果Table 3 Test results of optimized single tree biomass model of T. yunnanensis
由表2可知,导入前后最优模型(模型2、5)的R2分别0.762、0.812,说明导入后提高了模型的拟合效果。由表3可知,导入前后模型配对t检验的P值分别为0.040、0.620,意味着导入前模型的实测值、拟合值间差异显著,而导入后模型实测值、拟合值间差异不显著,表明导入后提高了模型的预测效果。导入前后模型的预测精度分别达89.67%、92.50%,导入后提高了模型预测精度。导入后模型的ME、MAE、MPE、MAPE分别为0.81 g、5.94 g、−5.22%、18.64%,总体较导入前模型理想。
综上,导入树冠形态特征因子后明显提高模型的拟合与预测效果,与前文中冠幅和冠形率显著促进枝叶生物量的结论相吻合。
3.2 单木茎干生物量模型拟合与比较
红豆杉单木茎干生物量模型的逐步回归拟合结果见表4。在模型的拟合过程中,变量D、H、CW、CSR依次进入,始终未出现CR变量,说明CR的单独作用效应不显著;在模型6~8的拟合过程,模型的参数均通过参数t检验(P<0.01),说明D、H、CW对茎干生物量均有显著直接作用;系列模型的R2随变量D、H、CW的依次进入而持续增大,说明导入CW变量提高了模型拟合效果。在模型8基础上导入CSR得模型9,但模型9中H变量未通过参数t检验(P=0.198>0.05),为此继续逐步回归拟合得模型10。模型10参数(常量、D、CW、CSR)均通过参数t检验(P<0.01)。综合6~10系列模型的参数t检验和R2值表现(表4),模型10所有参数均通过参数t检验(P<0.01)且R2值最大(0.818),为最优模型。模型中CW、CSR系数分别为0.540、8.105,均大于0,表明冠幅和冠形率显著促进茎干生物量效应。结果(表4)还显示,模型7为导入树冠形态特征前最优模型(R2最大为0.773)。
结果(表4)显示,导入树冠形态特征前后最优模型(模型7、10)的R2分别为0.773、0.818,说明导入后提高了模型的R2、拟合效果。结果(表3)显示,导入前后最优模型配对t检验的P值分别为0.033、0.527,意味着导入前的拟合值、实测值间差异显著,而导入后的实测值、拟合值差异不显著、预测效果好,表明导入后提高模型的预测效果。导入前后最优模型的预测精度分别为88.61%、92.36%,即导入后提高了模型的预测精度。导入后最优模型(模型10)的ME、MAE、MPE、MAPE分 别 为0.46 g、2.29 g、−2.73%、18.95%,较导入前(模型7)的相应偏差性状均有更好表现。

表4 单木茎干生物量数学模型拟合结果Table 4 Fitting results of the single tree stem biomass mathematical model of T. yunnanensis
综上,导入树冠形态特征因子明显提高模型的拟合与预测效果,与前文中冠幅和冠形率显著促进茎干生物量的结论相吻合。
3.3 单木地上生物量模型拟合与比较
红豆杉单木地上生物量模型的逐步回归拟合结果见表5。在模型的拟合过程中,变量D、H、CW、CSR依次进入,始终未出现CR变量,说明CR单独作用不显著;在模型11~13的拟合过程,模型参数均通过参数t检验(P<0.01),说明D、H、CW变量对地上生物量均有着显著的直接作用;模型R2随着变量D、H、CW的依次进入而持续增大、拟合效果持续变好,说明导入CW变量可提高模型拟合效果。在模型13基础上导入CSR变量得模型14,但模型14中H变量未通过参数t检验(P=0.170>0.05),继续回归拟合得模型15。综合系列模型11~15的参数t检验和R2值表现(表5),模型15的参数均通过的参数t检验(P<0.01)、且R2值最大(0.815),为最优模型。模型中CW、CSR的系数分别为1.721、26.427,均大于0,表明冠幅和冠形率对地上生物量有显著促进效应。结果(表5)还显示,模型12为导入树冠形态特征前最优模型(R2最大、0.766)。
由表5可知,导入前后最优模型(模型12、15)的R2分别为0.766、0.815,说明导入后提高了模型的R2、拟合效果。由表3可知,导入前后配对t检验的P值分别为0.041、0.587,意味着导入前拟合值、实测值间差异显著,预测效果差,而导入后拟合值、实测值间差异不显著,预测效果好,表明导入树冠形态特征可提高模型的预测效果。导入前后最优模型的预测精度分别89.39%、92.51%,即导入后提高了模型的预测精度。导入后最优模型(模型15)的ME、MAE、MPE、MAPE分别为1.28 g、8.22 g、−4.47%、19.82%,总体较导入前(模型12)的表现理想。

表5 单木地上生物量数学模型拟合结果Table 5 Fitting results of the single tree aboveground biomass mathematical model of T. yunnanensis
综上,导入树冠形态特征因子明显提高模型的拟合与预测效果,与前文中冠幅和冠形率显著促进地上生物量的结论相吻合。
3.4 单木红豆杉总生物量模型拟合与比较
红豆杉单木总生物量模型的逐步回归拟合结果见表6。在模型的拟合过程中,变量D、H、CR、CW依次进入,始终未出现CSR变量,说明冠形率CSR单独作用不显著。模型R2随变量D、H、CR、CW的依次进入而持续增大、拟合效果持续变好,说明导入树冠形态特征因子变量提高了模型拟合效果。综合系列模型16~19的参数t检验、R2值表现(表6),模型19中参数均通过参数t检验(P<0.01)、且R2值最大(0.831),为最优模型。模型中CR、CW的系数分别为65.347、0.808,均大于0,表明树冠率和冠幅对红豆杉总生物量有着显著性促进效应。结果(表6)还显示,模型17为导入前最优模型(R2最大为0.780)。
由表6可知,导入树冠形态特征前后最优模型(模型17、19)的R2分别为0.780、0.831,说明导入后提高了模型的R2、拟合效果。由表3可知,导入前后配对t检验的P值分别为0.060、0.587,虽然导入前后拟合值、实测值均无显著性差异,但导入后的配对t检验的P=0.587大于导入前的P=0.060,表明导入树冠形态特征可提高模型的预测效果。导入前后最优模型的预测精度分别90.57%、92.58%,即导入后提高了模型的预测精度。导入后最优模型(模型19)的ME、MAE、MPE、MAPE分别为0.31 g、9.64 g、−4.26%、13.96%,总体较导入前(模型17)的表现理想。

表6 单木红豆杉总生物量数学模型拟合结果Table 6 Fitting results of the single tree total biomass mathematical model of T. yunnanensis
综上,导入树冠形态特征因子后明显提高模型的拟合与预测效果,与前文中树冠率和冠幅显著促进红豆杉总生物量的结论相吻合。
4 结论与讨论
本研究全面导入系列树冠形态特征因子(冠幅、冠形率、树冠率)自变量后,建立了红豆杉系列单木生物量模型,提高了模型的预测精度与水平,可为同类研究提供参考与借鉴;系列单木生物量优化模型预测精度均在92.36%以上,且偏差表现良好,预测精度高,可用于红豆杉原料林生物量与产量预测预估。建议在云南红豆杉原料林培育过程中,首先采用良种壮苗、优化密度和幼林抚育以促进地径生长,其次宜结合培育模式(收获物的差异)调适树冠结构调控措施与策略。
研究结果显示,单木枝叶、茎干和地上生物量模型的实测值与拟合值差异显著(P<0.05),且预测精度均小于90%,表明拟合红豆杉单木枝叶、茎干和地上生物量模型仅导入地径、苗高等生长自变量是不够的,该系列单木生物量模型中导入树冠形态特征变量是必要的,验证了生物量模型主要以与冠幅相关的因子为自变量[27]以及含有树冠形态特征变量的单木生物量预测精度表现好[28]的研究结论一致。
系列单木生物量模型的逐步回归拟合过程显示,率先进入的变量均为地径,其次是树高,且均表现出显著促进作用,表明地径在直接促进系列生物量的作用超过树高,是主导影响因子。提高原料林生物量(产量),首先是促进地径生长。进入系列最优生物量模型的树冠形态特征与先后顺序明显变化,表明树冠形态特征作用系列生物量规律,以及利于系列单木生物量积累的理想树冠形态结构与调控措施明显不同,建议生产中宜根据目标收获物的差异调适树冠结构调控措施与策略。这与因调控目标改变调整树冠调控措施[7,17−18]的研究结论相互验证。先后依次进入单木枝叶、茎干和地上生物量最优模型的树冠形态特征是冠幅、冠形率,且均有显著促进生物量积累作用。这意味着促进该系列生物量积累的理想树冠形态结构首先是冠幅宽大、其次为狭长的冠形。促进生物量积累的树冠形态结构调控重点与方向是促进冠幅宽大,建议在以采收枝叶、地上生物量为目标的原料林矮林作业模式中,宜选择保留或采用冠幅宽大的个体、品种为主,或减少密度以促进冠幅生长。先后依次进入单木总生物量最优模型的树冠形态特征为树冠率、冠形率,且均有显著促进总生物量积累作用,意味着促进总生物量理想树冠形态结构首先是大的树冠率,其次是冠幅宽大。促进总生物量积累的树冠形态调控重点与方向是提高树冠率,建议在全挖式采收的短周期原料林模式中,宜保留或采用树冠率大的个体、品种为主,以及经营过程减少降低树冠率的行为措施。
导入树冠形态结构特征,采用逐步回归的方法,明显提升了云南红豆杉单木生物量模型的预测精度与水平,与前人采用逐步回归的方法优化预测模型[4−6,20,29]以及在自变量不断增加情况,模型的拟合精度有不断提升的趋势[28]结论相互验证。结果表明树冠结构显著影响原料林单木生物量积累,这是原料林树冠既是枝叶生产器官,又是光合作用的同化器官,其树冠结构改变了利用地上空间能力、影响同化能力与生产力所致。这与前人关于树冠结构影响同化能力、地上空间利用能力[7−8]和林木生产力[7−8,30]的研究结论相互验证。王轶夫等[31]认为冠长、冠幅等树冠自变量与马尾松(Pinus massoniana)树冠、树枝、叶花果等生物量相关关系不明显;造成结论差异的原因,与树种、培育模式以及年龄不同有关。同时亦验证了生物量模型的自变量选取因树种明显不同[27]的结论。
本研究仅从导入树冠形态特征自变量方面,拟合并优化单木生物量模型,有关模型的自变量类型、模型选择等优化研究尚待后续。
